Zebrafish Cyclin-Dependent Protein Kinase–Like 1 (zcdkl1): Identification and Functional Characterization

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Identification, Genomic Structure, and Synteny Analysis of zcdkl1
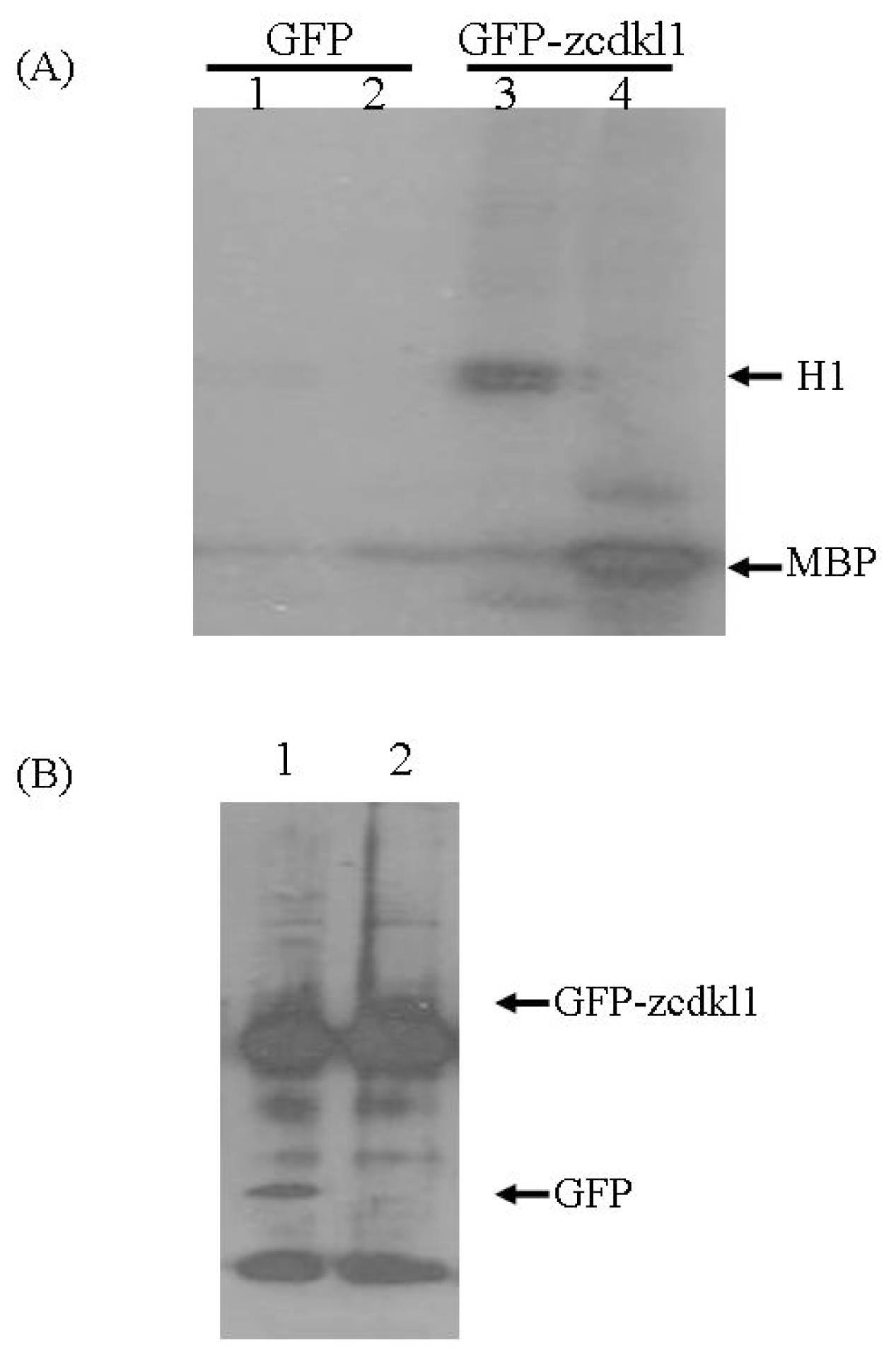
2.2. zcdkl1 Phosphorylates Myelin Basic Protein and Histone H1
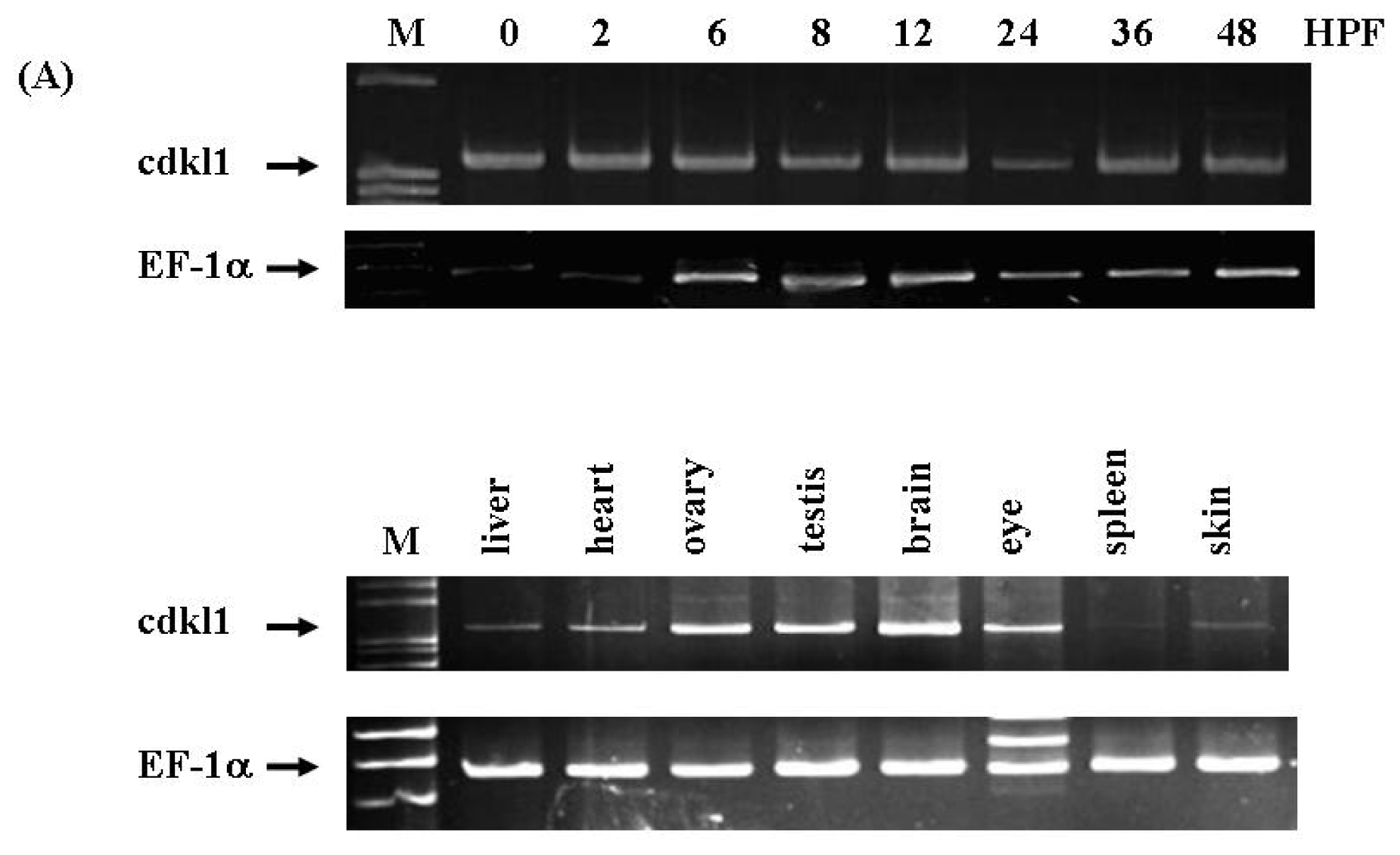
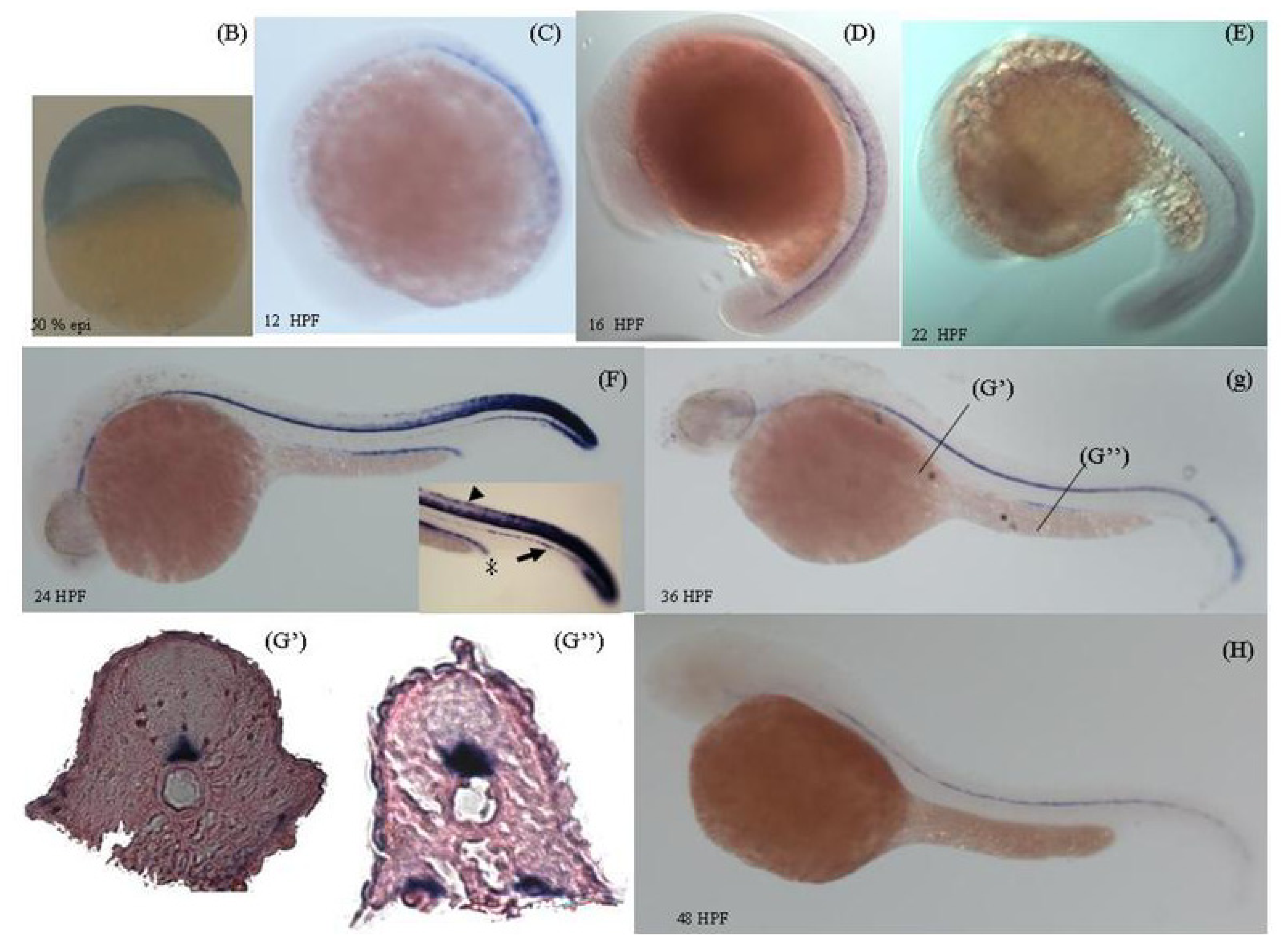
2.3. Temporal and Spatial Expression Pattern of zcdkl1 RNA Transcript in Developmental and Adult Tissues
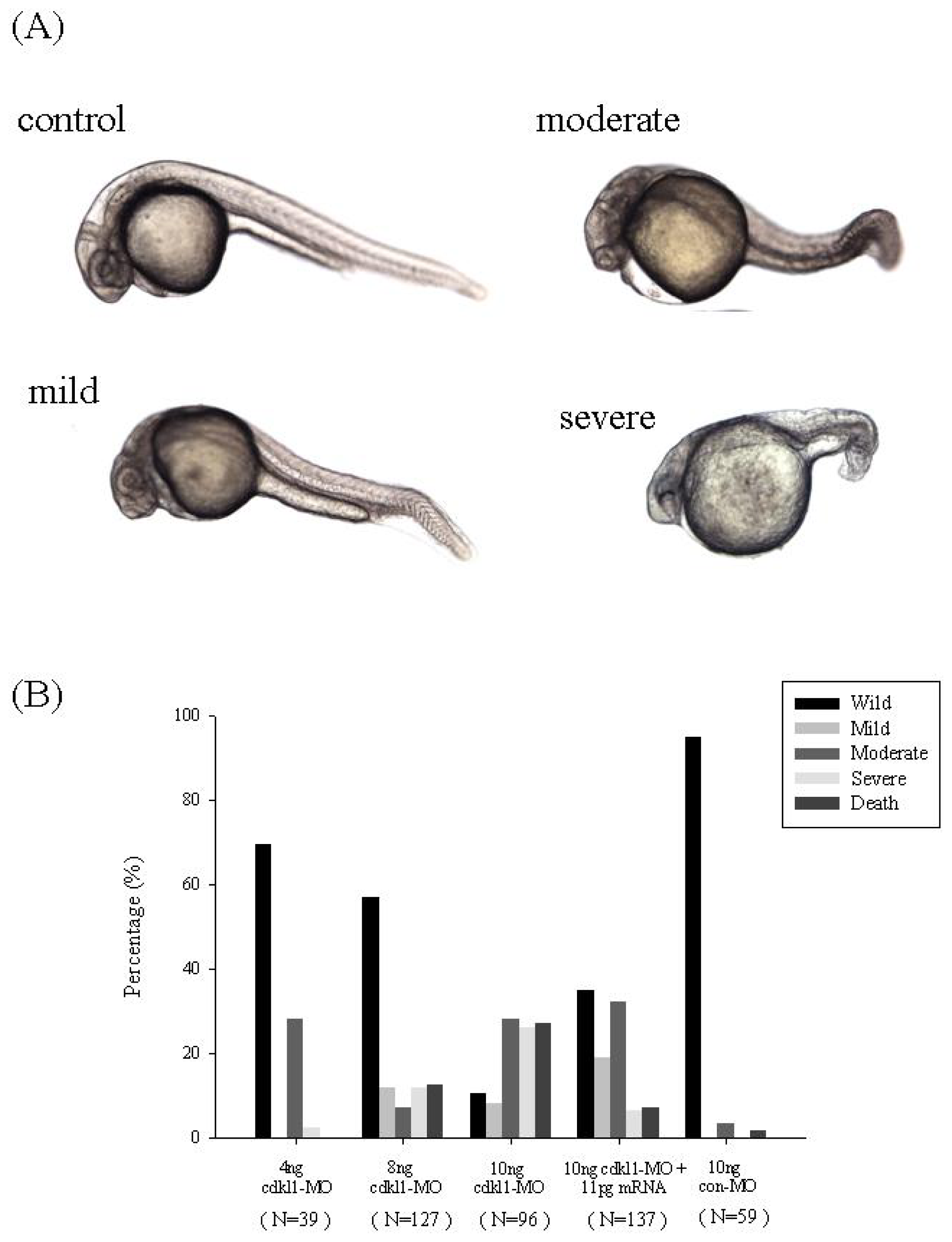
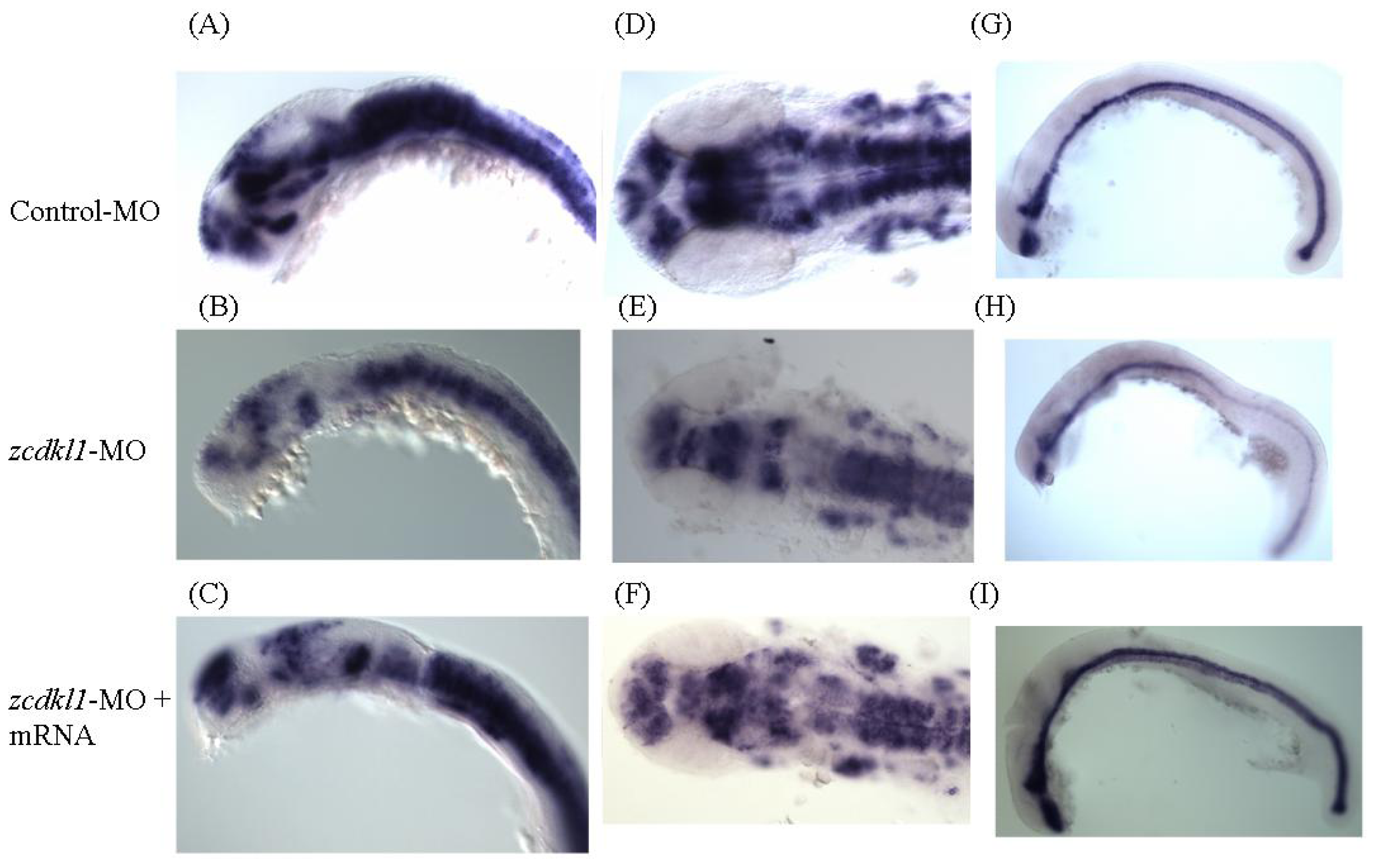
2.4. Knockdown of zcdkl1 Induced Malformation of Zebrafish
3. Experimental Section
3.1. Maintenance of Zebrafish
3.2. RT-PCR of zcdkl
3.3. Plasmid Construction
3.4. Whole Mount in situ Hybridization
3.5. Morpholino Injection and mRNA Rescue
3.6. Cell Culture and Transfection
3.7. Immunoprecipitation and Kinase Activity of zcdkl1
3.8. Western Blot Analysis
4. Discussion
5. Conclusion
Acknowledgments
References
- Morgan, DO. Principles of CDK regulation. Nature 1995, 374, 131–134. [Google Scholar]
- Pines, J; Hunter, T. Isolation of a human cyclin cDNA: Evidence for cyclin mRNA and protein regulation in the cell cycle and for interaction with p34cdc2. Cell 1989, 58, 833–846. [Google Scholar]
- Pines, J; Hunter, T. p34cdc2: The S and M kinase? New Biol 1990, 2, 389–401. [Google Scholar]
- Meyerson, M; Enders, GH; Wu, CL; Su, LK; Gorka, C; Nelson, C; Harlow, E; Tsai, LH. A family of human cdc2-related protein kinases. EMBO J 1992, 11, 2909–2917. [Google Scholar]
- Taglienti, CA; Wysk, M; Davis, RJ. Molecular cloning of the epidermal growth factor-stimulated protein kinase p56 KKIAMRE. Oncogene 1996, 13, 2563–2574. [Google Scholar]
- Yen, SH; Kenessey, A; Lee, SC; Dickson, DW. The distribution and biochemical properties of a Cdc2-related kinase, KKIALRE, in normal and Alzheimer brains. J. Neurochem 1995, 65, 2577–2584. [Google Scholar]
- Kim, SO; Katz, S; Pelech, SL. Expression of second messenger- and cyclin-dependent protein kinases during postnatal development of rat heart. J. Cell. Biochem 1998, 69, 506–521. [Google Scholar]
- Chen, E; Ekker, SC. Zebrafish as a genomics research model. Curr. Pharm. Biotechnol 2004, 5, 409–413. [Google Scholar]
- Westerfield, M; Doerry, E; Kirkpatrick, AE; Driever, W; Douglas, SA. An on-line database for zebrafish development and genetics research. Semin. Cell Dev. Biol 1997, 8, 477–488. [Google Scholar]
- Kimmel, CB; Ballard, WW; Kimmel, SR; Ullmann, B; Schilling, TF. Stages of embryonic development of the zebrafish. Dev. Dyn 1995, 203, 253–310. [Google Scholar]
- Hsu, LS; Chen, GD; Lee, LS; Chi, CW; Cheng, JF; Chen, JY. Human Ca2+/calmodulin-dependent protein kinase kinase beta gene encodes multiple isoforms that display distinct kinase activity. J. Biol. Chem 2001, 276, 31113–31123. [Google Scholar]
- Placzek, M; Briscoe, J. The floor plate: multiple cells, multiple signals. Nat. Rev. Neurosci 2005, 6, 230–240. [Google Scholar]
- Strahle, U; Lam, CS; Ertzer, R; Rastegar, S. Vertebrate floor-plate specification: Variations on common themes. Trends Genet 2004, 20, 155–162. [Google Scholar]
- Chiang, C; Litingtung, Y; Lee, E; Young, KE; Corden, JL; Westphal, H; Beachy, PA. Cyclopia and defective axial patterning in mice lacking Sonic hedgehog gene function. Nature 1996, 383, 407–413. [Google Scholar]
- Roelink, H; Porter, JA; Chiang, C; Tanabe, Y; Chang, DT; Beachy, PA; Jessell, TM. Floor plate and motor neuron induction by different concentrations of the amino-terminal cleavage product of sonic hedgehog autoproteolysis. Cell 1995, 81, 445–455. [Google Scholar]
- Ding, Q; Motoyama, J; Gasca, S; Mo, R; Sasaki, H; Rossant, J; Hui, CC. Diminished Sonic hedgehog signaling and lack of floor plate differentiation in Gli2 mutant mice. Development 1998, 125, 2533–2543. [Google Scholar]
- Marti, E; Bumcrot, DA; Takada, R; McMahon, AP. Requirement of 19K form of Sonic hedgehog for induction of distinct ventral cell types in CNS explants. Nature 1995, 375, 322–325. [Google Scholar]
- Krauss, S; Concordet, JP; Ingham, PW. A functionally conserved homolog of the Drosophila segment polarity gene hh is expressed in tissues with polarizing activity in zebrafish embryos. Cell 1993, 75, 1431–1444. [Google Scholar]
- Jeong, Y; Epstein, DJ. Distinct regulators of Shh transcription in the floor plate and notochord indicate separate origins for these tissues in the mouse node. Development 2003, 130, 3891–3902. [Google Scholar]
- Qian, X; Costa, RH. Analysis of hepatocyte nuclear factor-3 beta protein domains required for transcriptional activation and nuclear targeting. Nucleic Acids Res 1995, 23, 1184–1191. [Google Scholar]
- Ota, M; Ito, K. Induction of neurogenin-1 expression by sonic hedgehog: Its role in development of trigeminal sensory neurons. Dev. Dyn 2003, 227, 544–551. [Google Scholar]






© 2011 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Hsu, L.-S.; Liang, C.-J.; Tseng, C.-Y.; Yeh, C.-W.; Tsai, J.-N. Zebrafish Cyclin-Dependent Protein Kinase–Like 1 (zcdkl1): Identification and Functional Characterization. Int. J. Mol. Sci. 2011, 12, 3606-3617. https://doi.org/10.3390/ijms12063606
Hsu L-S, Liang C-J, Tseng C-Y, Yeh C-W, Tsai J-N. Zebrafish Cyclin-Dependent Protein Kinase–Like 1 (zcdkl1): Identification and Functional Characterization. International Journal of Molecular Sciences. 2011; 12(6):3606-3617. https://doi.org/10.3390/ijms12063606
Chicago/Turabian StyleHsu, Li-Sung, Cyong-Jhih Liang, Chen-Yuan Tseng, Chi-Wei Yeh, and Jen-Ning Tsai. 2011. "Zebrafish Cyclin-Dependent Protein Kinase–Like 1 (zcdkl1): Identification and Functional Characterization" International Journal of Molecular Sciences 12, no. 6: 3606-3617. https://doi.org/10.3390/ijms12063606
APA StyleHsu, L. -S., Liang, C. -J., Tseng, C. -Y., Yeh, C. -W., & Tsai, J. -N. (2011). Zebrafish Cyclin-Dependent Protein Kinase–Like 1 (zcdkl1): Identification and Functional Characterization. International Journal of Molecular Sciences, 12(6), 3606-3617. https://doi.org/10.3390/ijms12063606