1. Introduction
In 1999, Mitsui [
1] criticized the power-stroke model on the muscle contraction mechanism and proposed a new model. In 2008, Mitsui and Ohshima [
2] refined the new model and discussed the steady filament sliding in detail demonstrating that the calculation results were in good agreement with experimental observations. They also outlined the discussion on the isometric tension transient and the isotonic velocity transient given in [
1] by citing some calculation results. Thereafter, however, a few readers of [
1] commented that it was very difficult to understand the discussion on the transient phenomena in [
1], since there was no detailed explanation on the molecular processes upon which the theoretical treatment was based. In the present paper, we have largely revised that part of [
1], trying to make the discussion more readable.
Now the article [
2] is regarded as Part I of Remarks series, in which the basic ideas of our model are introduced and steady muscle behaviors are discussed. In the present paper (Remarks II), non-steady muscle behaviors are discussed. We are preparing an article as Remarks III, in which discussion will be done on more recent experimental studies as cited in the articles [
3,
4].
The basic ideas of our model introduced in [
2] are summarized as follows. A simple thermodynamic relation is derived, which indicates that there is an inconsistency in the power stroke model or swinging lever model. Our model is proposed to avoid this difficulty. It is assumed that a myosin head forms a complex with three actin molecules when it attaches to an actin filament. Here it should be noted that Andreeva
et al. [
5] found the evidence that the crossbridge can interact with more than 1 actin monomer. The complex corresponds to the crossbridge. According to the X-ray diffraction studies [
6–
9], the intensity ratio of the [1, 0] and [
1,
1] equatorial reflections increases only minimally as the shortening velocity increases, indicating that the total number of myosin heads in the vicinity of the actin filament decreases only slightly. Taking this fact into account, it is assumed that about 41% of the myosin heads forms the crossbridges at any sliding velocity. Then mutual cooperativity takes place among the crossbridges in filament sliding, so that energy dissipation becomes reasonable magnitude (the order of
kT) for one step of the crossbridge movement. Calculation based upon the model well reproduce the force-velocity relation given by Hill [
10] and the energy liberation rate
vs. force relation given by Hill [
11].
Since the present study is based upon ideas that are quite different from the power stroke model and others, the basic ideas of our study are explained in some detail in Section 2. In Section 3, discussion is done of how variation in the crossbridge binding affects the muscle tension. In Section 4, discussion is done on the difference between the molecular processes in Phases 1 of the isometric tension transient and of the isotonic velocity transient, and extensibility ratios for the crossbridge, the myosin filament and the actin filament are estimated. Time course of the isometric tension transient is studied in Section 5. Time course of the isotonic velocity transient is studied in Section 6. The deviation from the Hill-type force-velocity relation is derived in Section 7. Obtained results are summarized and discussed in Section 8.
3. Effect of U*12 Transition on the Tension
Equation 8 leads us to the Hill-type force-velocity relation. The stress
T and
yc related by
Equation 8 is denoted as
TH and
ycH, which are in agreement with the observation when
ycH >
ycdev. The observed
T and
yc are denoted as
T* and
yc* when
ycH <
ycdev.
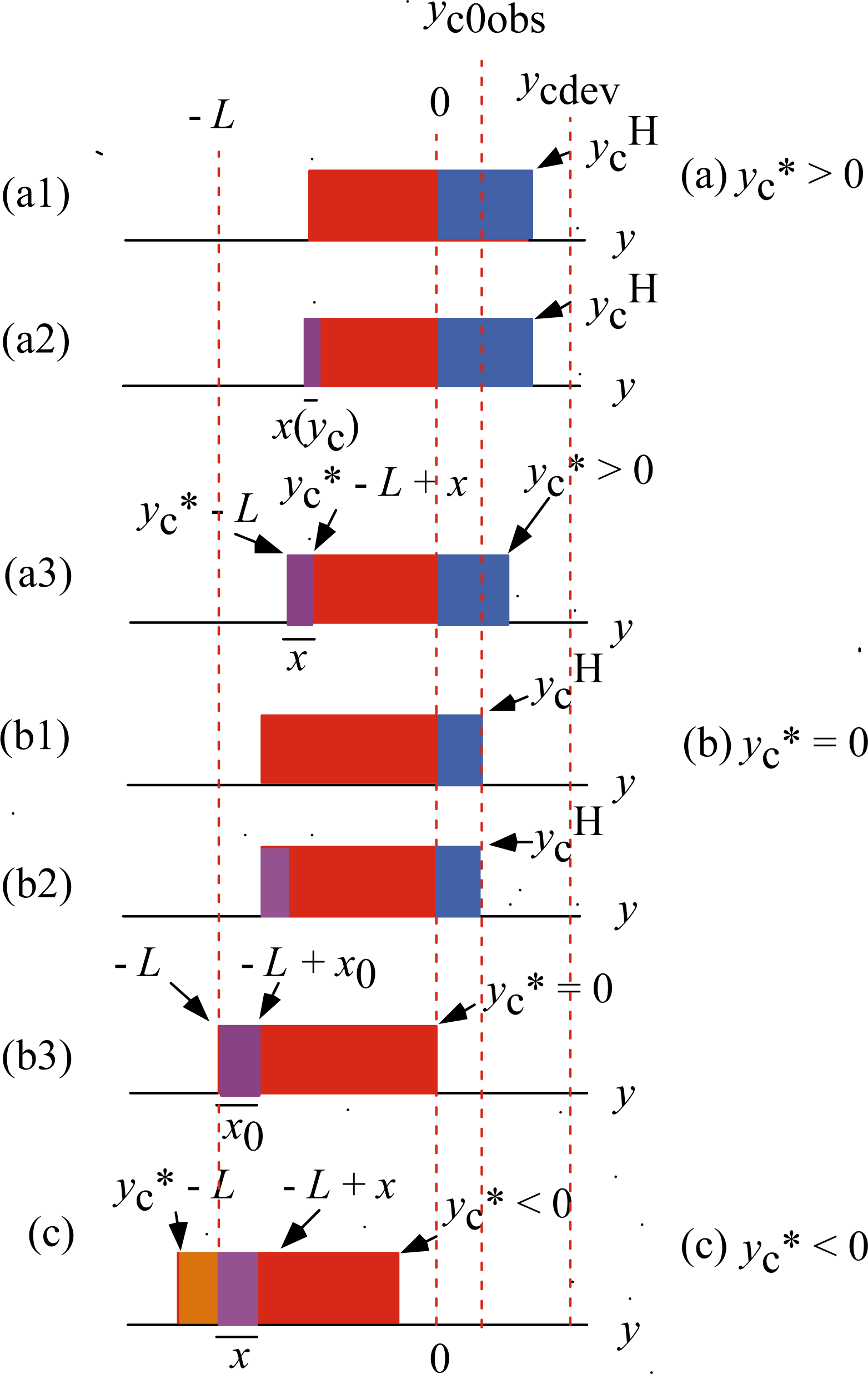
Figure 7 illustrates the
ρ distributions in various cases of
ycH <
ycdev.
Figure 7(a1) shows the
ρ distribution by
Equation 8 having the edge
ycH. This
ρ distribution gives
TH even though
yc <
ycdev. It is an imaginary state in which the
U*
12 transition is absent. Actually, however, the
U*
12 transition takes place and the violet area appears causing decrease of the red area as shown in (a2), where the width of the violet area is denoted as
x(
yc). Naturally
T in (a2) is smaller than
T in (a1). Then
yc in (a2) is changed into
yc* in (a3) to make
T the same as
T in (a1), where the width of the violet region is denoted as
x. The change of the blue area from
yc to
yc* causes an increase of the tension while the change of the violet area from
x(
yc) to
x causes an decrease of the tension. If the effect of the former change is larger than that of the latter change, the tension in (a2) will increase and can be the same as in (a1). Below we shall assume that such tension adjustment actually takes place and discuss the relation between
yc* and
T*.
Since the tension is the same in
Figures 7(a1) and (a3), the blue area in (a1) is the same as the sum of the blue area and violet area in (a3).
Figures 7(b1)–(b3) show the case of
y* = 0. Similarly to (a1) and (a3), the blue area in (b1) is the same as the violet area in (b3). The width
x of the violet area in (b3) is denoted as
x0.
Figure 7 (c) shows the case that
yc* becomes negative. The brown area means that crossbridges in this area bind to A1 and produce positive stress. The width of the violet area is indicated as
x. Extrapolating the change from
x in (a3) to
x0 in (b3),
x in (c) is assumed to be larger than
x0. Then sum of the red area and brown area in (c) is smaller than red area in (b3) and thus the tension produced in (c) is smaller than the tension in (b3). Then the stress produced in (b3) is the maximum that the muscle machine can produce, and should be equal to
T0obs. Accordingly,
ycH in (b1) should be equal to
yc0obs. Since the red area in (b1) is the same as the red area in (b3),
ycH in (b1) is equal to
x0 in (b3). Thus we have
The tension
T* deviates from
TH at
yc =
yc* =
ycdev, and reaches its maximum at
yc* = 0. To express such characteristics of
T*
vs. y* relation, the following set of equations are used:
In
Figure 8, the blue curve illustrates the
T*/
T0 vs. yc* relation given by this set of equations, and the red curve shows the Hill-type
TH/
T0 vs. ycH relation. The blue curve deviates from the red curve at
yc =
yc* =
ycdev and exhibits its maximum at
yc* = 0 where
T*/
T0 =
T0obs/
T0.
Up to here,
T/
T0 is used for the relative tension, where
T0 is the maximum tension in the extended Hill-type force-velocity relation. The experimentally observed maximum tension, however, is
T0obs where
T0obs = 0.88
T0 (
Equation 2). Hereafter discussion is concerned with experimental data and relative tension is defined as
T/
T0obs. In our model
T/
T0 =
p/
p0 (Equation 3–1–2 in [
2]), and the force
p corresponding to
T0obs is denoted as
p0obs. Then
and
If we put
yc =
yc0obs in
Equation 7 and use
x0 =
yc0obs (
Equation 11), we get theoretical expression of
p0obs as
4. Phases 1 in the Isometric Tension Transient and Isotonic Velocity Transient, and Extensibility Ratios for the Crossbridge, Myosin Filament and Actin Filament
Huxley [
15] divided the transient responses to the sudden reduction of length or of load into four Phases. In our model, however, there is no exact correspondence between molecular processes of the four Phases in the tension transient and those in the velocity transient. To avoid confusion, we use the terms, Phase Tn in the tension transient and Phase Vn in the velocity transient, where n = 1, 2, 3, 4.
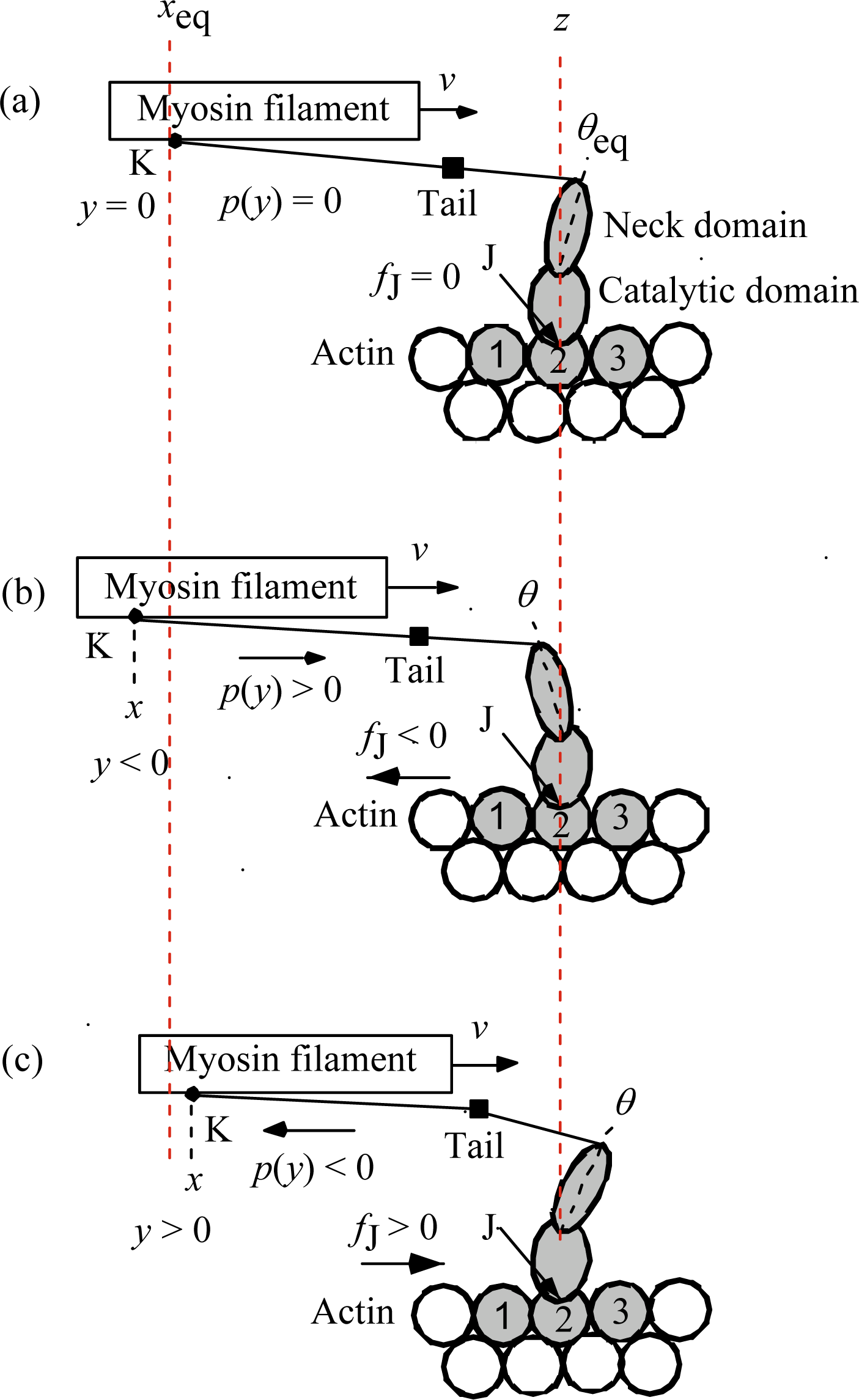
Phase T1 is simultaneous decrease of tension in the isometric tension transient, while Phase V1 is simultaneous shortening of muscle in the velocity transient. The length change per half sarcomere is denoted as Δ
Lhs in both transients. The experimentally measured isometric tension is
T0obs and the relative tension is defined by
T/
T0obs. The relative load used in the paper by Civan and Podolsky [
8] is expressed as the relative tension
T/
T0obs below.
Figure 9 shows the experimental results on Phases T1 and V1, by circles for the isometric tension transient cited from the paper by Ford
et al. [
16] and by squares for the isotonic velocity transient cited from the paper by Civan and Podolsky [
17].
At first sight, it was puzzling to see that distributions of the experimental data are quite different for the two cases, since the structural changes seem to be purely elastic both in Phases T1 and V1. Then it was reminded that the length change of sarcomere is a sum of those of the crossbridge, myosin filament and actin filament, which are proportional to each other (cf. the review by Irving [
18]). This means that there are three elastic components, and it is a possibility that they have different response times in elastic changes from each other.
In this connection, the experimental data reported by Julian and Sollins [
19] seem important. They measured tension changes of single frog skeletal muscle fiber at increasing speed of step shortening. Their
Figures 2 and
4 show experimental data on relative force
vs. Δ
Lhs relation at different speeds of shortening. The distribution of open circles in their
Figure 2 (the length change period of about 1 ms) is similar to that of the data by Civan and Podolsky (1∼2 ms) cited in our
Figure 9. The distribution of filled triangles in
Figure 4 of [
9] (length change period of 0.4 ms) is close to the data by Ford
et al. (the length change period of 0.2 ms) cited in our
Figure 9. These facts seem to indicate that there are two kinds of elastic process in the tension response: The fast one occurs within about 0.4 ms and the slow one occurs between about 0.4 and 1 ms after the length change. The fast process seems responsible to the change in Phase T1 and combination of the fast and slow processes seems responsible to Phase V1.
It seems plausible that elastic response of crossbridge and myosin filament almost simultaneously occurs since they belong to the same molecule and elastic response of actin filament occurs with some delay. Thus it is assumed that crossbridge and myosin filament are responsible to the change in Phase T1 and that all three components are responsible to the change in Phase V1. Now changes of
yc in Phases T1 and V1 should be different from each other even when the length change Δ
Lhs is the same. Calculation is done on this assumption by using the rectangular
ρ distribution shown in
Figure 10.
Figure 10(a) shows the
ρ distribution at the isometric tension (the same as
Figure 7 (b3)).
Figure 10(b) shows the
ρ distribution just after Phase T1 and (c) the one just after Phase V1 for the same length change Δ
Lhs as (b). The violet areas in
Figure 10 (b) and (c) are the same as (a) since
U*
12 transition does not occur yet. Changes of the edge
yc in Phases T1 and V1 are denoted as Δ
ycT1 and Δ
ycV1. For the same Δ
Lhs, the tension
T in Phase T1 is smaller than
T in Phase V1 in
Figure 9. The edge Δ
ycT1 in
Figure 10 (b) is set larger than Δ
ycV1 in (c) so as to make the red area in (b) smaller than that in (c), in accordance with the fact that the tension
T in Phase T1 is smaller than
T in Phase V1.
Since elastic changes of the crossbridge, myosin filament and actin filament are proportional to each other [
10], the Δ
ycT1 in (b) and Δ
ycV1 in (c) are proportional to Δ
Lhs, and expressed by
and
where
CCBT and
CCBV are constants.
Let the tensions in
Figure 10(b) and (c) be denoted as
TT1 and
TV1, respectively. They are given by integration of −
κb{
y − (−
L)} in the violet area, −
κfy in the red area and −
κby in the blue area. Thus we have
and
Calculations were done for various trial values of
CCBT (
Equation 18) and
CCBV (
Equation 19). The best fit for the experimental data are obtained for the values
Figure 9 shows the calculation results for
TT1/
T0obs by the green curve and for
TV1/
T0obs by the brown curve. They are in good agreement with the experimental data.
Let us denote the ratios of extensibilities of the crossbridge, the myosin filament and the actin filament as
rCB,
rM, and
rA at the elastic equilibrium respectively. Shortenings of the three per half sarcomere are denoted, respectively, as Δ
yCB, Δ
yM and Δ
yA, and the length change of half sarcomere as Δ
Lhs. Then, at the elastic equilibrium, they are given by
Naturally,
On the above assumption, only the length changes of the crossbridge and the myosin filament contribute to Phase T1. Then we have
Thus,
As assumed above, the load change period is long enough and elastic changes of the crossbridge, myosin filament and actin filament contribute to Phase V1. Thus, Δ
ycV1 is given by
Hence,
By using the values of
CCBT (
Equation 22) and
CCBV (
Equation 23),
Equations 25,
27 and
29 give
The extensibility ratios were investigated by X-ray diffraction by Huxley
et al. [
20] and Wakabayashi
et al. [
21]. The values reported by Wakabayashi
et al. [
21] are
rCB = 0.31,
rM = 0.27 and
rA = 0.42. Considering approximate nature of the theory and experimental errors, our values in
Equation 30 are in reasonable agreement with the X-ray values.
In this section, it is assumed that elastic change of the actin filament does not take place during the fast length change in Phase T1. Then the elastic change of the actin filament should contribute to the next step, the tension recovery in Phase T2. This problem is discussed in next section.
5. Isometric Tension Transient
As noted in Section 4, Huxley [
15] divided the transient responses to the sudden reduction of length into four phases. Time courses of these Phases are as follows. Phase 1 is instantaneous drop of tension. Phase 2 is rapid early tension recovery in next 1∼2 ms. Phase 3 is extreme reduction or even reversal of rate of tension recovery during next 5∼20 ms. Phase 4 is the gradual recovery of tension, with asymptotic approach to the isometric tension.
As in Section 4, we use the terms Phase Tn (n = 1, 2, 3, 4) in the tension transient to avoid confusion. Phases Tn are more related with molecular process rather than time sequence. Phase T1 is simultaneous drop of tension caused by elastic shortening of the crossbridge and myosin filament. Phase T2 is due to elastic shortening of the actin filament. Phase T3 is related with the
U*
12 transition, and divided into T3a and T3b. Phase T3a is the first part of Phase T3. Both Phases T2 and T3a contribute to the rapid early tension recovery (Phase 2 of Huxley). Phase T3b is the second part of T3 where the extreme reduction or reversal of rate of tension recovery occurs (Phase 3 of Huxley). Phase T4 is the gradual recovery of tension, with asymptotic approach to isometric tension (Phase 4 of Huxley). (There were misprints in Section 4.4 of the article [
2], and the last three sentences of the section should be neglected.)
Figures 11 and
12 illustrate how the rectangular
ρ distribution changes during these Phases.
Figure 11 is for the case of Δ
ycT1 <
ycdev and
Figure 12 is for the case of Δ
ycT1 >
ycdev, where Δ
ycT1 is
yc just after Phase T1. (cf.
Figure 10(b)).
As mentioned above, it is assumed that the elastic changes of the crossbridge and myosin filament occur in Phase T1 and then the elastic change of actin filament occurs in Phase T2.
Figure 11(a) represents the states just after Phase T1.
Figure 11(b) shows the state just after Phase T2. There is a time lag for the
U*
12 transition to occur, and its effect is neglected in Phases T1 and T2, so that the width of the violet area is kept the same as the isometric value
x0.
As noted referring to
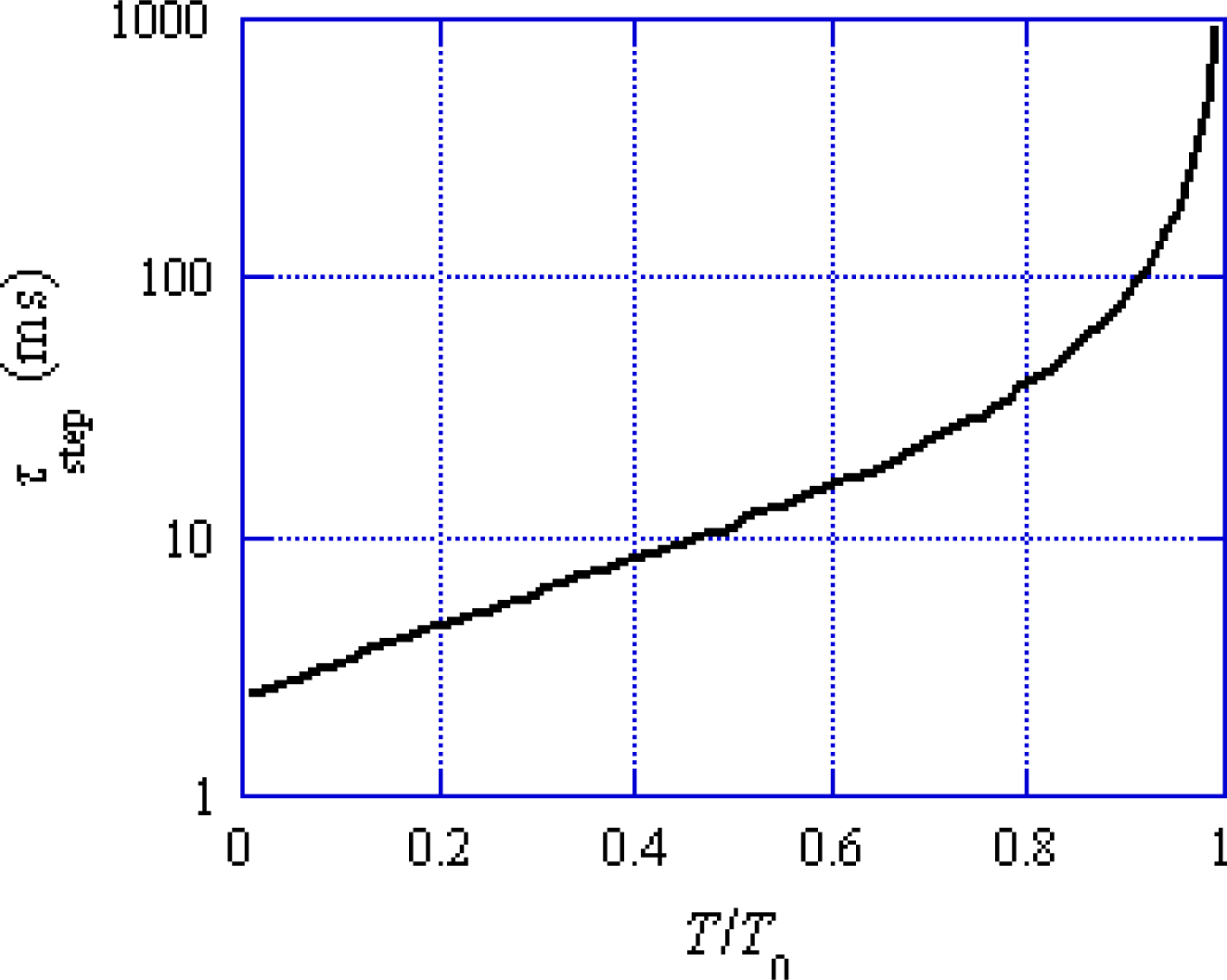
Figure 5, the mean time (
τstep) needed for the
U* transition is relatively large (e.g., larger than 10 ms for
T/
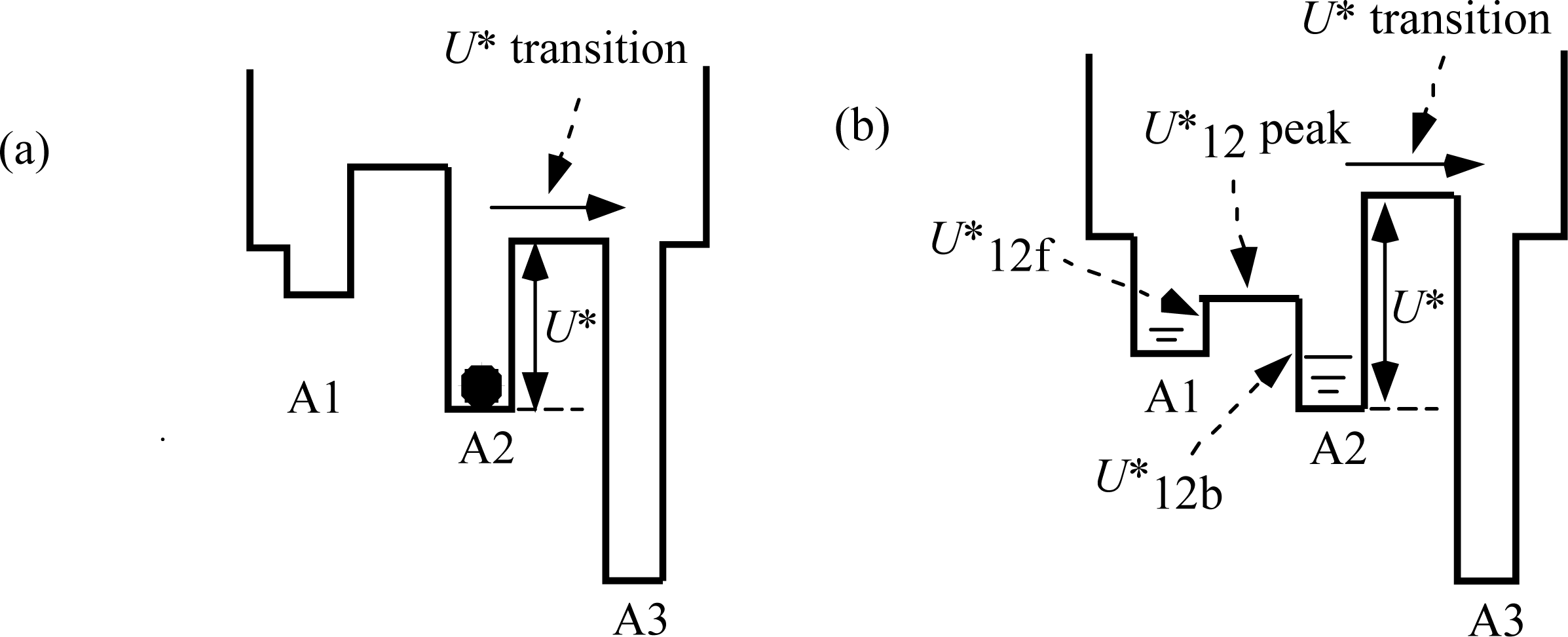
T0 > 0.5). It is assumed that the potential barrier
U*
12f (f: forward) is lower than
U* as shown in
Figure 3 (b) and thus the
U*
12f transition starts before the
U* transition. Phase T3a is regarded as the state that the
U*
12f transition is present but the
U* transition does not occur yet, while the
U*
12f and
U* transitions coexist in Phase T3b. In Phase 4, only the
U* transition exists.
Figure 11(c) and (d), respectively, show the states at the end of Phases T3a and T3b. The violet area in (c) is smaller than in (b) because a portion of the crossbridges in the violet area change their binding partners from A1 to A2 through
U*
12f transition. Thus the red area increases and the stress is stored and the tension increases in Phase T3a. The
U* transition starts at (c) and the stress stored during Phase T3a is released by the shift of the edge
yc from
ycT3a in (c) to
ycT3b in (d). The shift of the edge from
ycT3a (c) to
ycT3b (d) causes increase of the blue area and decrease of the violet area. These two effects tend to cancel each other and reduce change of the red area in the time course from (c) to (d),
i.e., in Phase T3b. The reduced change of the red area may cause the “extreme reduction of rate of tension recovery” mentioned at the beginning of this section. In Phase 4, the
U* transition and the filament sliding continue and the state approaches to the tetanus state (e).
Figure 12 shows changes of the
ρ distribution in the isometric tension transient for the case of Δ
ycT1 >
ycdev.
Figure 12(a) and (b) are similar to
Figure 11(a) and (b). Since Δ
ycT1 >
ycdev, the
U*
12f transition does not leave the violet area and the red area significantly increases in (c). Then the
U* transition starts and the
ρ distribution shifts to the right as shown in (d). The red area significantly decreases from (c) to (d), and tension decreases and thus the “reversal of rate of tension recovery” cited at the beginning of this section is expected. The filament sliding continues in Phase 4, and the
ρ distribution asymptotically approaches the isometric tetanus state (e).
Figure 13 illustrates changes in Phases T1∼T4 in another way by the
T/
T0obs vs. Δ
Lhs relation. The solid black arrows “Phase Tn” indicate an example of change of relative tension in Phase Tn for the case of Δ
y <
ycdev and the dashed black arrows for the case of Δ
y >
ycdev. The edge Δ
ycT1 in
Figures 11(a) or
12(a) is resulted from the elastic changes of the crossbridge and myosin filament, and is given as a function of Δ
Lhs by combining
Equations 18 and
22:
The tension variation
TT1/
T0obs in Phase T1 can be calculated as a function of Δ
Lhs by using
Equations 20 and
31. Calculation results are shown by the green curve in
Figure 13. The edge
ycT2 in
Figures 11(b) or
12(b) are the same quantity as Δ
ycV1 in
Figure 10(c), since they result from the elastic changes of the crossbridge, myosin filament and actin filament. Then, from
Equations 19 and
23, we have
Δ
ycT2 = Δ
ycV1 means
TT2/
T0obs =
TV1/
T0obs. By using
Equation 32 and replacing
TV1 by
TT2 and Δ
ycV1 by Δ
ycT2 in
Equation 21,
TT2/
T0obs can be calculated as a function of Δ
Lhs. The calculation result is given by the brown curve in
Figure 13. In the steady filament sliding, there is the definite relation between the tension
T and the parameter
yc as shown in
Figure 8, where the red curve shows the relation for the Hill-type filament sliding and the blue curve shows the relation when the filament sliding deviates from the Hill-type. The parameter
yc in
Figure 8 is related with Δ
yc in
Figures 11 and
12 by the relation
yc =
yc0obs + Δ
yc since Δ
yc is the change of
yc from the value at the isometric tension (
yc0obs). In the steady filament sliding, all elastic elongations of the crossbridge, the myosin filament and the actin filament contribute to the muscle elongation and thus Δ
yc (=
yc −
yc0obs) is equal to −Δ
Lhs/4.6 as in
Equation 32. The relative tension
T/
T0 in
Figure 8 can be converted to
T/
T0obs by using the ratio
T0obs/
T0 = 0.88 (
Equation 2). Based upon these considerations, the red and blue curves in
Figure 8 are reproduced with the same colors in
Figure 13.
In
Figure 13, the Phases are indicated by the solid black arrows “Phase Tn” in the case of Δ
y <
ycdev (the case of
Figure 11), and by the dashed black arrows in the case of Δ
y >
ycdev (the case of
Figure 12). The rapid tension change in Phase T1 occurs along the green curve as indicated by the solid or dashed arrows “Phase T1”. Phase T2 is a result of the elastic change of the actin filament and thus the arrows “Phase T2” start from the green curve and end on the brown curve. The main part of Phase T3a corresponds to the rapid tension increase from
Figure 11(b) to (c) or from
Figure 12(b) to (c). It is difficult to determine where Phase T3a turns into Phase T3b in
Figure 13. Trial calculations, however, show that fairly good agreement can be obtained by assuming that Phase T3a ends at the blue curve or the red curve. Thus, in the case of Δ
y <
ycdev, the solid arrow “Phase T3a” is depicted with its tip on the blue curve. As discussed above, it is plausible that the red area in
Figure 11(d) is nearly equal to that in (c). Taking this point into account, the solid arrow “Phase T3b” is depicted almost parallel to the abscissa, in accordance with the observation of the “extreme reduction of rate of tension recovery” noted at the beginning of this section. In the case of Δ
y >
ycdev, as discussed above, it is plausible that the red area significantly decreases from
Figure 12(c) to (d). Accordingly, the dashed arrow “Phase T3b” is depicted downward, in accordance with the observation of the “reversal of rate of tension recovery ” noted at the beginning of this section. After Phase T3b, the tension
T approaches
T0obs by the filament sliding as shown by the arrows “Phase T4”. The above argument on the solid and dashed arrows “Phase T3b” suggests that the reversal of rate of tension recovery occurs when |Δ
Lhs| is large. Supporting this conclusion,
Figure 1 of the paper by Julian and Sollins [
19] shows that the reversal of the rate occurs when |Δ
Lhs| is large.
Ford
et al. [
16] reported experimental data on variation of
T/
T0obs during 0–9 ms, as cited by green circles in
Figure 14. The range of Δ
Lhs was +1.5∼−6.0 nm in their experiment. Since our model is not applicable for positive Δ
Lhs, the case of +1.5 nm is omitted in
Figure 14. Now we shall try to reproduce these results by calculation.
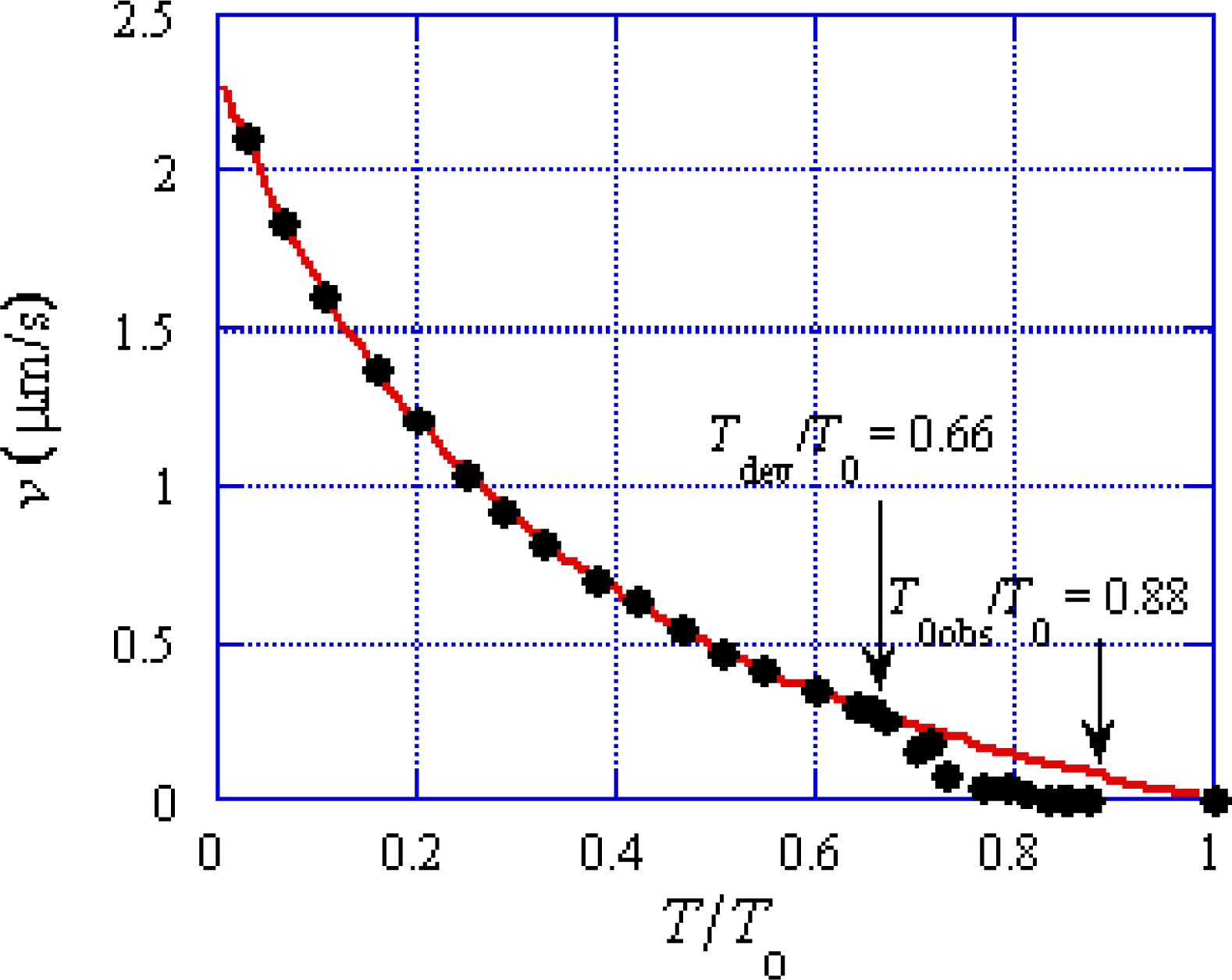
In
Figure 14, values of
T/
T0obs quickly increase within about 1 ms and then gradually approach to the value at 9 ms. The mean time duration needed for one
U* transition is longer than 10 ms for
T/
T0 > 0.5 in
Figure 5. Accordingly, effect of
U* transition is not considered. The experimental length changes | Δ
Lhs | are smaller than 7.0 nm as seen in
Figure 14. Hence, the following discussion is done referring to the solid arrows which are in this | Δ
Lhs | range. The tension changes in Phases T1, T2 and T3a are denoted as
T1,
T2 and
T3a, respectively. Also symbols
T12 and
T123a are defined as follows:
According to the scheme in
Figure 13, the change of
T1 is given by the green curve, and
T2 changes from the green curve to the brown curve. In
Figure 14, the origin of time
t is set equal to 0 when the change of Phase T1 finishes. The parameters
Cini2 and
C fin2 are defined as the initial and final values of
T2/
T0obs, which are given by the green and brown curves, respectively in
Figure 13. If
T2 is approximately expressed by a single decay constant
τT2,
T12/
T0obs is given by,
The constant
τT2 is related with the elastic change of the actin filament and is independent of Δ
Lhs. The solid arrow “Phase T3b” is drawn between the brown curve and the blue curve in
Figure 13, so that the magnitude of tension of
T/
T0obs in T3b is given by the difference between these curves. It is uncertain when the
U*
12 transition and thus Phase T3a start. To make calculation simple, it is assumed that it starts at the same moment as Phase T2,
i.e., at
t = 0. Also the change of
T3a is approximately expressed with a single decay constant
τT3a:
where (
Cini3a −
Cfin3a) represents the magnitude of the tension variation.
Cini3a and
Cfin3a are given by the brown and blue curves, respectively in
Figure 13.
Now the problem is how the parameter
τT3a changes with Δ
Lhs. In the discussion on the force-velocity relation in [
2], the average time
tc for a myosin head at
yc to cross over the potential barrier
U*(
yc) is expressed as
tc = (1/
A) exp (
U*(
yc)/
kT) (Equations 4–2–1 in [
2]), and
U*(
yc) is expressed by
U*(
yc) =
U*
0 −
byc (Equations 4–2–2 in [
2]). These formulae give
tc =
B exp (−
cyc) where
B and
c =
b/
kT are constants. In
Figure 11(b) the boundary between the violet and red areas is indicated as −
L +
ycT2 +
x0. The relative relation between this boundary and A1 is similar to that between
yc and A2. Then, in analogy to the above relation, with constants
B’ and
c’, the relation
T3a =
B’ exp (−
cyc’) is expected as an approximate expression, where
yc’ = −
L +
ycT2 +
x0 (
Figure 11(b)). Then, as
L and
x0 are constants,
τT3a in
Equation 36 is approximately given by
τT3a =
Bexp(−
c’ycT2), where
c’ is constant. Since
ycT2 = −Δ
Lhs/4.6 (
Equation 32), this relation can be rewritten as,
where
A3a and
c3a are constants. This relation implies that
τT3a becomes small and the tension variation becomes fast when | Δ
Lhs | increases.
Trial calculations were done for
T123a =
T12 +
T3a (
Equation 34) to explain the experimental observations by changing parameters
τT2,
A3a and
c3a in
Equations 35,
36 and
37. Fairly good agreement with the experimental data is obtained as shown in
Figure 14 by using the following parameter values:
In Section 4, it is assumed that the elastic change of the actin filament occurs 0.4–1 ms after the length changes of the crossbridge and myosin filament. The time constant
τT2 = 0.7 ms is in consistency with this assumption.
6. Isotonic Velocity Transient
Isotonic velocity transients were studied by Civan and Podolsky [
17], Huxley
et al. [
15] and Sugi and Tsuchiya [
22,
23]. A muscle was stimulated and initially held at a constant length. It was then released suddenly and allowed to shorten under a constant load. In this section, discussion will refer to the experimental data presented in
Figure 3 of the article by Civan and Podolsky [
17]. Responses of muscle to the sudden load change are classified into four Phases by Huxley [
15]. Analogously, we use terminology “Phase Vn” as mentioned in Section 4. While Phase number of Huxley is related with time sequence of the length changes, Phase Vn is more related with molecular processes rather than the time sequence. Phase V1 is the length change which simultaneously occurs with load change. Phase V2 is rapid early shortening. Phase V3 is extreme reduction or reversal of shortening speed. Phase V4 is responsible to the fact that the filament sliding velocity temporarily becomes larger than the steady value. Phases V2, V3 and V4 overlap each other in their time courses.
Figure 15 shows changes of the
ρ distribution in these four Phases.
Figure 15 (a) is the
ρ distribution just after Phase V1. Since
yc = 0 at the isometric tension (cf.
Figure 10 (a)),
yc in
Figure 15 (a) is equal to the variation from the isometric tension, Δ
ycV1 (cf.
Figure 10 (c)). We shall discuss the case of Δ
ycV1 >
ycdev, since most experiments on the velocity transient were done for Δ
yc >
ycdev.
In
Figure 15, the vertical red arrows show time courses of Phases V2, V3 and V4. They overlap each other indicating the overlap of their time courses. The width of the violet area
x0 in
Figure 15(a) is the same as in the isometric tension since the
U*
12 transition does not occur yet. Then the
U*
12 transition starts,
i.e., Phase V2 starts. As mentioned above, there is a time delay in occurrence of the
U* transition and thus of the filament sliding.
Figure 15 (b1) shows the state in midway of Phase V2, where the
U* transition starts. In (b1), a portion of the violet area has turned into red due to the
U*
12f transition and internal stress increases, which causes shrinkage of the
ρ distribution, and thus of the muscle. At (b2), the
U*
12f transition is over and all the violet area is turned into the red area as Δ
ycV1 >
ycdev. The process from (a) to (b2) is related with the
U*
12 transition and is called Phase V2, where the red area increases and the internal stress increases. The width of
ρ distribution changes from
L in (a) to
L-Δ
L in (b2). Accordingly, a fast shortening of the muscle is expected.
During Phase V2 the U* transition starts at (b1). There is a mutual interaction between the U* transition and the internal stress. The U* transition causes the filament sliding, releases the internal stress and tends to make yc larger. The internal stress tends to expand the ρ distribution, increases yc and accelerates the U* transition. The former process is called Phase V3. The latter process is called Phase V4. The mutual interaction almost disappears leaving a large value of yc at (c). Then the frequency of the U* transition and yc change toward their steady values from (c) to (d), where the ρ distribution at the steady filament sliding is shown. Phases V3 and V4 occur in parallel starting with the U* transition at (b1) and end near (d).
Figure 16 illustrates changes in Phase V1∼V4 in another way. This figure is drawn considering the case of small
T/
T0obs. (An example of the experimental data for small
T/
T0obs can be seen in
Figure 18 (c) given later where Δ
T/
T0obs = 0.87,
i.e.,
T/
T0obs = 0.13.) The red, blue and brown curves are the same as the curves of the same colors in
Figure 13. The black arrows indicate an example of changes of Δ
Lhs in the four Phases. As discussed above, there is the overlap between the Phases, and the arrow “Phase Vn” indicates the Phase which mainly contributes to the process.
The length change in Phase V1 occurs along the brown curve, since the elastic changes of the crossbridge, the myosin filament and the actin filament contribute to this change. The arrow “Phase V2” corresponds to the rapid decrease of muscle length from
Figure 15 (a) to (b2). The length of the arrow “Phase V2” is tentatively depicted as 6 nm (the same order of magnitude of the rapid drop of 5 nm in
Figure 18 (c) given later). As discussed above, the time courses of Phases 3 and 4 overlap each other. The direction of arrow “Phase V3” is reversed, representing the muscle elongation from
Figure 15 (b2) to (c). Through “Phase 4”, the system approaches to the red curve corresponding to the change from (c) to (d) in
Figure 15. The green arrow “
Lhsv” symbolically represents the steady filament sliding corresponding to the state in
Figure 15 (d).
Now let us numerically reproduce the experimental data presented in
Figure 3 of the paper by Civan and Podolsky [
17]. The length change per half sarcomere at the steady filament sliding is denoted as
Lhsv which corresponds to the dashed line in
Figure 3 of [
17]. Discussion will be done referring to
Lhsv expressed by
Here
v is the sliding velocity determined by the experimental data in [
17].
The length change caused by the
U*
12 transition in Phase V2 is denoted as
Lhs2. The speed of
Lhs2 depends upon the frequency of
U*
12 transition and will be large at the beginning and gradually decay. Its time course is approximately expressed with decay constant
τV2 by
where
BV2 is a constant.
As discussed above, there is the mutual interaction between the U* transition and the internal stress. (a) The U* transition causes the filament sliding which releases the internal stress. (b) The internal stress tends to expand the ρ distribution, pushes yc forward and accelerates the U* transition. The interaction (a) is discussed first. The interaction (b) will be discussed later in relation with Phase 4.
The interaction (a) releases the internal stress and thus elongates the muscle. As shown in
Figure 15, there is an overlap between Phase V2 and V3, and thus there is an overlap between length change
Lhs2 and the length change due to the interaction (a). The combined length change is denoted as
Lhs23, which is expressed by multiplying
Lhs2 by exp(−
t/
τV3):
An example of
Lhs23 is shown by the dotted black curve in
Figure 17 to demonstrate its characteristics, together with
Lhsv (the blue line).
The decay times
τV2 and
τV3 should be functions of
T/
T0obs or
v. The magnitude of
τV2 is proportional to the time duration of occurrence of the
U*
12 transition. Analogously to the manner to derive
Equation 37, the relation
τT3a =
Bexp (−
cyc’) is expected as an approximate expression, where
yc’ is the
y at the boundary between the violet area and red area in
Figure 15(a), where
yc’ = −L + Δ
ycV1+
x0. The tension
T becomes smaller as Δ
ycV1 becomes larger. As a simple approximation, Δ
T =
T0obs −
T is set proportional to Δ
ycV1. Then the decay time
τV2 is given approximately by
where
aV2 and
cV2 are constants. The time constant
τV3 in Phase V3 is related with the
U* transition, and its magnitude seems to be an order of
τstep =
L/
v (
Equation 5). Hence
τV3 is set as
where
bV3 is a constant.
To see the characteristics of combination of
Lhsv and
Lhs23,
Lhs23v is defined by
An example of
Lhs23v is illustrated by the dashed violet curve in
Figure 17.
Now let us consider about the part (b) of the interaction that the internal stress tends to expand the
ρ distribution, pushes
yc forward and accelerates the
U* transition. This effect causes muscle elongation. In
Figure 15, this effect is illustrated by depicting
ycVc in (c) larger than
ycVd in (d). In Phase 4,
ycVc changes into
ycVd and the filament sliding approaches to the steady value. As shown by the overlap of the arrows of Phases V3 and V4 in
Figure 15, these effects overlap with each other. The length change in Phase V4 is denoted as
Lhs4. In analogy to the expression of
Lhs23 (
Equation (41)),
Lhs4 is approximately expressed by
The characteristics of
Lhs4 expressed by this equation are illustrated by the dashed green curve in
Figure 17. The internal stress is zero at
t = 0 and increases by the term {1 − exp(−
t/
τV4)}
2. The term exp(−
t/
τV5) corresponds to the decrease of the muscle length due to the filament sliding. Concerning the magnitude of
BV4, it is plausible that the internal stress pushes
yc forward more effectively when the internal stress rapidly increases,
i.e., when the frequency of the
U*
12 transition rapidly increases. The frequency of the
U*
12 transition is given by the reciprocal of the duration
τV2 given by
Equation 42. Thus
Bv4 is expressed by
where
Gv4 is a constant. The time constants
τV4 and
τV5 will be mainly related with the
U* transitions, and in analogy to
Equation 43 they are expressed as
where
b4 and
b5 are constant.
Since the sum of
Lhsv,
Lhs23 and
Lhs4 is the total length change, it is denoted as Δ
Lhs’
Note that Δ
Lhs’ is the length change from the moment when Phase 1 finishes, while the abscissa Δ
Lhs in
Figure 16 includes the length change in Phase 1.
Time course of Δ
Lhs’ was calculated with various trial values of the parameters looking for good agreement with the experimental data in the cases of (
T0obs −
T)/
T0obs = 0.22, 0.44 and 0.87. Calculation results with the following parameter values are shown in
Figure 18.
Characteristic features of the series of experimental data are fairly well reproduced by the calculation. The drastic change of the curve shape for different Δ
T/
T0obs is mainly due to the
T dependence of the frequency of the occurrence of the
U*
12 transition,
i.e., the relation
τV2 =
aV2exp(
cV2T/
T0obs) (
Equation 42).
8. Summary and Discussion
In our previous paper [
2] (the first part of this Remarks series), difficulty of the power stroke model is pointed out and a new model is proposed to avoid the difficulty. In the model, it is proposed that about 41% of the myosin heads are bound to actin filament and each bound head forms complex MA
3 with three actin molecules. The complex MA
3 translates along the actin filament changing its partner actin molecules in cooperation with
U* transition. This model well explains the properties in the steady filament sliding such as the tension-dependence of the muscle stiffness, the Hill-type force velocity relation and the tension-dependence of energy liberation rate. In the present paper, the isometric tension transient and isometric velocity transient are studied based upon the model. Statistical ensemble of crossbridges is considered and the binding probability density
ρ is introduced. On rectangular
ρ approximation, the edge of the rectangle,
yc determines dynamic properties of muscle in cooperation with
U* and
U*
12 transitions. The internal structure of the MA
3 complex becomes temporally unstable by the sudden length change or by the sudden load change. The complex muscle behaviors observed in these transients are related with the process that the disturbed internal structure returns to its stationary state.
Results reported in the present paper are summarized as follows.
The tension variations in the first Phases in the isometric tension transient (Ford
et al. [
16]) and the isotonic velocity transient (Civan and Podolsky [
17]) are well explained as shown in
Figure 9.
Ratios of extensibilities of crossbridge, myosin filament and actin filament are estimated as 0.22, 0.26 and 0.52 (
Equation 30), in reasonable agreement with the approximate values (0.31, 0.27, 0.42) determined by X-ray diffraction by Wakabayashi
et al. [
21].
The experimental data on the isometric tension transient reported by Ford
et al. [
16] are fairly well explained as shown in
Figure 14.
The characteristic features of muscle in the isotonic velocity transient observed by Civan and Podolsky [
8] are fairly well explained as shown in
Figure 18.
The deviation from the Hill-type force-velocity relation observed by Edman [
12] is reproduced as shown in
Figure 19(b).
It should be noted that the above-mentioned agreements between experimental data and calculation results are obtained by using the muscle stiffnesses
κf and
κb determined in [
2], whose numerical values are given in
Appendix.
The obtained results suggest that the ideas of the ensemble of crossbridges and the rectangular ρ approximation are useful tools in theoretical studies of muscle contraction.
In
Figure 8, tangent of the
T*/
T0 vs. yc* relation (the blue curve) is 0 at
yc* = 0. Accordingly, small fluctuation of tension around
T0obs can produce significant variation of the muscle length. In this connection, the spontaneous oscillatory contraction (SPOC) of muscle (cf. the review by Ishiwata and Yasuda [
24]) seems interesting. As mentioned in [
24], it is a possibility that SPOC has some relation with the activities of cardiac muscles (cf. [
25]).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}


















