Genetic Diversity and Differentiation of the Orange-Spotted Grouper (Epinephelus coioides) Between and Within Cultured Stocks and Wild Populations Inferred from Microsatellite DNA Analysis

Abstract
:1. Introduction
2. Materials and Methods
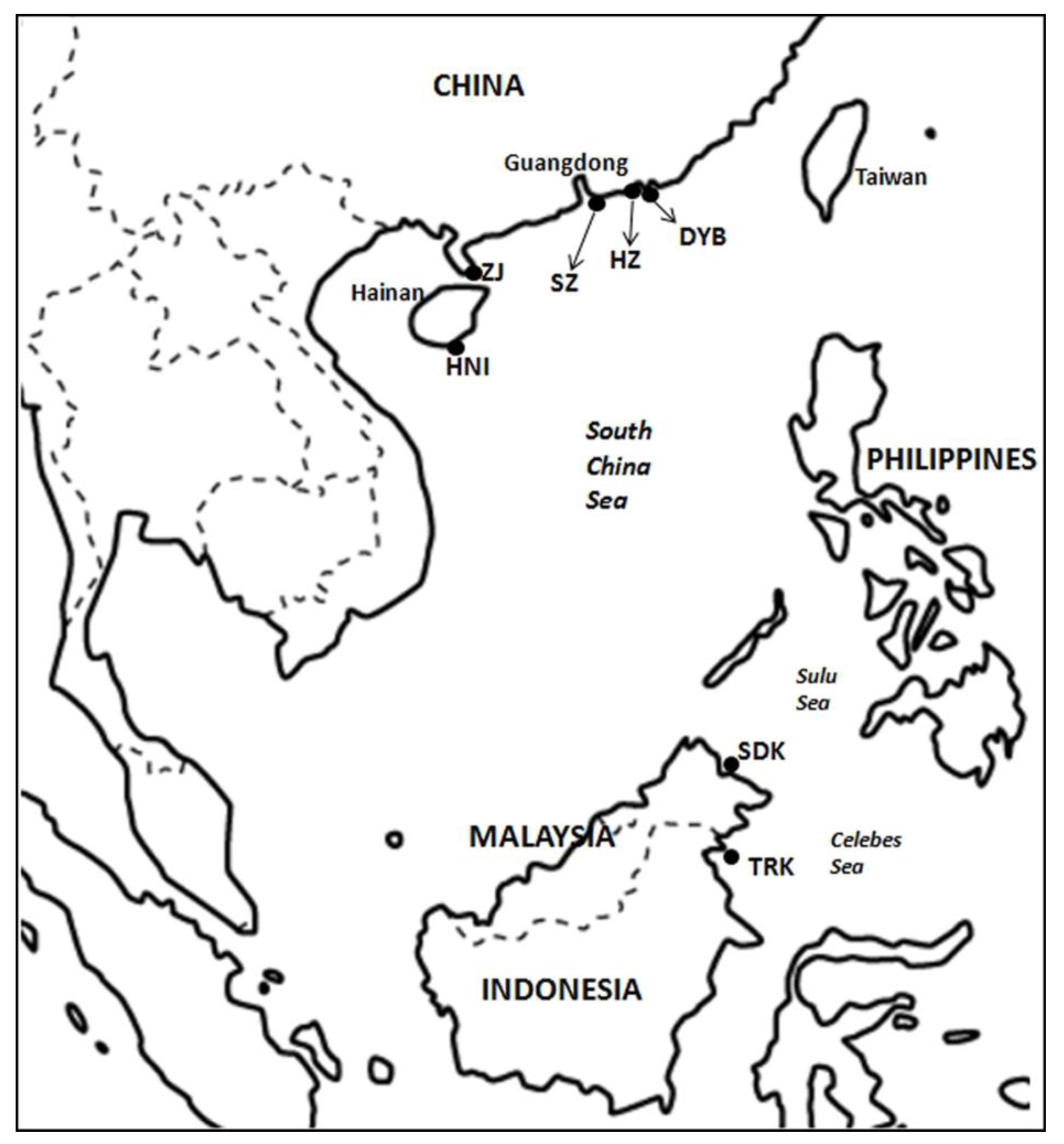
2.1. Sample Collection and DNA Extraction
2.2. Microsatellite Genotyping
2.3. Statistical Analysis
3. Results
3.1. Polymorphisms of Microsatellites
3.2. Intra-Population Genetic Diversity
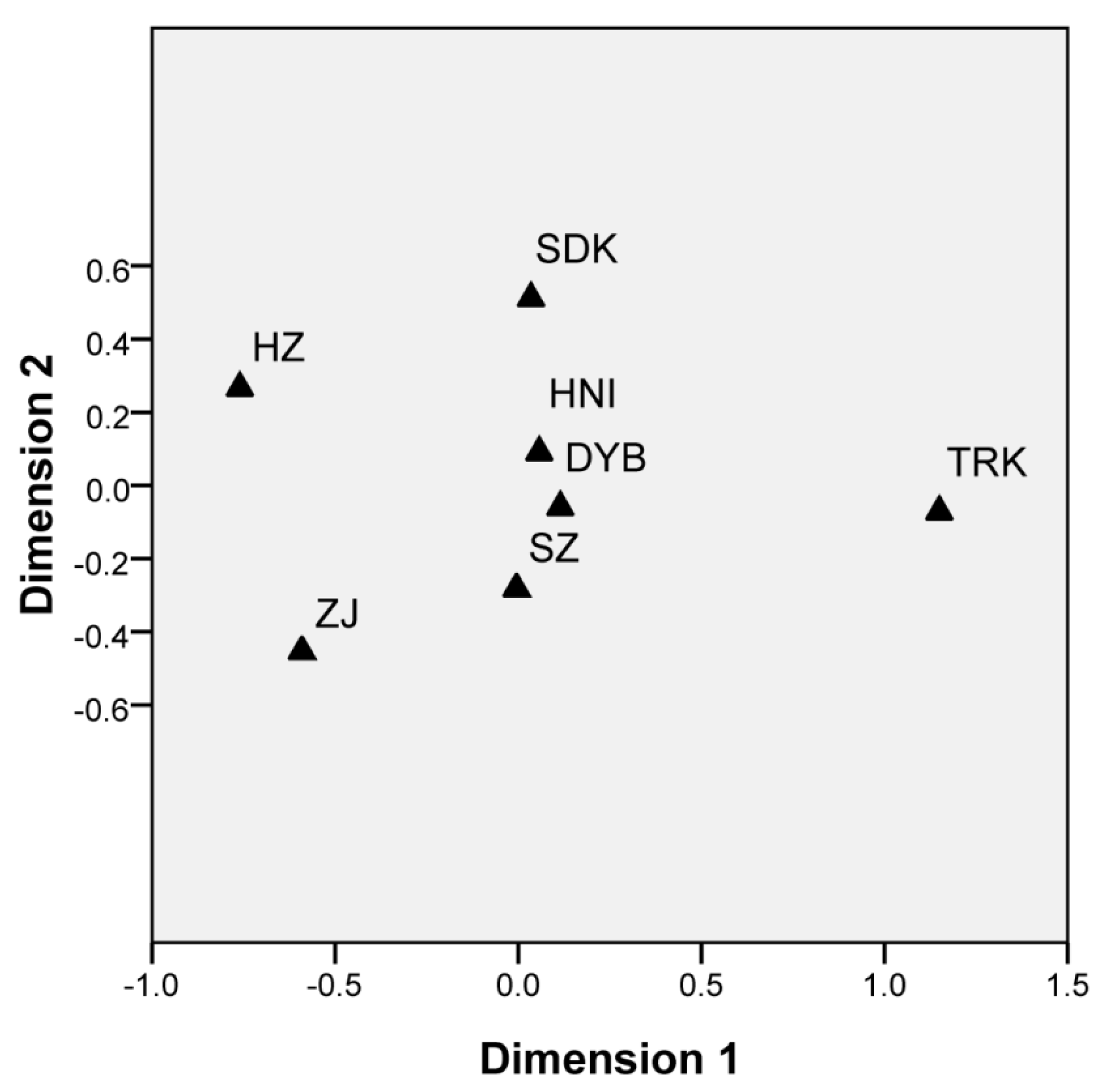
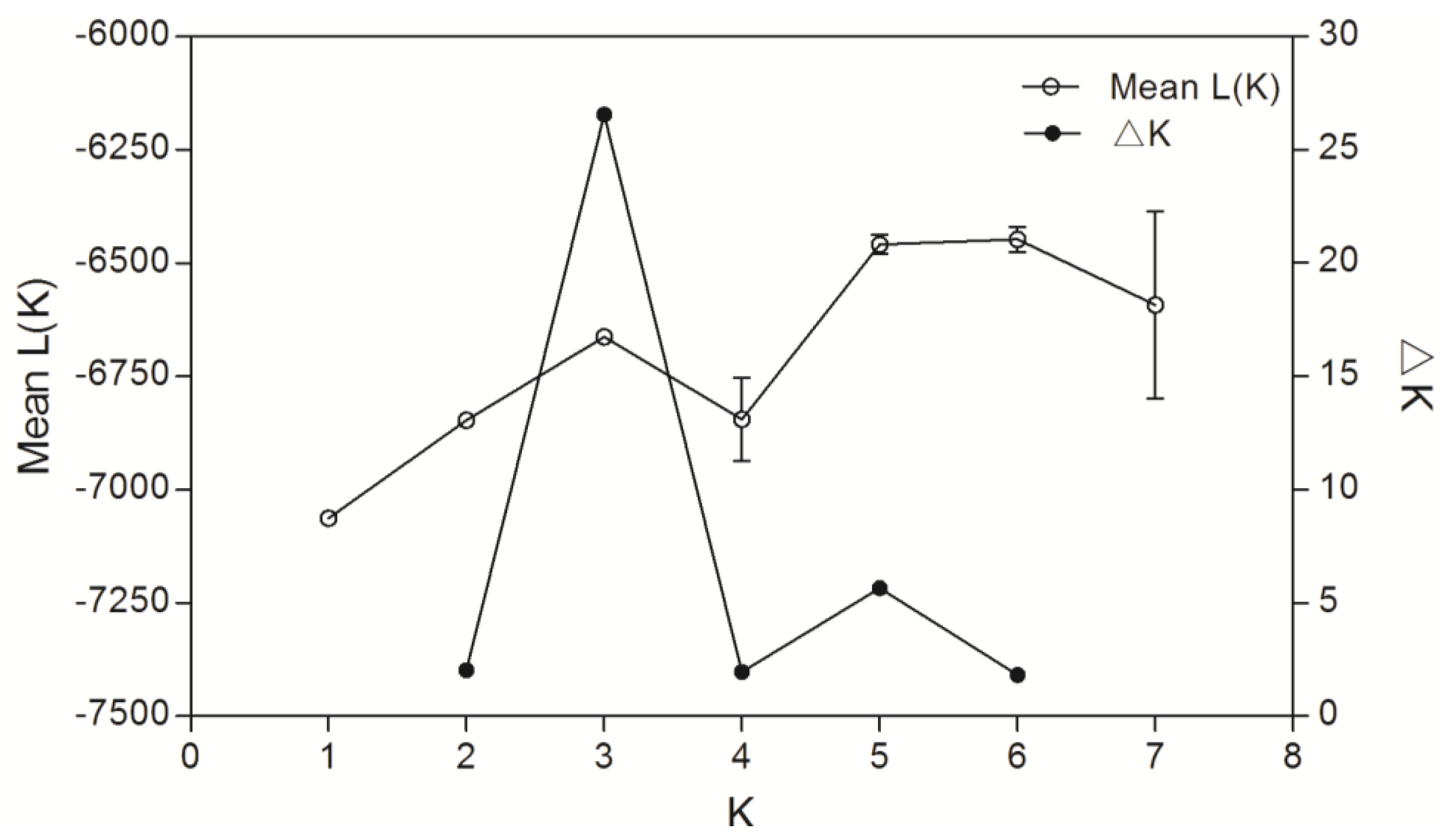
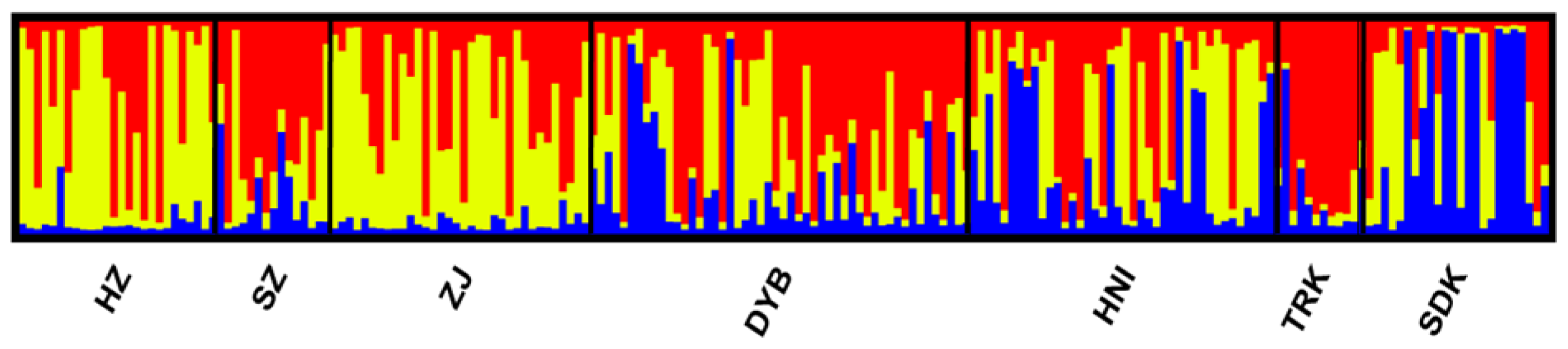
3.3. Inter-Population Genetic Differentiation
4. Discussion
4.1. Microsatellites Polymorphism
4.2. Genetic Variability Between Cultured Stocks and Wild Populations
4.3. Genetic Differentiation Between and Within Cultured Stocks and Wild Populations
5. Conclusions
Acknowledgments
References
- Nelson, JS. Fishes of the World; John Wiley and Sons: New York, NY, USA, 1994. [Google Scholar]
- Heemstra, PC; Randall, JE. FAO Fisheries Synopsis: Groupers of the World (Family Serranidae, Subfamily Epinephelinae); Food and Agriculture Organization of the United Nations: Rome, Italy, 1993; Volume 16, pp. 1–382. [Google Scholar]
- IUCN red list of threatened species; IUCN: Cambridge, UK, 2011. Available online: http://www.iucnredlist.org accessed on 28 June 2011.
- Tupper, M; Sheriff, N. Capture-based aquaculture of groupers. In FAO Fisheries Technical Paper; FAO: Rome, Italy, 2008; pp. 217–253. [Google Scholar]
- FAO, Cultured Aquatic Species Information Programme, Epinephelus coioides; FAO Fisheries and Aquaculture Department: Rome, Italy, 2010. Available online: http://www.fao.org accessed on 28 June 2011.
- Froese, R; Pauly, D. FishBase, version (06/2011); FishBase: Paris, France, 2011. Available online: http://www.fishbase.org accessed on 29 June 2011.
- Allendorf, FW; Phelps, SR. Loss of genetic variation in a hatchery stock of cutthroat trout. Trans. Am. Fish. Soc 1980, 109, 537–543. [Google Scholar]
- Lacy, RC. Loss of genetic diversity from managed populations: Interacting effects of drift, mutation, selection, and population subdivision. Conserv. Biol 1987, 2, 143–158. [Google Scholar]
- Lind, CE; Evans, BS; Knauer, J; Taylor, JJ; Jerry, DR. Decreased genetic diversity and a reduced effective population size in cultured silver-lipped pearl oysters (Pinctada maxima). Aquaculture 2009, 286, 12–19. [Google Scholar]
- Coughlan, JP; Imsland, AK; Galvin, PT; Fitzgerald, RD; Naevdal, G; Cross, TF. Microsatellite DNA variation in wild populations and farmed strains of turbot from Ireland and Norway: A preliminary study. J. Fish. Biol 1998, 52, 916–922. [Google Scholar]
- Norris, AT; Bradley, DG; Cunningham, EP. Microsatellite genetic variation between and within farmed and wild Atlantic salmon (Salmo salar) populations. Aquaculture 1999, 180, 247–264. [Google Scholar]
- Perez-Enriquez, R; Takagi, M; Taniguchi, N. Genetic variability and pedigree tracing of a hatchery-reared stock of red sea bream (Pagrus major) used for stock enhancement, based on microsatellite DNA markers. Aquaculture 1999, 173, 413–423. [Google Scholar]
- Sekino, M; Hara, M; Taniguchi, N. Loss of microsatellite and mitochondrial DNA variation in hatchery strains of Japanese flounder Paralichthys olivaceus. Aquaculture 2002, 213, 101–122. [Google Scholar]
- Lundrigan, TA; Reist, JD; Ferguson, MM. Microsatellite genetic variation within and among Arctic charr (Salvelinus alpinus) from aquaculture and natural populations in North America. Aquaculture 2005, 244, 63–75. [Google Scholar]
- Kohlmann, K; Kersten, P; Flajshans, M. Microsatellite-based genetic variability and differentiation of domesticated, wild and feral common carp (Cyprinus carpio L.) populations. Aquaculture 2005, 247, 253–266. [Google Scholar]
- Antoro, S; Na-Nakorn, U; Koedprang, W. Study of genetic diversity of orange-spotted grouper, Epinephelus coioides, from Thailand and Indonesia using microsatellite markers. Mar. Biotechnol 2006, 8, 17–26. [Google Scholar]
- Wang, J; Guo, F; Ding, S; Wang, J. Genetic diversity of different geographical stocks of Epinephelus coioides by microsatellite DNA. Mar. Sci 2009, 11, 60–64. [Google Scholar]
- Mccusker, MR; Bentzen, P. Historical influences dominate the population genetic structure of a sedentary marine fish, Atlantic wolffish (Anarhichas lupus), across the North Atlantic Ocean. Mol. Ecol 2010, 19, 4228–4241. [Google Scholar]
- Yue, GH; Zhu, ZY; Lo, LC; Wang, CM; Lin, G; Feng, F; Pang, HY; Li, J; Gong, P; Liu, HM; Tan, J; Chou, R; Lim, H; Orban, L. Genetic variation and population structure of Asian seabass (Lates calcarifer) in the Asia-Pacific region. Aquaculture 2009, 293, 22–28. [Google Scholar]
- Meng, X; Wang, Q; Jang, I; Liu, P; Kong, J. Genetic differentiation in seven geographic populations of the fleshy shrimp Penaeus (Fenneropenaeus) chinensis based on microsatellite DNA. Aquaculture 2009, 287, 46–51. [Google Scholar]
- Molecular Ecology Resources Primer Development Consortium. Permanent genetic resources added to molecular ecology resources database, 1 August 2009–30 September 2009. Mol. Ecol. Resour 2010, 10, 232–236.
- Hoelzel, AR; Green, A. Analysis of population-level variation by sequencing PCR-amplified DNA. In Molecular Genetic Analysis of Populations: A Practical Approach; IRL Press: Oxford, UK, 1992; pp. 159–187. [Google Scholar]
- Dong, Q; Liu, C; Guo, Y; Liu, L; Xu, T. Isolation and characterization of polymorphic microsatellite markers in Epinephelus awoara. J. Fish. China 2007, 31, 841–847. [Google Scholar]
- Zatcoff, MS; Ball, AO; Chapman, RW. Characterization of polymorphic microsatellite loci from black grouper, Mycteroperca bonaci (Teleostei: Serranidae). Mol. Ecol. Notes 2002, 2, 217–219. [Google Scholar]
- Zhu, ZY; Lo, LC; Lin, G; Xu, YX; Yue, GH. Isolation and characterization of polymorphic microsatellites from red coral grouper (Plectropomus maculatus). Mol. Eco. Notes 2005, 5, 579–581. [Google Scholar]
- Rivera, M; Graham, GC; Roderick, GK. Isolation and characterization of nine microsatellite loci from the Hawaiian grouper Epinephelus quernus (Serranidae) for population genetic analyses. Mar. Biotechnol 2003, 5, 126–129. [Google Scholar]
- Ramirez, MA; Patricia-Acevedo, J; Planas, S; Carlin, JL; Funk, SM; McMillan, WO. New microsatellite resources for groupers (Serranidae). Mol. Ecol. Notes 2006, 6, 813–817. [Google Scholar]
- Van Oosterhout, C; Hutchinson, WF; Wills, D; Shipley, P. MICRO-CHECKER: Software for identifying and correcting genotyping errors in microsatellite data. Mol. Ecol. Notes 2004, 4, 535–538. [Google Scholar]
- Hered, GJ. FSTAT (Version 1.2): A computer program to calculate F-statistics. J. Hered 1995, 86, 485–486. [Google Scholar]
- Mousadik, AE; Petit, RJ. High level of genetic differentiation for allelic richness among populations of the argan tree [Argania spinosa (L.) Skeels] endemic to Morocco. Theor. Appl. Genet 1996, 92, 832–839. [Google Scholar]
- Sokal, RR; Rohlf, FJ. Biometry: The Principles and Practice of Statistics in Biological Research, 3rd ed; W.H. Freeman and Co: New York, NY, USA, 1995; pp. 427–434. [Google Scholar]
- Rousset, F. GENEPOP'007: A complete re-implementation of the GENEPOP software for Windows and Linux. Mol. Ecol. Resour 2008, 8, 103–106. [Google Scholar]
- Rice, WR. Analyzing tables of statistical tests. Evolution 1989, 43, 223–225. [Google Scholar]
- Cornuet, JM; Luikart, G. Description and power analysis of two tests for detecting recent population bottlenecks from allele frequency data. Genetics 1996, 144, 2001–2014. [Google Scholar]
- Lewis, P; Zaykin, D. Genetic data analysis: Computer program for the analysis of allelic data, Version 1.1; Paul Lewis Phylogenetics: Storrs, CT, USA, 2002. Available online: http://hydrodictyon.eeb.uconn.edu/people/plewis/software.php accessed on 21 June 2011.
- Excoffier, L; Laval, G; Schneider, S. Arlequin ver. 3.0: An integrated software package for population genetics data analysis. Evol. Bioinform 2005, 1, 47–50. [Google Scholar]
- Miller, MP. Tools for Population Genetic Analyses (TFPGA) 1.3: A Windows Program for the Analysis of Allozyme and Molecular Population Genetic Data; Utah State University: Logan, UT, USA, 1997. [Google Scholar]
- Hubisz, MJ; Falush, D; Stephens, M; Pritchard, JK. Inferring weak population structure with the assistance of sample group information. Mol. Ecol. Resour 2009, 9, 1322–1332. [Google Scholar]
- Evanno, G; Regnaut, S; Goudet, J. Detecting the number of clusters of individuals using the software Structure a simulation study. Mol. Ecol 2005, 14, 2611–2620. [Google Scholar]
- Bentzen, P; Taggart, CT; Ruzzante, DE; Cook, D. Microsatellite polymorphism and the population structure of Atlantic cod (Gadus morhua) in the Northwest Atlantic. Can. J. Fish. Aquat. Sci 1996, 53, 2706–2721. [Google Scholar]
- Takagi, M; Taniguchi, N; Cook, D; Doyle, RW. Isolation and characterization of microsatellite loci rom red sea bream Pagrus major and detection in closely related species. Fish. Sci 1997, 63, 199–204. [Google Scholar]
- Nugroho, E; Ferrel, DJ; Smith, P; Taniguchi, N. Genetic divergence of kingfish from Japan, Australia and New Zealand inferred by microsatellite DNA and mitochondrial DNA control region markers. Fish. Sci 2001, 67, 843–850. [Google Scholar]
- Koedprang, W; Na-Nakorn, U; Nakajima, M; Taniguchi, N. Evaluation of genetic diversity of eight grouper species Epinephelus spp. based on microsatellite variations. Fish. Sci 2007, 73, 227–236. [Google Scholar]
- Sun, XW; Liu, CY; Liang, LQ. Isolation of microsatellite with enrichment by magnetic beads in grass carp Ctenopharyngodon idella. J. Fish. China 2005, 29, 286–482. [Google Scholar]
- Koljonen, ML; Tähtinen, J; Säisä, M; Koskiniemi, J. Maintenance of genetic diversity of Atlantic salmon (Salmo salar) by captive breeding programmes and the geographic distribution of microsatellite variation. Aquaculture 2002, 212, 69–92. [Google Scholar]
- Mjolnerod, IB; Refseth, UH; Karlsen, E; Balstad, T; Jakobsen, KS; Hindar, K. Genetic differences between two wild and one farmed population of Atlantic salmon (Salmo salar) revealed by three classes of genetic markers. Hereditas 1997, 127, 239–248. [Google Scholar]
- Allendorf, FW. Genetic drift and the loss of alleles versus heterozygosity. Zoo Biol 1986, 5, 181–190. [Google Scholar]
- Comps, B; Gomory, D; Letouzey, J; Thiebaut, B; Petit, RJ. Diverging trends between heterozygosity and allelic richness during postglacial colonization in the European beech. Genetics 2001, 157, 389–397. [Google Scholar]
- Skaala, O. Genetic population structure of Norwegian brown trout. J. Fish Biol 1992, 41, 631–646. [Google Scholar]
- Allendorf, FW; Ryman, N. Genetic management of hatchery stocks. In Population Genetics and Fishery Management; Ryman, N, Utter, F, Eds.; Washington Sea Grant Program, University of Washington Press: Seattle, WA, USA, 1987; pp. 141–159. [Google Scholar]
- Eknath, AE; Doyle, RW. Indirect selection for growth and life-history traits in Indian carp aquaculture: I. Effects of broodstock management. Aquaculture 1985, 49, 73–84. [Google Scholar]
- Oceanic and Fisheries Administrator of Guangdong Province, Guangzhou, China. 2010. Available online: http://www.gdofa.gov.cn accessed on 28 June 2011.
- De Innocentiis, S; Sola, L; Cataudella, S; Bentzen, P. Allozyme and microsatellite loci provide discordant estimates of population differentiation in the endangered dusky grouper (Epinephelus marginatus) within the Mediterranean Sea. Mol. Ecol 2001, 10, 2163–2175. [Google Scholar]
- Zhang, DC; Yu, LN; FYL. A study on genetic diversity of wild and cultured grass carp populations using RAPD. Freshwater Fish 2004, 4, 5–7. [Google Scholar]
- Liu, F; Xia, J; Bai, Z; Fu, J; Li, J; Yue, GH. High genetic diversity and substantial population differentiation in grass carp (Ctenopharyngodon idella) revealed by microsatellite analysis. Aquaculture 2009, 297, 51–56. [Google Scholar]
- Wade, MJ. Extinction and recolonization: Their effects on the genetic differentiation of local population. Evolution 1988, 42, 995–1005. [Google Scholar]
- Li, J; Wang, G; Bai, Z. Genetic variability in four wild and two farmed stocks of the Chinese freshwater pearl mussel (Hyriopsis cumingii) estimated by microsatellite DNA Markers. Aquaculture 2009, 287, 286–291. [Google Scholar]
- Sugama, K; Tridjoko, H; Budi, S; Cholik, F. Genetic variation and population structure in the humpback grouper, Cromileptes altivelis, throughout its range in Indonesian waters. Indo. Fish. Res. J 1999, 1, 32–38. [Google Scholar]
- Carlin, JL; Robertson, DR; Bowen, BW. Ancient divergences and recent connections in two tropical Atlantic reef fishes Epinephelus adscensionis and Rypticus saponaceous (Percoidei: Serranidae). Mar. Biol 2003, 143, 1057–1069. [Google Scholar]
- Hastings, A; Harrison, S. Metapopulation dynamics and genetics. Annu. Rev. Ecol. Syst 1994, 25, 167–188. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Locus | GenBank no. | Primer sequence (5′-3′) | Tann (°C) | Reference |
|---|---|---|---|---|
| D496 | DQ914905 | F: TTACTGGCAGCAATGGAC | 50 | [23] |
| R: GATGTATGACTACGAATGG | ||||
| Mbo061 | AY063512 | F: TGAAGAATGTCAGATATTTTGTGGTG | 53 | [24] |
| R: TCCCAAGAGTGTTGAAGTTATAGG | ||||
| Pm12 | AY688385 | F: AGAAAAAGCTCCACAACACAACAA | 55 | [25] |
| R: GAGCCCCAGTCCCAAATATTG | ||||
| CA-2 | AF539606 | F: GACTTGATTCAGCAAAATAAAGATG | 55 | [26] |
| R: AGAGACGGTGCCAGTAAATGAA | ||||
| CA-6 | AF539608 | F: GTGTTGCTGGGGTTACTAATGAAG | 50 | [26] |
| R: TTAGACACATTGTCACGATGGTCC | ||||
| GAA-1 | AF539612 | F: GGAGTGTTAAATATGCCCACCA | 60 | [26] |
| R: CAGAAATCGCTGAGGACAAGAG | ||||
| RH_GATA_003 | DQ223790 | F: GGGCAATTTGGTTCTTCACA | 57 | [27] |
| R: TGTCAATGCCACAGGATACA | ||||
| Ec_122 | GQ267997 | F: CATTCCTTAAAGTATTCTGTG | 55 | [21] |
| R: CCACAGCCAGTCTAGGTATTC | ||||
| Ec_154 | GQ429007 | F: AGCTGCTCAACAGGTTGTGTT | 56 | [21] |
| R: CAAGTTCCATATGTGCTCTGACA | ||||
| Ec_157 | GQ429008 | F: TGGAACAAGTTGGCATGGTA | 56 | [21] |
| R: CAAATACAACACCCTAGATTTT | ||||
| Ec_158 | GQ429009 | F: TGAGAGACAGTGGAGCACAAA | 56 | [21] |
| R: CGTGGTTACATTCTACCCCCTA |
| Locus | HZ n = 26 | SZ n = 15 | ZJ n = 34 | DYB n = 50 | HNI n = 40 | TRK n = 11 | SDK n = 25 | Mean | |
|---|---|---|---|---|---|---|---|---|---|
| D496 | A | 13 | 12 | 17 | 23 | 25 | 13 | 17 | 17.143 |
| Ar | 9.417 | 10.691 | 9.031 | 12.647 | 12.764 | 12.260 | 11.542 | 11.193 | |
| He | 0.900 | 0.926 | 0.831 | 0.945 | 0.945 | 0.931 | 0.934 | 0.916 | |
| ƒ | 0.063 | 0.062 | 0.053 | 0.052 | 0.045 | 0.047 | 0.038 | ||
| P | 0.185 | 0.917 | 0.270 | 0.418 | 0.450 | 0.091 | 0.061 | ||
| Mbo061 | A | 4 | 5 | 6 | 10 | 11 | 5 | 6 | 6.714 |
| Ar | 2.692 | 3.992 | 3.974 | 4.930 | 5.508 | 4.900 | 4.247 | 4.320 | |
| He | 0.246 | 0.405 | 0.343 | 0.459 | 0.518 | 0.584 | 0.667 | 0.460 | |
| ƒ | −0.100 | −0.095 | −0.097 | −0.087 | −0.081 | −0.087 | −0.148 | ||
| P | 1.000 | 1.000 | 1.000 | 0.897 | 0.948 | 1.000 | 0.000* | ||
| Pm12 | A | 6 | 8 | 8 | 8 | 8 | 7 | 9 | 7.714 |
| Ar | 4.634 | 7.285 | 5.469 | 6.528 | 6.513 | 6.810 | 6.674 | 6.273 | |
| He | 0.727 | 0.871 | 0.612 | 0.826 | 0.784 | 0.831 | 0.811 | 0.780 | |
| ƒ | 0.061 | 0.036 | 0.047 | 0.042 | −0.011 | 0.027 | 0.027 | ||
| P | 0.227 | 0.395 | 0.173 | 0.305 | 0.001 | 0.351 | 0.036 | ||
| CA-2 | A | 4 | 6 | 5 | 9 | 10 | 3 | 18 | 7.857 |
| Ar | 3.973 | 5.565 | 3.470 | 4.894 | 5.842 | 2.818 | 10.627 | 5.313 | |
| He | 0.732 | 0.735 | 0.485 | 0.607 | 0.672 | 0.177 | 0.886 | 0.613 | |
| ƒ | 0.021 | 0.043 | 0.044 | 0.020 | 0.034 | 0.025 | −0.009 | ||
| P | 0.522 | 0.922 | 0.135 | 0.657 | 0.553 | 1.000 | 0.001 | ||
| GAA-1 | A | 3 | 6 | 7 | 3 | 6 | 2 | 3 | 4.286 |
| Ar | 2.999 | 4.896 | 4.460 | 2.954 | 3.704 | 2.000 | 2.979 | 3.427 | |
| He | 0.657 | 0.598 | 0.612 | 0.534 | 0.606 | 0.247 | 0.549 | 0.543 | |
| ƒ | −0.008 | −0.006 | 0.006 | 0.019 | 0.046 | 0.014 | 0.016 | ||
| P | 0.420 | 0.149 | 0.101 | 0.900 | 0.175 | 1.000 | 0.842 | ||
| RH_GATA_003 | A | 7 | 5 | 8 | 11 | 9 | 5 | 7 | 7.429 |
| Ar | 5.188 | 4.526 | 4.803 | 5.908 | 6.277 | 5.000 | 5.462 | 5.309 | |
| He | 0.698 | 0.540 | 0.648 | 0.672 | 0.731 | 0.616 | 0.743 | 0.664 | |
| ƒ | −0.020 | −0.018 | 0.017 | −0.027 | −0.032 | −0.034 | 0.003 | ||
| P | 0.417 | 0.510 | 0.302 | 0.131 | 0.013 | 0.131 | 0.478 | ||
| Ec_122 | A | 10 | 11 | 11 | 15 | 17 | 11 | 16 | 13.000 |
| Ar | 7.451 | 9.297 | 7.924 | 9.507 | 9.707 | 11.528 | 10.475 | 9.413 | |
| He | 0.808 | 0.880 | 0.847 | 0.887 | 0.876 | 0.944 | 0.883 | 0.875 | |
| ƒ | −0.006 | −0.032 | −0.019 | −0.050 | −0.044 | −0.039 | −0.054 | ||
| P | 0.034 | 0.395 | 0.046 | 0.953 | 0.749 | 0.655 | 0.062 | ||
| Ec_154 | A | 9 | 7 | 10 | 20 | 18 | 7 | 21 | 13.143 |
| Ar | 6.921 | 6.118 | 6.021 | 9.872 | 9.771 | 7.000 | 12.842 | 8.364 | |
| He | 0.794 | 0.738 | 0.740 | 0.888 | 0.896 | 0.726 | 0.944 | 0.818 | |
| ƒ | −0.087 | −0.091 | −0.065 | −0.169 | −0.138 | −0.109 | −0.117 | ||
| P | 0.014 | 0.053 | 0.000* | 0.094 | 0.792 | 0.976 | 0.873 | ||
| Ec_158 | A | 4 | 9 | 7 | 13 | 12 | 5 | 17 | 9.571 |
| Ar | 3.307 | 7.695 | 5.367 | 7.265 | 7.003 | 4.991 | 10.944 | 6.653 | |
| He | 0.608 | 0.874 | 0.758 | 0.821 | 0.793 | 0.714 | 0.887 | 0.779 | |
| ƒ | 0.040 | 0.038 | 0.052 | 0.012 | 0.014 | 0.038 | 0.047 | ||
| P | 0.096 | 0.748 | 0.035 | 0.078 | 0.125 | 0.698 | 0.000* | ||
| Mean | A | 6.667 | 7.667 | 8.778 | 12.444 | 12.889 | 6.444 | 12.667 | |
| Ar | 5.176 | 6.674 | 5.613 | 7.167 | 7.454 | 6.367 | 8.421 | ||
| He | 0.686 | 0.730 | 0.653 | 0.738 | 0.758 | 0.641 | 0.812 |
| Pop | HZ | SZ | ZJ | DYB | HNI | TRK | SDK |
|---|---|---|---|---|---|---|---|
| HZ | 0 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 |
| SZ | 0.053 | 0 | 0.000 | 0.063 | 0.000 | 0.000 | 0.000 |
| ZJ | 0.044 | 0.039 | 0 | 0.000 | 0.000 | 0.000 | 0.000 |
| DYB | 0.043 | 0.010 | 0.032 | 0 | 0.279 | 0.000 | 0.000 |
| HNI | 0.038 | 0.030 | 0.039 | 0.002 | 0 | 0.000 | 0.000 |
| TRK | 0.129 | 0.070 | 0.112 | 0.031 | 0.049 | 0 | 0.000 |
| SDK | 0.047 | 0.041 | 0.073 | 0.032 | 0.020 | 0.075 | 0 |
| Source of variation | Sum of squares | Variance components | Percentage variation | P value |
|---|---|---|---|---|
| Among groups | 18.448 | 0.047 | 1.381 | 0.014 |
| Among populations with groups | 43.486 | 0.102 | 2.987 | 0.030 |
| Among individuals within populations | 623.593 | −0.027 | −0.801 | −0.008 |
| Within individuals | 657.000 | 3.284 | 96.433 | 0.036 |
| Total | 1342.527 | 3.406 | 100.000 |
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Wang, L.; Meng, Z.; Liu, X.; Zhang, Y.; Lin, H. Genetic Diversity and Differentiation of the Orange-Spotted Grouper (Epinephelus coioides) Between and Within Cultured Stocks and Wild Populations Inferred from Microsatellite DNA Analysis. Int. J. Mol. Sci. 2011, 12, 4378-4394. https://doi.org/10.3390/ijms12074378
Wang L, Meng Z, Liu X, Zhang Y, Lin H. Genetic Diversity and Differentiation of the Orange-Spotted Grouper (Epinephelus coioides) Between and Within Cultured Stocks and Wild Populations Inferred from Microsatellite DNA Analysis. International Journal of Molecular Sciences. 2011; 12(7):4378-4394. https://doi.org/10.3390/ijms12074378
Chicago/Turabian StyleWang, Le, Zining Meng, Xiaochun Liu, Yong Zhang, and Haoran Lin. 2011. "Genetic Diversity and Differentiation of the Orange-Spotted Grouper (Epinephelus coioides) Between and Within Cultured Stocks and Wild Populations Inferred from Microsatellite DNA Analysis" International Journal of Molecular Sciences 12, no. 7: 4378-4394. https://doi.org/10.3390/ijms12074378
APA StyleWang, L., Meng, Z., Liu, X., Zhang, Y., & Lin, H. (2011). Genetic Diversity and Differentiation of the Orange-Spotted Grouper (Epinephelus coioides) Between and Within Cultured Stocks and Wild Populations Inferred from Microsatellite DNA Analysis. International Journal of Molecular Sciences, 12(7), 4378-4394. https://doi.org/10.3390/ijms12074378