Molecular Characterization and Expression Pattern of Tripartite Motif Protein 39 in Gallus gallus with a Complete PRY/SPRY Domain

Abstract
:1. Introduction
2. Results and Discussion
2.1. Defining the Complete TRIM 39 Gene in Gallus Gallus
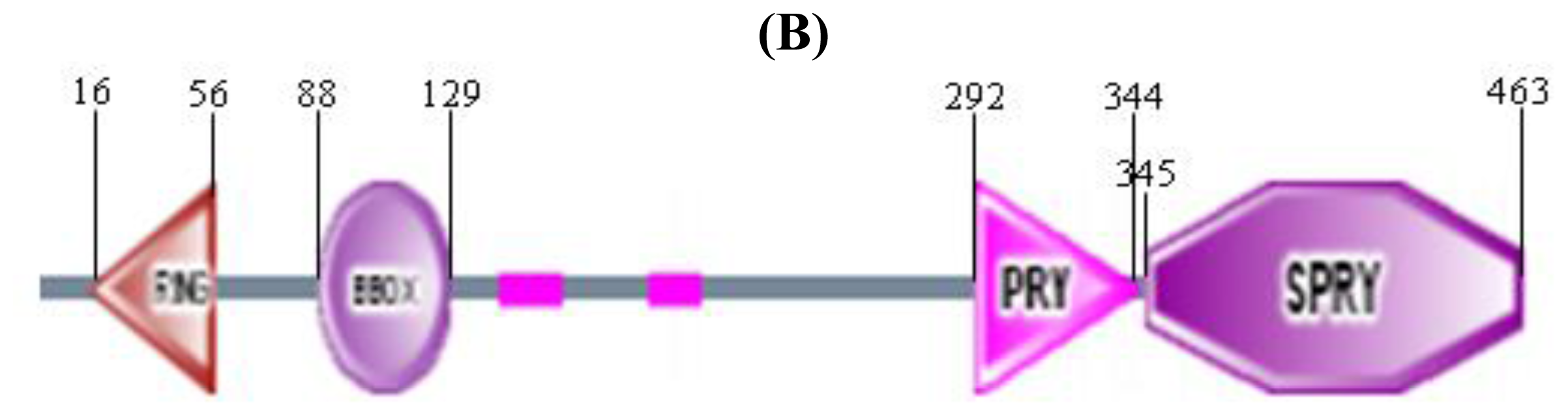
2.2. Domain Composition of Chicken TRIM 39 Protein
2.3. Alignment of Chicken TRIM 39 with Other Species
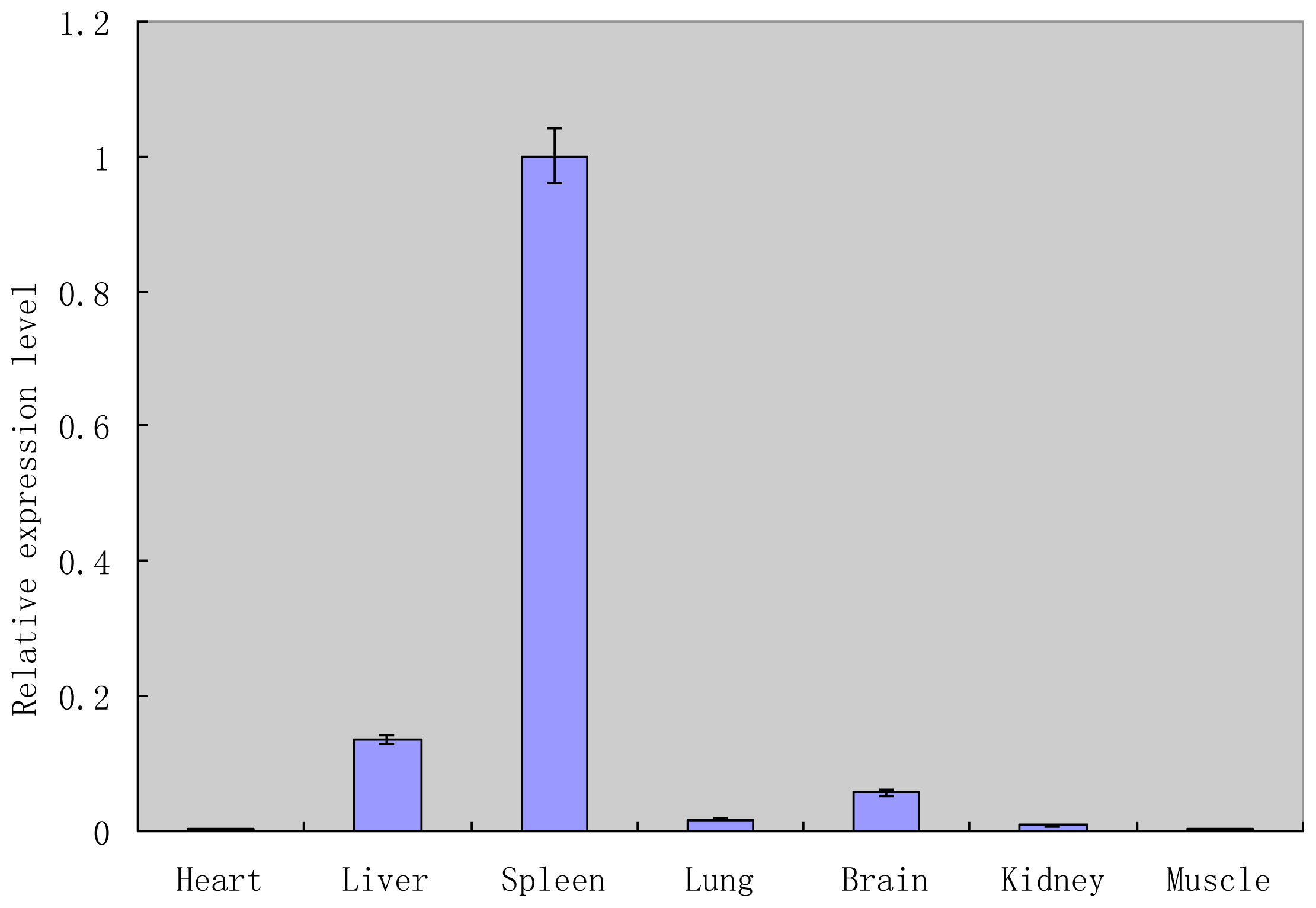
2.4. Tissue Expression Analysis
2.5. Discussion
3. Experimental Section
3.1. Experimental Animal and Reagents
3.2. Total RNA Isolation
3.3. Reverse Transcription Polymerase Chain Reaction
3.4. Real-Time RT-PCR Assay
3.5. Prediction of Protein Structure and Analysis of Sequence Homology
4. Conclusions
References
- Reymond, A; Meroni, G; Fantozzi, A; Merla, G; Cairo, S; Luzi, L; Riganelli, D; Zanaria, E; Messali, S; Cainarca, S; et al. Ballabio A: The tripartite motif family identifies cell compartments. EMBO J 2001, 20, 2140–2151. [Google Scholar]
- Joazeiro, CAP; Weissman, AM. RING finger proteins: Mediators of ubiquitin ligase activity. Cell 2000, 102, 549–552. [Google Scholar]
- Meroni, G; Diez-Roux, G. TRIM/RBCC, a novel class of ‘single protein ring finger’ E3 ubiquitin ligases. Bioessays 2005, 27, 1147–1157. [Google Scholar]
- Reddy, BA; Kloc, M; Etkin, L. The cloning and characterization of a maternally expressed novel zinc finger nuclear phosphoprotein (xnf7) in Xenopus laevis. Dev. Biol 1991, 148, 107–116. [Google Scholar]
- Miyamoto, K; Nakamura, N; Kashiwagi, M; Honda, S; Kato, A; Hasegawa, S; Takei, Y; Hirose, S. RING finger, B-box, and coiled-coil (RBCC) protein expression in branchial epithelial cells of Japanese eel, Anguilla japonica. Eur. J. Biochem 2002, 269, 6152–6161. [Google Scholar]
- Van der Aa, LM; Levraud, JP; Yahmi, M; Lauret, E; Briolat, V; Herbomel, P; Benmansour, A; Boudinot, P. A large new subset of trim genes highly diversified by duplication and positive selection in teleost fish. BMC Biol 2009, 7, 7. [Google Scholar]
- The C. elegans Sequencing Consortium. Genome sequence of the nematode C. elegans: A platform for investigating biology. Science 1998, 282, 2012–2018.
- Nisole, S; Stoye, JP; Saib, A. Trim family proteins: Retroviral restriction and antiviral defence. Nat. Rev. Microbiol 2005, 3, 799–808. [Google Scholar]
- Uchil, PD; Quinlan, BD; Chan, WT; Luna, JM; Mothes, W. TRIM E3 ligases interfere with early and late stages of the retroviral life cycle. PLoS Pathog 2008, 4, e16. [Google Scholar]
- Meroni, G; Diez-Roux, G. Trim/RBCC, a novel class of ‘single protein ring finger’ E3 ubiquitin ligases. Bioessays 2005, 27, 1147–1157. [Google Scholar]
- Quaderi, NA; Schweiger, S; Gaudenz, K; Franco, B; Rugarli, EI; Berger, W; Feldman, GJ; Volta, M; Andolfi, G; Gilgenkrantz, S; et al. Opitz G/BBB syndrome, a defect of midline development, is due to mutations in a new ring finger gene on Xp22. Nat. Genet 1997, 17, 285–291. [Google Scholar]
- Scaglioni, PP; Pandolfi, PP. The theory of APL revisited. Curr. Top. Microbiol. Immunol 2007, 313, 85–100. [Google Scholar]
- Torok, M; Etkin, LD. Two b or not two b? Overview of the rapidly expanding b-box family of proteins. Differentiation 2001, 67, 63–71. [Google Scholar]
- Barr, SD; Smiley, JR; Bushman, FD. The interferon response inhibits HIV particle production by induction of TRIM22. PLoS Pathog 2008, 4, e1000007. [Google Scholar]
- Shiina, T; Briles, WE; Goto, RM; Hosomichi, K; Yanagiya, K; Shimizu, S; Inoko, H; Miller, MM. Extended gene map reveals tripartite motif, C-type lectin, and IG superfamily type genes within a subregion of the chicken MHC-B affecting infectious disease. J. Immunol 2007, 178, 7162–7172. [Google Scholar]
- Ruby, T; Bed’Hom, B; Wittzell, H; Morin, V; Oudin, A; Zoorob, R. Characterisation of a cluster of TRIM-B30.2 genes in the chicken MHC B locus. Immunogenetics 2005, 57, 116–128. [Google Scholar]
- Ponting, C; Schultz, J; Bork, P. Spry domains in ryanodine receptors (Ca2+-release channels). Trends Biochem. Sci 1997, 22, 193–194. [Google Scholar]
- Vernet, C; Boretto, J; Mattei, MG; Takahashi, M; Jack, LJW; Mather, IH; Rouquier, S; Pontarotti, P. Evolutionary study of multigenic families mapping close to the human MHC class I region. J. Mol. Evol 1993, 37, 600–612. [Google Scholar]
- Bateman, A; Coin, L; Durbin, R; Finn, RD; Hollich, V; Griffiths-Jones, S; Khanna, A; Marshall, M; Moxon, S; Sonnhammer, ELL; et al. The Pfam protein families database. Nucl. Acid. Res 2004, 32, D138–D141. [Google Scholar]
- Rhodes, DA; de Bono, B; Trowsdale, J. Relationship between SPRY and B30.2 protein domains. Evolution of a component of immune defence. Immunology 2005, 116, 411–417. [Google Scholar]
- Henry, J; Mather, IH; McDermott, MF; Pontarotti, P. B30.2-like domain proteins: Update and new insights into a rapidly expanding family of proteins. Mol. Biol. Evol 1998, 15, 1696–1705. [Google Scholar]
- James, LC; Keeble, AH; Khan, Z; Rhodes, DA; Trowsdale, J. Structural basis for pryspry-mediated tripartite motif (TRIM) protein function. Proc. Nat. Acad. Sci. USA 2007, 104, 6200–6205. [Google Scholar]
- ExPASy Proteomics Server (ExPASy). Swiss Institute of Bioinformatics (SIB): Lausanne, Switzerland, 2011. Available online: http://www.expasy.ch/tools/ accessed on 27 April 2011.
- Simple Modular Architecture Research Tool (SMART). Biobyte Solutions GmbH: Berlin, Germany, 2011. Available online: http://smart.embl-heidelberg.de/ accessed on 27 April 2011.







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Primer sequence (5′→3′) |
|---|---|
| Primer 1 | CCGTCGCTATGGATGAAG |
| Primer 2 | GTGGGAGGGTACGCAAAA |
| Primer 3 | CGCGGATCCATGGATGAAGATAACCCAGCTG |
| Primer 4 | ACGCGTCGACCTTTGGGGGCAGAGTGCAG |
| Primer 5 | TGTCCAAACTGTCGCCTCT |
| Primer 6 | ACGCGTCGACGCGGGGACAGCGTGAAGCGG |
| Primer 7 | CTCCTGCTCCCTCTGCTTG |
| Primer 8 | GGTTTCCTCCTGTTCCTCCC |
| Primer 9 | ATGGCATCCAAGGAGTGAGC |
| Primer 10 | CAGAACTGAGCGGTGGTGAA |
| Dog | Human | House mouse | Chimpanzee | Sumatran orangutan | Norway rat | Pig | Chicken (NCBI) | Chicken (Ensembl) | |
|---|---|---|---|---|---|---|---|---|---|
| Human | 91.67% | ||||||||
| House mouse | 95.38% | 91.89% | |||||||
| Chimpanzee | 91.48% | 99.61% | 91.89% | ||||||
| Sumatran orangutan | 96.39% | 93.44% | 96.72% | 93.24% | |||||
| Norway rat | 95.38% | 91.89% | 99.18% | 91.89% | 96.72% | ||||
| Pig | 97.39% | 94.02% | 97.75% | 93.82% | 98.98% | 97.75% | |||
| Chicken (NCBI) | 30.28% | 29.20% | 30.63% | 29.20% | 30.97% | 30.63% | 30.97% | ||
| Chicken (Ensembl) | 38.40% | 36.92% | 39.18% | 36.92% | 38.78% | 39.18% | 39.18% | 73% | |
| Chicken (our study) | 38.80% | 37.31% | 39.39% | 37.12% | 39.18% | 39.59% | 39.59% | 73% | 99.35% |
| Species | NCBI Accession No. | The domain architectures of TRIM39 |
|---|---|---|
| Canis lupus | XP_855659 | RING, BBOX, BBC, PRY, SPRY |
| Homo sapiens | NP_067076 | RING, BBOX, BBC, PRY, SPRY |
| Mus musculus | NP_077788 | RING, BBOX, PRY, SPRY |
| Pan troglodytes | NP_001065263 | RING, BBOX, BBC, PRY, SPRY |
| Pongo abelii | NP_001125160 | RING, BBOX, PRY, SPRY |
| Rattus norvegicus | NP_998727 | RING, BBOX, BBC, PRY, SPRY |
| Sus scrofa | NP_001121951 | RING, BBOX, BBC, PRY, SPRY |
| Gallus gallus (NCBI) | NP_001006196 | RING, BBOX, PRY |
| Gallus gallus (Ensembl) | ENSGALP00000000151 | RING, BBOX, PRY, SPRY |
| Gallus gallus (our study) | RING, BBOX, PRY, SPRY |
© 2011 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Pan, C.; Zhao, H.; Shen, L.; Sheng, J. Molecular Characterization and Expression Pattern of Tripartite Motif Protein 39 in Gallus gallus with a Complete PRY/SPRY Domain. Int. J. Mol. Sci. 2011, 12, 3797-3809. https://doi.org/10.3390/ijms12063797
Pan C, Zhao H, Shen L, Sheng J. Molecular Characterization and Expression Pattern of Tripartite Motif Protein 39 in Gallus gallus with a Complete PRY/SPRY Domain. International Journal of Molecular Sciences. 2011; 12(6):3797-3809. https://doi.org/10.3390/ijms12063797
Chicago/Turabian StylePan, Chunqing, Heng Zhao, Lin Shen, and Jiping Sheng. 2011. "Molecular Characterization and Expression Pattern of Tripartite Motif Protein 39 in Gallus gallus with a Complete PRY/SPRY Domain" International Journal of Molecular Sciences 12, no. 6: 3797-3809. https://doi.org/10.3390/ijms12063797
APA StylePan, C., Zhao, H., Shen, L., & Sheng, J. (2011). Molecular Characterization and Expression Pattern of Tripartite Motif Protein 39 in Gallus gallus with a Complete PRY/SPRY Domain. International Journal of Molecular Sciences, 12(6), 3797-3809. https://doi.org/10.3390/ijms12063797