Variations of Antioxidant Properties and NO Scavenging Abilities during Fermentation of Tea

Abstract
:1. Introduction
2. Results
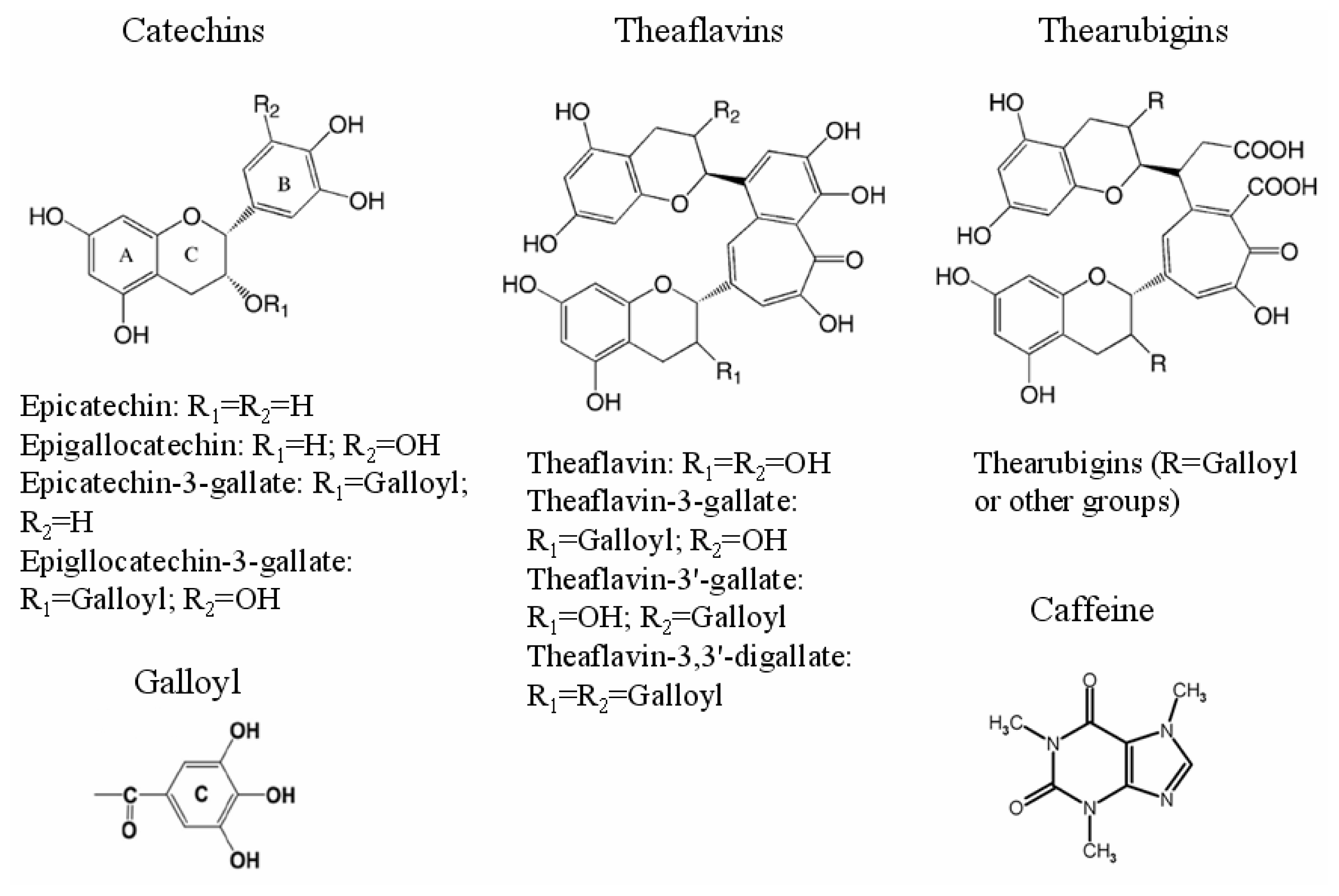
2.1. Contents of Several Polyphenol Ingredients in Tea
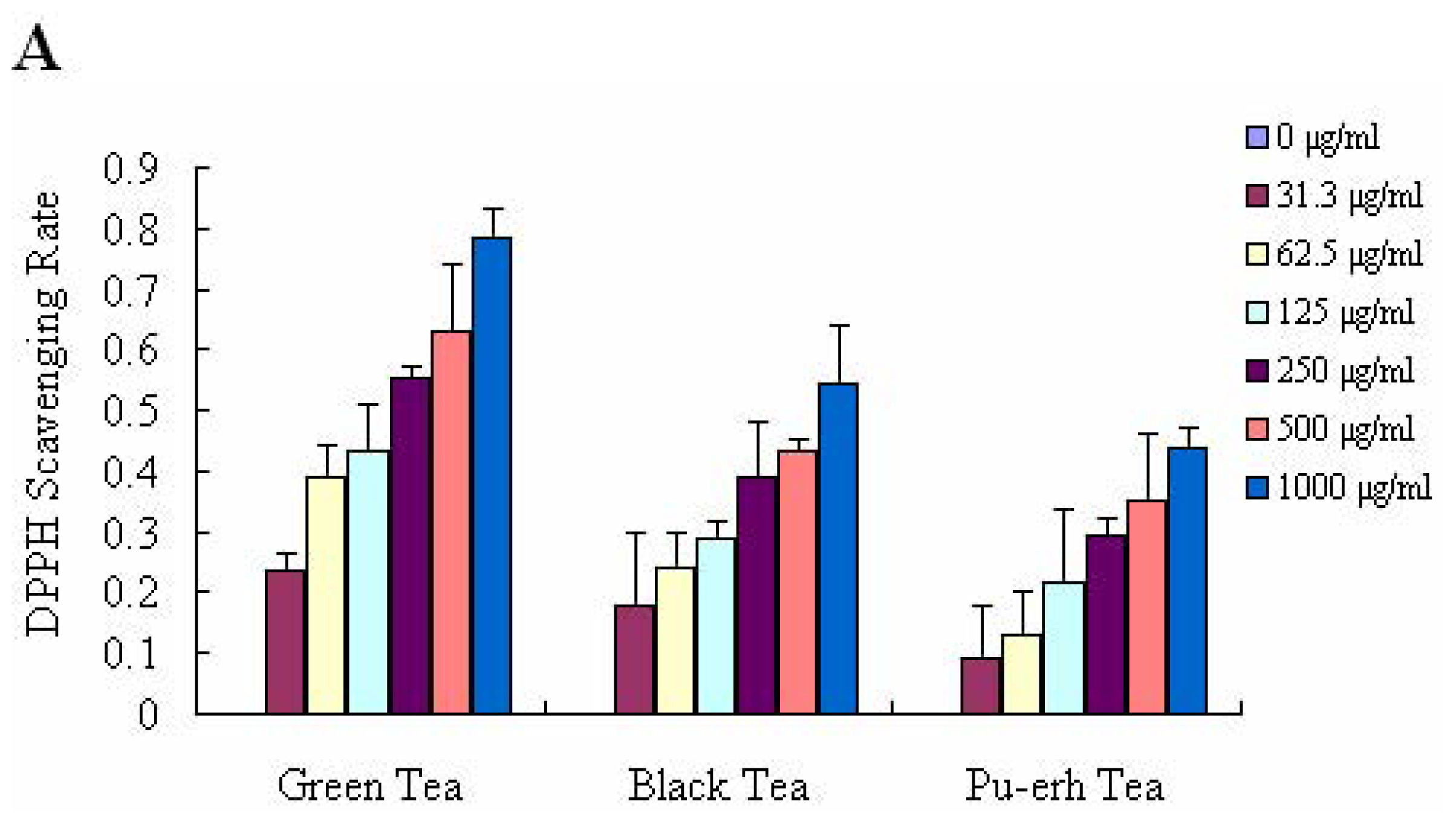
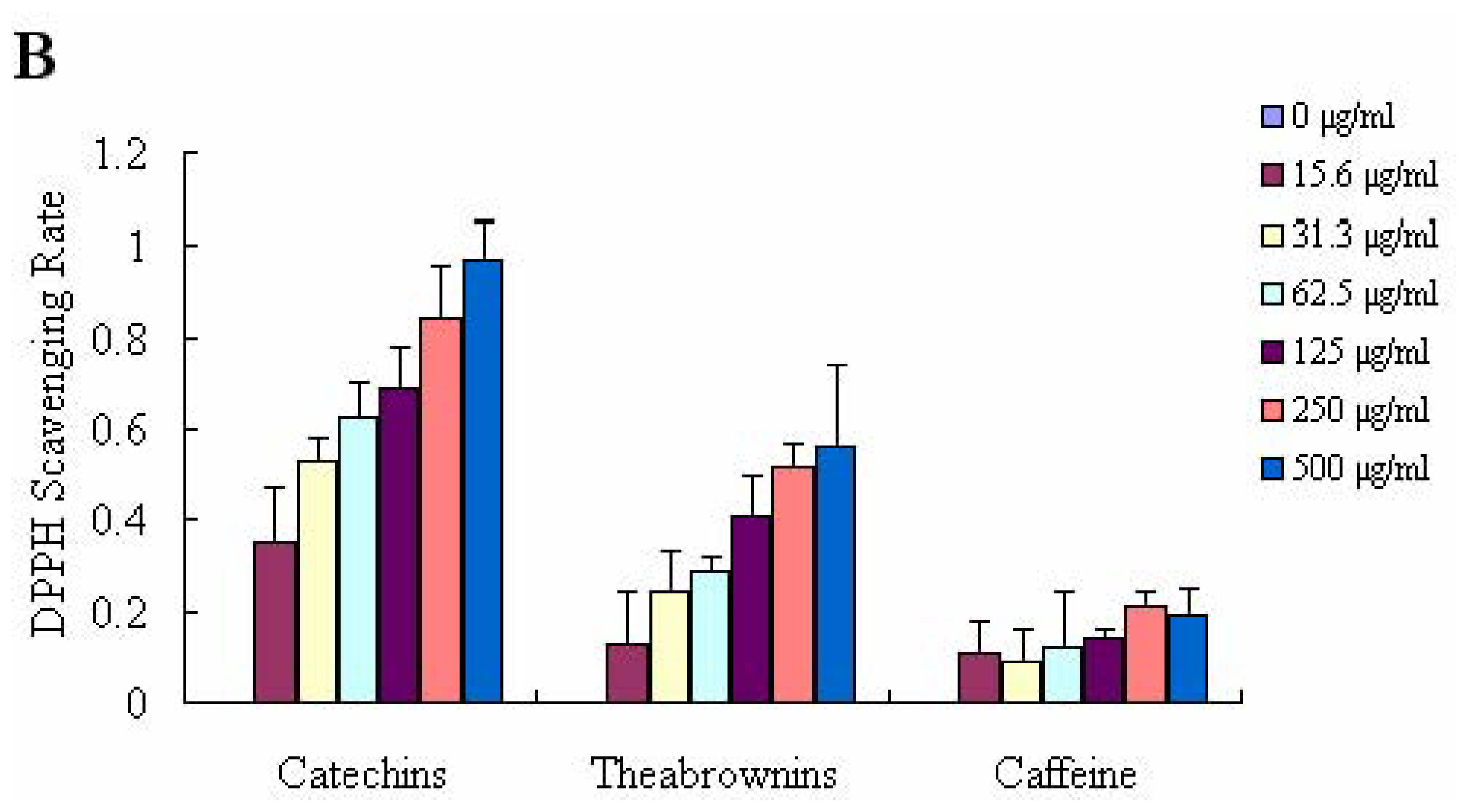
2.2. DPPH Radical Scavenging Activity and ABTS+ Decolourisation Assay
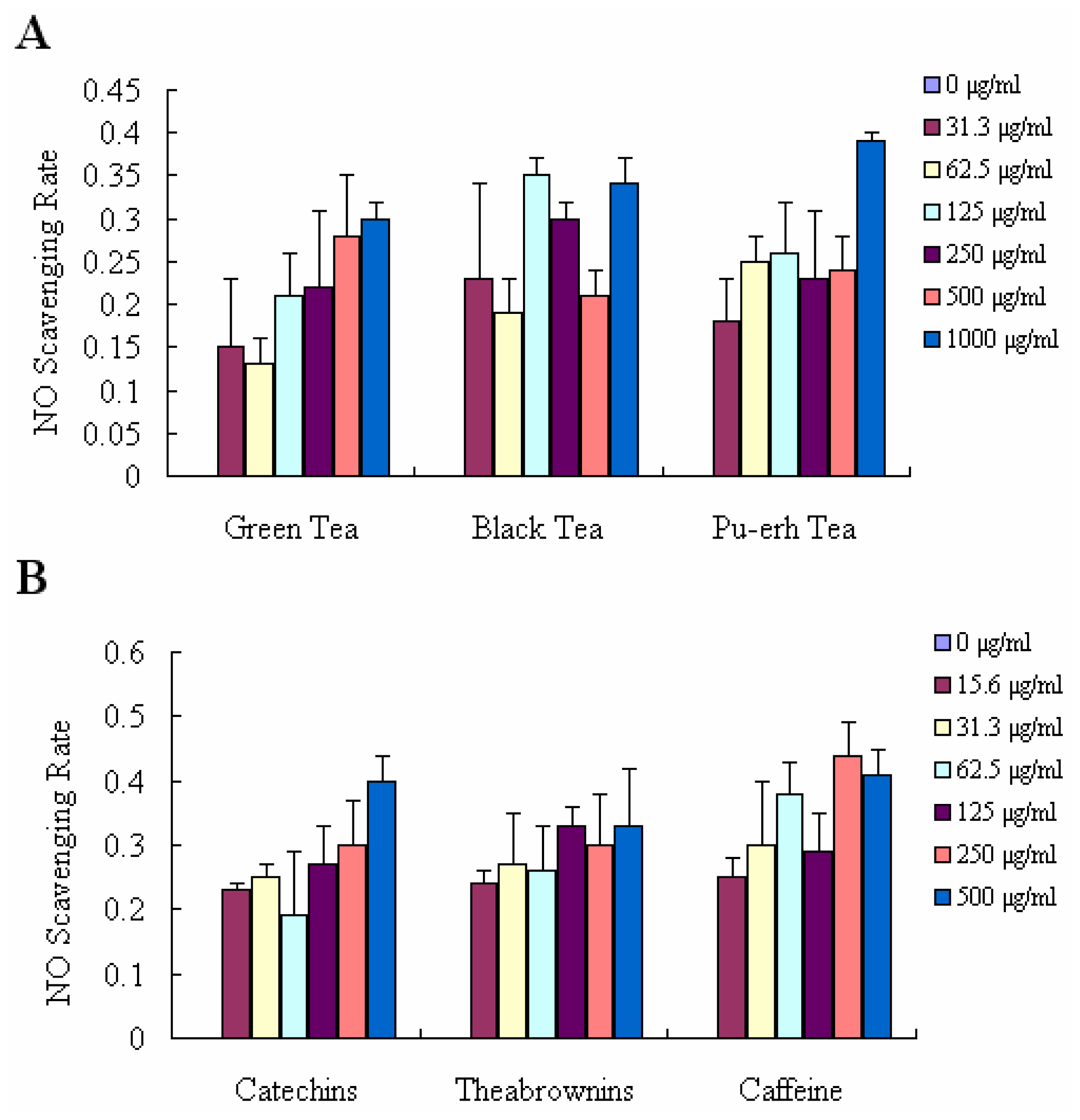
2.3. NO Scavenging Effect
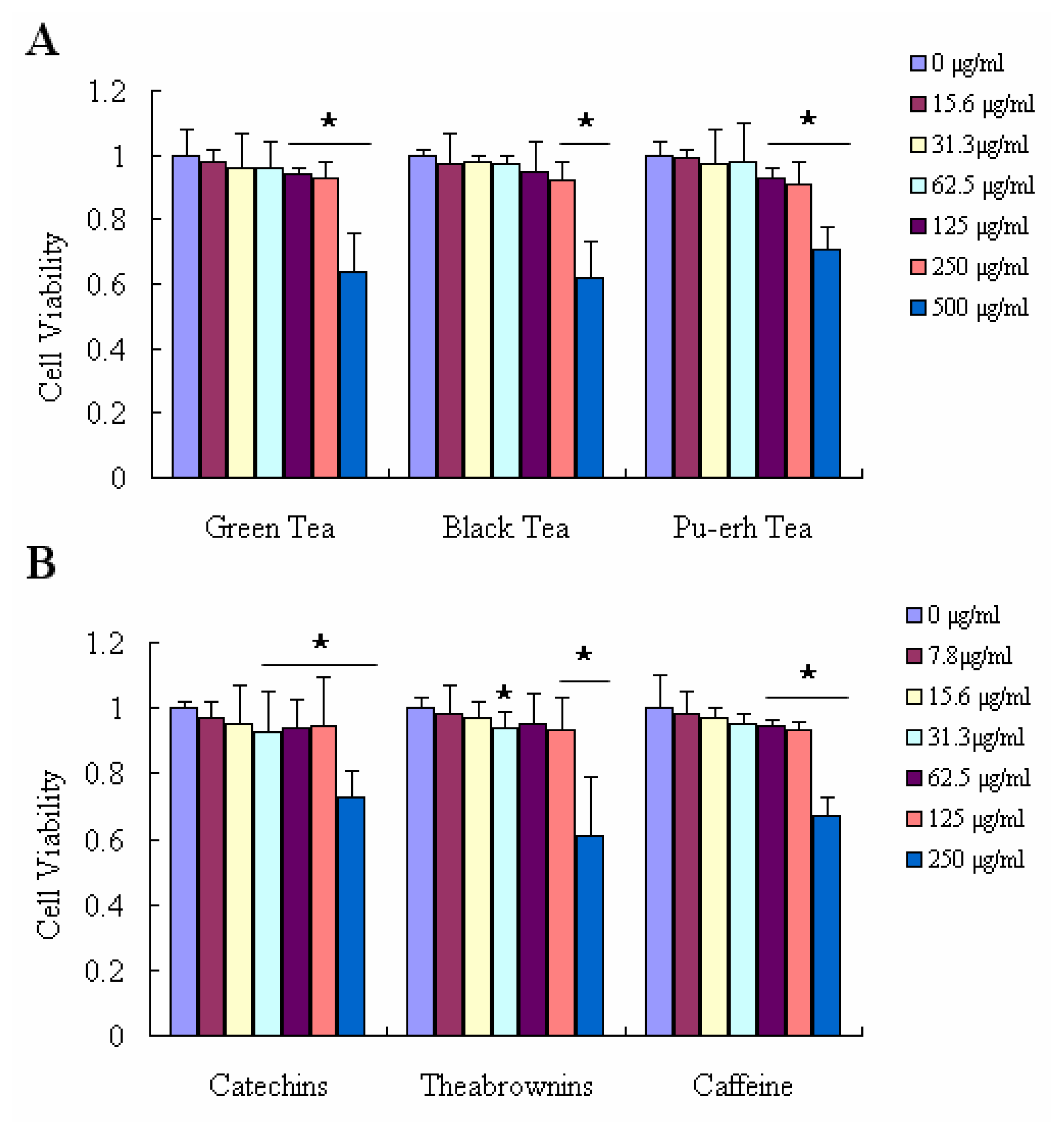
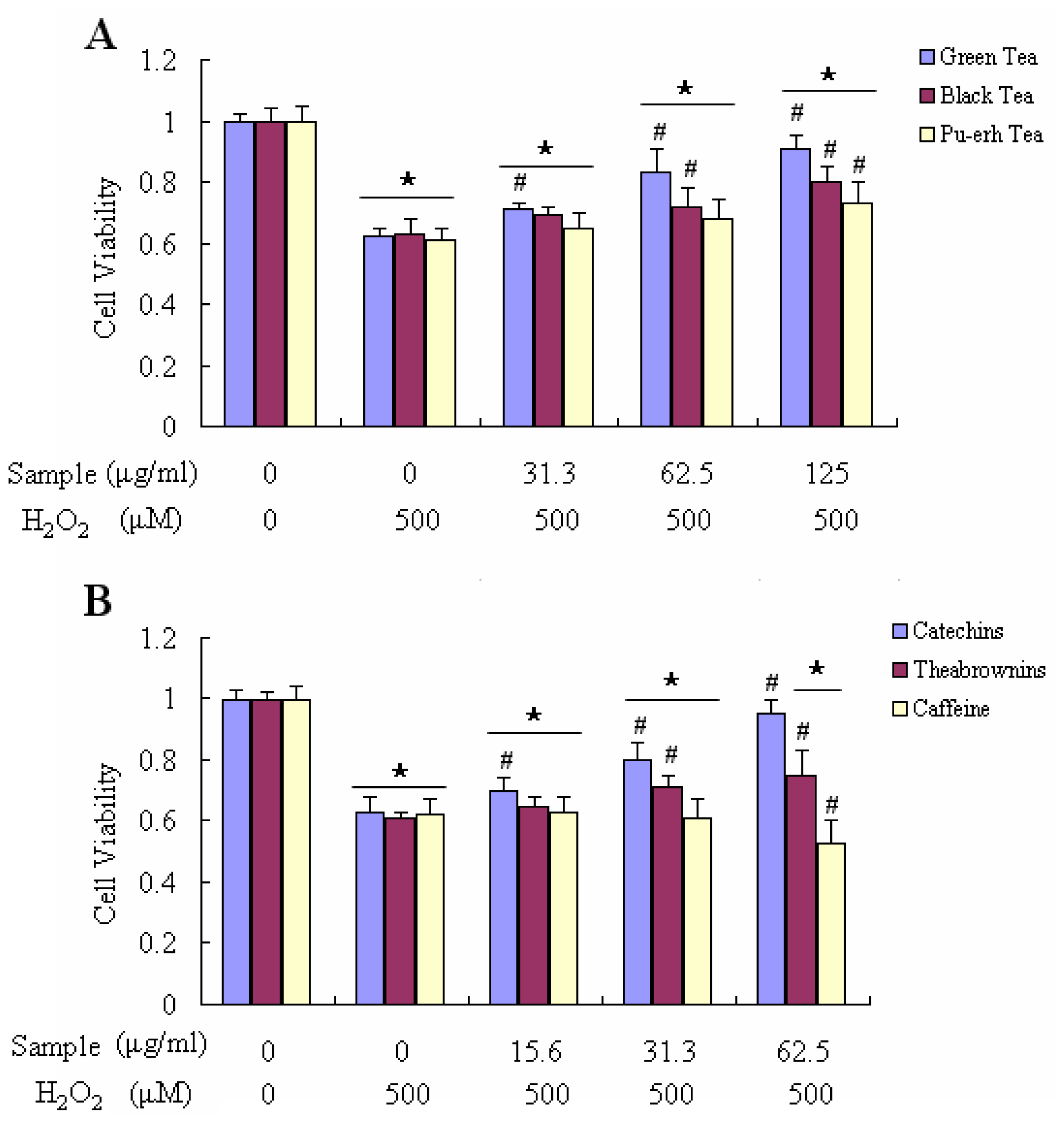
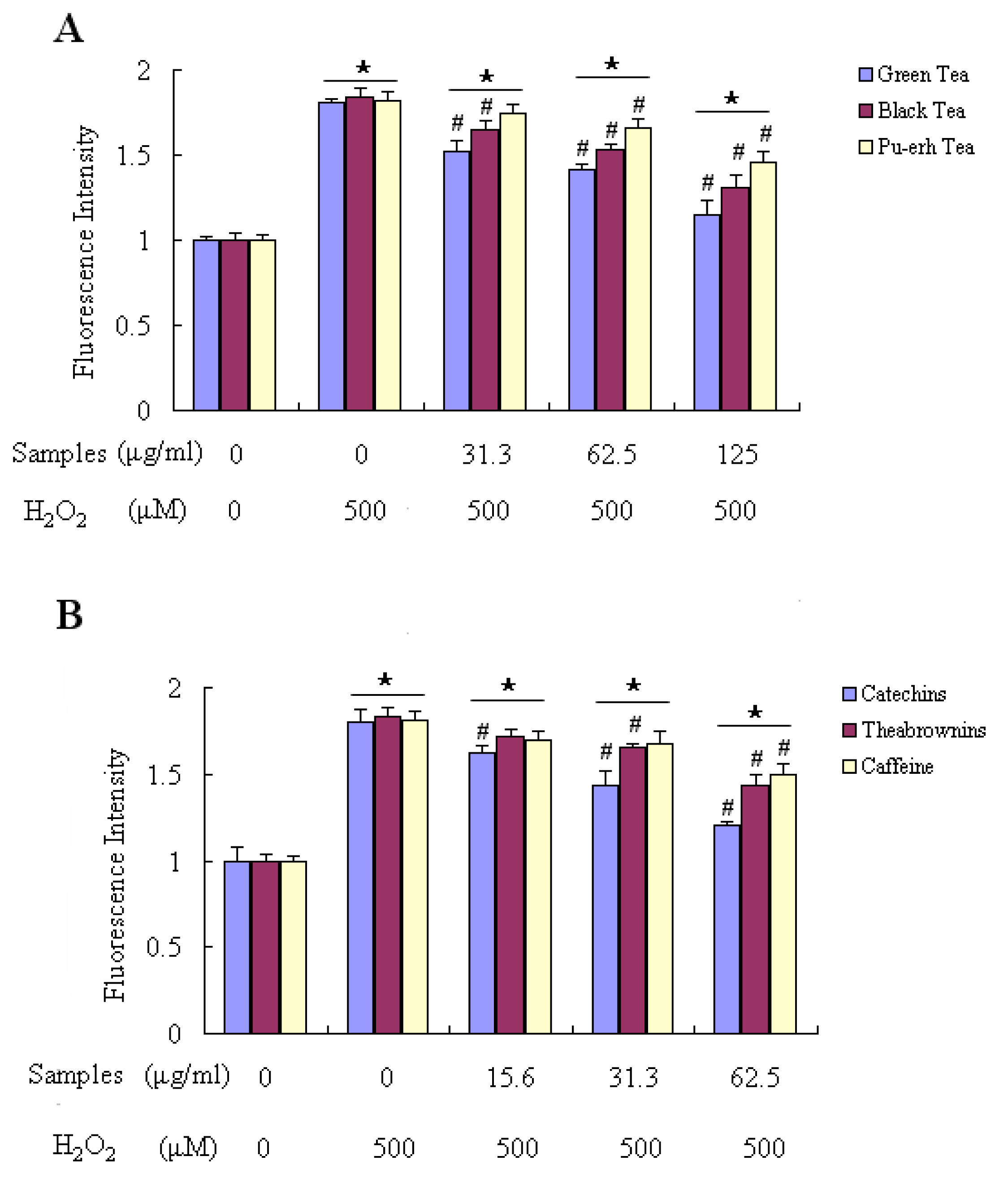
2.4. Effect on Ameliorating H2O2-Induced Loss of Cell Viability and Decreasing H2O2-Induced Accumulation of ROS in Raw 264.7 Cells
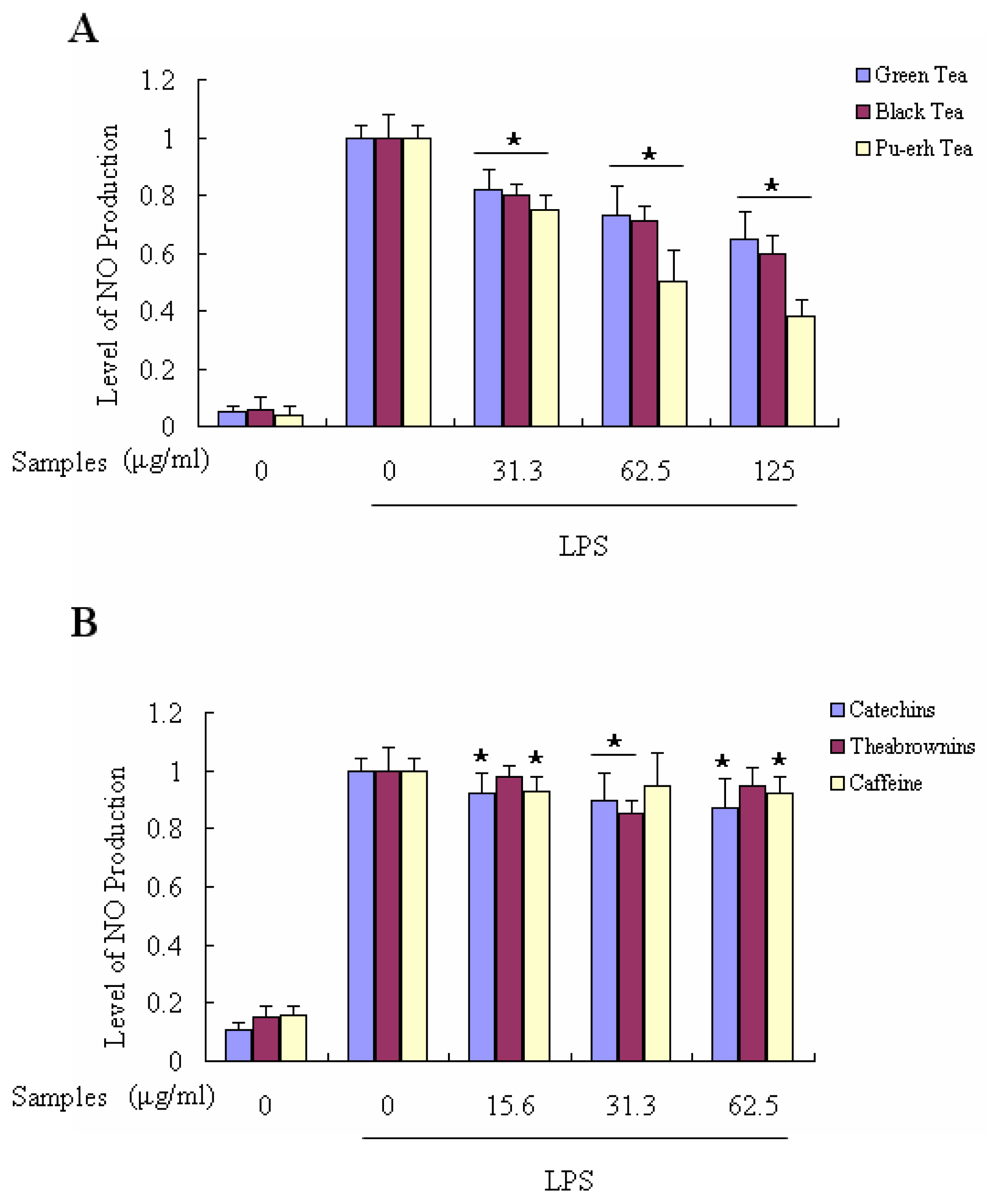
2.5. Suppressing Effects of Tea Extracts on LPS-Induced NO Production in Macrophages
3. Experimental Section
3.1. Materials
3.2. Preparation of Tea Extracts [22]
3.3. Determination of Polyphenol, Polysaccharides, and Caffeine Content in Concentrated Tea Extracts [22]
3.4. DPPH-Radical Scavenging Activity
3.5. ABTS+ Decolorization Assay
3.6. NO-Scavenging Activity
3.7. Cell Culture and Assessment of Cell Viability
3.8. Measurement of Intracellular Reactive Oxygen Species
3.9. Determination of NO Production
3.10. Statistical Analysis
4. Discussion
5. Conclusions
Acknowledgements
References
- Binns, CW; Lee, AH; Fraser, ML. Tea or coffee? A case study on evidence for dietary advice. Public Health Nutr 2008, 11, 1132–1141. [Google Scholar]
- McKay, DL; Blumberg, JB. The role of tea in human health: An update. J. Am. Coll. Nutr 2002, 21, 1–13. [Google Scholar]
- Melgarejo, E; Urdiales, JL; Sánchez-Jiménez, F; Medina, MA. Targeting polyamines and biogenic amines by green tea epigallocatechin-3-gallate. Amino Acids 2010, 38, 519–523. [Google Scholar]
- Jayabalan, R; Marimuthu, S; Swaminathan, K. Changes in content of organic acids and tea polyphenols during kombucha tea fermentation. Food Chem 2007, 102, 392–398. [Google Scholar]
- Zhao, JW; Chen, QS; Huang, XY. Qualitative identification of tea categories by near infrared spectroscopy and support vector machine. J. Pharm. Biomed. Anal 2006, 41, 1198–1204. [Google Scholar]
- Grove, KA; Lambert, JD. Laboratory, epidemiological, and human intervention studies show that tea (Camellia sinensis) may be useful in the prevention of obesity. J. Nutr 2010, 140, 446–453. [Google Scholar]
- Moore, RJ; Jackson, KG; Minihane, AM. Green tea (Camellia sinensis) catechins and vascular function. Br. J. Nutr 2009, 102, 1790–1802. [Google Scholar]
- Rains, TM; Agarwal, S; Maki, KC. Antiobesity effects of green tea catechins: A mechanistic review. J. Nutr. Biochem 2011, 22, 1–7. [Google Scholar]
- Hodgson, JM; Croft, KD. Tea flavonoids and cardiovascular health. Mol. Aspects Med 2010, 31, 495–502. [Google Scholar]
- Yanagimoto, K; Ochi, H; Lee, KG; Shibamoto, T. Antioxidative activities of volatile extracts from green tea, oolong tea, and black tea. J. Agric. Food Chem 2003, 51, 7396–7401. [Google Scholar]
- Yao, LH; Liu, X; Jiang, YM; Caffin, N; D’Arcy, B; Singanusong, R; Datta, N; Xu, Y. Compositional analysis of teas from Australian supermarkets. Food Chem 2006, 94, 115–122. [Google Scholar]
- Lin, YL; Juan, IM; Chen, YL; Liang, YC; Lin, JK. Composition of polyphenols in fresh tea leaves and associations of their oxygen-radical-absorbing capacity with antiproliferative actions in fibroblast cells. J. Agric. Food Chem 1996, 44, 1387–1394. [Google Scholar]
- Yang, ZY; Tu, YY; Baldermann, S; Fang Dong, F; Xu, Y; Watanabe, N. Isolation and identification of components from the ethanolic extract of flowers of the tea (Camellia sinensis) plant and their contribution to the antioxidant capacity. LWT-Food Sci. Technol 2009, 42, 1439–1443. [Google Scholar]
- Gong, J; Peng, C; Chen, T; Gao, B; Zhou, H. Effects of theabrownin from pu-erh tea on the metabolism of serum lipids in rats: Mechanism of action. J. Food Sci 2010, 75, 182–189. [Google Scholar]
- Yadav, SK; Ahuja, PS. Towards generating caffeine-free tea by metabolic engineering. Plant Foods Hum. Nutr 2007, 62, 185–191. [Google Scholar]
- Lambert, JD; Yang, CS. Cancer chemopreventive activity and bioavailability of tea and tea polyphenols. Mutat Res 2003, 523–524, 201–208. [Google Scholar]
- Duh, PD; Yen, GC; Yen, WJ; Wang, BS; Chang, LW. Effects of pu-erh tea on oxidative damage and nitric oxide scavenging. J. Agric. Food Chem 2004, 52, 8169–8176. [Google Scholar]
- Cheng, Z; Yan, G; Li, Y; Chang, W. Determination of antioxidant activity of phenolic antioxidants in a Fenton-type reaction system by chemiluminescence assay. Anal. Bioanal. Chem 2003, 375, 376–380. [Google Scholar]
- Lin, YS; Tsai, YJ; Tsay, JS; Lin, JK. Factors affecting the levels of tea polyphenols and caffeine in tea leaves. J. Agric. Food Chem 2003, 51, 1864–1873. [Google Scholar]
- Lin, CC; Lu, MJ; Chen, SJ; Ho, SC. Heavy fermentation impacts NO-suppressing activity of tea in LPS-activated Raw 264.7 macrophages. Food Chem 2006, 98, 483–489. [Google Scholar]
- Yang, Z; Jie, G; Dong, F; Xua, Y; Watanabe, N; Tu, Y. Radical-scavenging abilities and antioxidant properties of theaflavins and their gallate esters in H2O2-mediated oxidative damage system in the HPF-1 cells. Toxicol. in Vitro 2008, 22, 1250–1256. [Google Scholar]
- Zhao, H; Zhang, M; Zhao, L; Ge, Y; Sheng, J; Shi, W. Changes of constituents and activity to apoptosis and cell cycle during fermentation of tea. Int. J. Mol. Sci 2011, 12, 1862–1875. [Google Scholar]
- Zhu, QY; Hackman, RM; Ensunsa, JL; Holt, RR; Keen, CL. Antioxidative activities of oolong tea. J. Agric. Food Chem 2002, 50, 6929–6934. [Google Scholar]
- Jayaprakasha, GK; Singh, RP; Sakariah, KK. Antioxidant activity of grape seed (Vitis Vinifera) extracts on peroxidation models in vitro. Food Chem 2001, 73, 285–290. [Google Scholar]
- Re, R; Pelegrini, N; Proteggente, A; Pannala, A; Yang, M; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med 1999, 26, 1231–1237. [Google Scholar]
- Kim, H; Lee, HS; Chang, KT; Ko, TH; Baek, KJ; Kwon, NS. Chloromethyl ketones block induction of nitric oxide synthase in murine macrophages by preventing activation of nuclear factor-jB. J. Immunol 1995, 154, 4741–4748. [Google Scholar]
- Green, LC; Wagner, DA; Glogowski, J; Skipper, PL; Wishnok, JS; Tannenbaum, SR. Analysis of nitrate, nitrite and [15N] nitrite in biological fluids. Anal. Biochem 1982, 126, 131–138. [Google Scholar]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar]
- Denizot, F; Lang, R. Rapid colorimetric assay for cell growth and survival: Modifications to the tetrazolium dye procedure giving improved sensitivity and reliability. J. Immunol. Methods 1986, 89, 271–277. [Google Scholar]
- Yamamoto, T; Hsu, S; Lewis, J; Wataha, J; Dickinson, D; Singh, B; Bollag, WB; Lockwood, P; Ueta, E; Osakil, T; Schuster, G. Green tea polyphenol causes differential oxidative environments in tumor versus normal epithelial cells. J. Pharm. Exp. Ther 2003, 307, 230–236. [Google Scholar]
- Larsen, CA; Dashwood, RH; Bisson, WH. Tea catechins as inhibitors of receptor tyrosine kinases: Mechanistic insights and human relevance. Pharm. Res 2010, 62, 457–464. [Google Scholar]
- Baruah, AM. Fermentation characteristics of some sssamica clones and process optimization of black tea manufacturing. J. Agric. Food Chem 2003, 51, 6578–6588. [Google Scholar]
- Pryor, WA. Oxy-radicals and related species: Their formation, lifetimes, and reactions. Annu. Rev. Physiol 1986, 48, 657–667. [Google Scholar]
- Mayer, B; Hemmens, B. Biosynthesis and action of nitric oxide in mammalian cell. Trends Biochem. Sci 1997, 22, 477–481. [Google Scholar]
- Beckman, JS; Koppenol, WH. Nitric oxide, superoxide, and peroxynitrite: The good, the bad, and ugly. Am. J. Physiol 1996, 271, 1424–1437. [Google Scholar]
- Cooke, JP; Dzau, VJ. Nitric oxide synthase: Role in the genesis of vascular disease. Annu. Rev. Med 1997, 48, 489–509. [Google Scholar]
- Yermilov, V; Rubio, J; Becchi, M; Friesen, MD; Pignatelli, B; Ohshima, H. Formation of 8-nitroguanine by the reaction of guanine with peroxynitrite in vitro. Carcinogenesis 1995, 16, 2045–2050. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample (%w/w) | Polyphenols | Polysaccharides | Caffeine | Theabrownins |
|---|---|---|---|---|
| Green Tea | 56.23 ± 5.17b | 1.01 ± 0.11b | 8.62 ± 0.14b | – |
| Black Tea | 42.40 ± 3.35c | 3.42 ± 0.05c | 8.92 ± 0.19b | – |
| Pu-erh Tea | 33.13 ± 3.18d | 4.81 ± 0.13d | 9.31 ± 0.09b | 7.32–10.50 |
| Sample | Concentration (μg/mL)
| ||||||
|---|---|---|---|---|---|---|---|
| 0 | 31.3 | 62.5 | 125 | 250 | 500 | 1000 | |
| Green Tea | 0 | 30.21 ± 0.83b | 40.75 ± 0.23c | 50.47 ± 0.76d | 62.49 ± 0.58e | 78.22 ± 0.12f | 88.23 ± 0.73g |
| Black Tea | 0 | 22.38 ± 0.62b | 25.22 ± 0.21c | 30.37 ± 0.28d | 39.10 ± 0.62e | 45.21 ± 0.13f | 58.12 ± 0.91g |
| Pu-erh Tea | 0 | 11.35 ± 0.91b | 20.01 ± 0.12c | 25.46 ± 0.84d | 30.23 ± 0.62e | 31.2 ± 0.94d,e | 45.00 ± 0.61f |
| Sample | Concentration (μg/mL)
| ||||||
|---|---|---|---|---|---|---|---|
| 0 | 15.6 | 31.3 | 62.5 | 125 | 250 | 500 | |
| Catechins | 0 | 22.12 ± 0.42b | 35.40 ± 0.80c | 42.23 ± 0.71d | 63.54 ± 0.38e | 77.33 ± 0.13f | 90.23 ± 0.71g |
| Theabrownins | 0 | 13.32 ± 0.92b | 25.34 ± 0.23c | 29.13 ± 0.21d | 33.59 ± 0.22e | 39.98 ± 0.41f | 50.99 ± 0.62g |
| Caffeine | 0 | 9.09 ± 0.72b | 12.78 ± 0.72c | 7.29 ± 0.51d | 11.32 ± 0.32e | 15.45 ± 0.72f | 19.54 ± 0.97g |
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Xu, Y.; Zhao, H.; Zhang, M.; Li, C.-J.; Lin, X.-Z.; Sheng, J.; Shi, W. Variations of Antioxidant Properties and NO Scavenging Abilities during Fermentation of Tea. Int. J. Mol. Sci. 2011, 12, 4574-4590. https://doi.org/10.3390/ijms12074574
Xu Y, Zhao H, Zhang M, Li C-J, Lin X-Z, Sheng J, Shi W. Variations of Antioxidant Properties and NO Scavenging Abilities during Fermentation of Tea. International Journal of Molecular Sciences. 2011; 12(7):4574-4590. https://doi.org/10.3390/ijms12074574
Chicago/Turabian StyleXu, Yang, Hang Zhao, Min Zhang, Chun-Jie Li, Xue-Zhen Lin, Jun Sheng, and Wei Shi. 2011. "Variations of Antioxidant Properties and NO Scavenging Abilities during Fermentation of Tea" International Journal of Molecular Sciences 12, no. 7: 4574-4590. https://doi.org/10.3390/ijms12074574
APA StyleXu, Y., Zhao, H., Zhang, M., Li, C. -J., Lin, X. -Z., Sheng, J., & Shi, W. (2011). Variations of Antioxidant Properties and NO Scavenging Abilities during Fermentation of Tea. International Journal of Molecular Sciences, 12(7), 4574-4590. https://doi.org/10.3390/ijms12074574