Epigenetics: New Questions on the Response to Hypoxia

Abstract
:1. Introduction
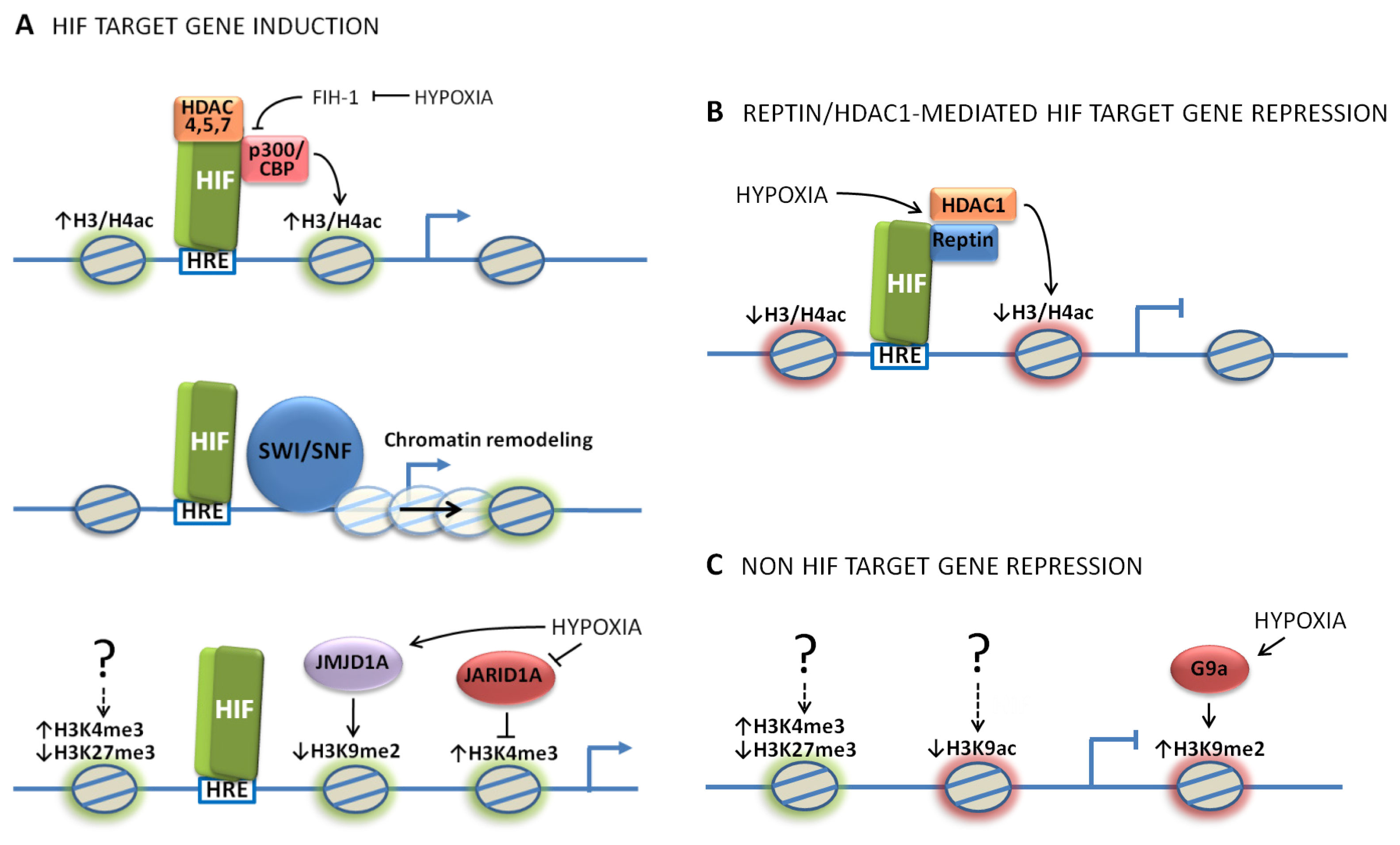
2. Role of HIF Co-Factors in the Response to Hypoxia
2.1. Role of Histone Acetyltransferases and Histone Deacetylases in the Response to Hypoxia
2.2. Role of Chromatin Remodeling in the Response to Hypoxia
3. Role of Jumonji-Domain Containing Histone Demethylases in the Response to Hypoxia
4. Gene-Specific Histone Modifications Induced under Hypoxia
5. Global Histone Modifications Induced under Hypoxia
6. Concluding Remarks
Acknowledgments
References
- Kaelin, WG, Jr; Ratcliffe, PJ. Oxygen sensing by metazoans: The central role of the HIF hydroxylase pathway. Mol Cell 2008, 30, 393–402. [Google Scholar]
- Semenza, GL. Regulation of oxygen homeostasis by hypoxia-inducible factor 1. Physiology (Bethesda) 2009, 24, 97–106. [Google Scholar]
- Majmundar, AJ; Wong, WJ; Simon, MC. Hypoxia-inducible factors and the response to hypoxic stress. Mol. Cell 2010, 40, 294–309. [Google Scholar]
- Stone, J; Itin, A; Alon, T; Pe’er, J; Gnessin, H; Chan-Ling, T; Keshet, E. Development of retinal vasculature is mediated by hypoxia-induced vascular endothelial growth factor (VEGF) expression by neuroglia. J. Neurosci 1995, 15, 4738–4747. [Google Scholar]
- Chen, EY; Fujinaga, M; Giaccia, AJ. Hypoxic microenvironment within an embryo induces apoptosis and is essential for proper morphological development. Teratology 1999, 60, 215–225. [Google Scholar]
- Krishnan, J; Ahuja, P; Bodenmann, S; Knapik, D; Perriard, E; Krek, W; Perriard, JC. Essential role of developmentally activated hypoxia-inducible factor 1alpha for cardiac morphogenesis and function. Circ. Res 2008, 103, 1139–1146. [Google Scholar]
- Dunwoodie, SL. The role of hypoxia in development of the Mammalian embryo. Dev. Cell 2009, 17, 755–773. [Google Scholar]
- Nangaku, M. Chronic hypoxia and tubulointerstitial injury: A final common pathway to end-stage renal failure. J. Am. Soc. Nephrol 2006, 17, 17–25. [Google Scholar]
- Shi, H. Hypoxia inducible factor 1 as a therapeutic target in ischemic stroke. Curr. Med. Chem 2009, 16, 4593–4600. [Google Scholar]
- Lu, X; Kang, Y. Hypoxia and hypoxia-inducible factors: Master regulators of metastasis. Clin. Cancer Res 2010, 16, 5928–5935. [Google Scholar]
- Semenza, GL. HIF-1: Upstream and downstream of cancer metabolism. Curr. Opin. Genet. Dev 2010, 20, 51–56. [Google Scholar]
- Maxwell, PH; Pugh, CW; Ratcliffe, PJ. Inducible operation of the erythropoietin 3′ enhancer in multiple cell lines: Evidence for a widespread oxygen-sensing mechanism. Proc. Natl. Acad. Sci. USA 1993, 90, 2423–2427. [Google Scholar]
- Wang, GL; Semenza, GL. General involvement of hypoxia-inducible factor 1 in transcriptional response to hypoxia. Proc. Natl. Acad. Sci. USA 1993, 90, 4304–4308. [Google Scholar]
- Semenza, GL. HIF-1, O(2), and the 3 PHDs: How animal cells signal hypoxia to the nucleus. Cell 2001, 107, 1–3. [Google Scholar]
- Bruick, RK. Oxygen sensing in the hypoxic response pathway: Regulation of the hypoxia-inducible transcription factor. Genes. Dev 2003, 17, 2614–2623. [Google Scholar]
- Bruick, RK; McKnight, SL. A conserved family of prolyl-4-hydroxylases that modify HIF. Science 2001, 294, 1337–1340. [Google Scholar]
- Epstein, AC; Gleadle, JM; McNeill, LA; Hewitson, KS; O'Rourke, J; Mole, DR; Mukherji, M; Metzen, E; Wilson, MI; Dhanda, A; et al. C. elegans EGL-9 and mammalian homologs define a family of dioxygenases that regulate HIF by prolyl hydroxylation. Cell 2001, 107, 43–54. [Google Scholar]
- Schofield, CJ; Ratcliffe, PJ. Oxygen sensing by HIF hydroxylases. Nat. Rev. Mol. Cell Biol 2004, 5, 343–354. [Google Scholar]
- Maxwell, PH; Wiesener, MS; Chang, GW; Clifford, SC; Vaux, EC; Cockman, ME; Wykoff, CC; Pugh, CW; Maher, ER; Ratcliffe, PJ. The tumour suppressor protein VHL targets hypoxia-inducible factors for oxygen-dependent proteolysis. Nature 1999, 399, 271–275. [Google Scholar]
- Ivan, M; Kondo, K; Yang, H; Kim, W; Valiando, J; Ohh, M; Salic, A; Asara, JM; Lane, WS; Kaelin, WG, Jr. HIFalpha targeted for VHL-mediated destruction by proline hydroxylation: Implications for O2 sensing. Science 2001, 292, 464–468. [Google Scholar]
- Jaakkola, P; Mole, DR; Tian, YM; Wilson, MI; Gielbert, J; Gaskell, SJ; Kriegsheim, A; Hebestreit, HF; Mukherji, M; Schofield, CJ; Maxwell, PH; Pugh, CW; Ratcliffe, PJ. Targeting of HIF-alpha to the von Hippel-Lindau ubiquitylation complex by O2-regulated prolyl hydroxylation. Science 2001, 292, 468–472. [Google Scholar]
- Johnson, AB; Denko, N; Barton, MC. Hypoxia induces a novel signature of chromatin modifications and global repression of transcription. Mutat. Res 2008, 640, 174–179. [Google Scholar]
- Kornberg, RD; Lorch, Y. Twenty-five years of the nucleosome, fundamental particle of the eukaryote chromosome. Cell 1999, 98, 285–294. [Google Scholar]
- Hayes, JJ; Hansen, JC. Nucleosomes and the chromatin fiber. Curr. Opin. Genet. Dev 2001, 11, 124–129. [Google Scholar]
- Horn, PJ; Peterson, CL. Molecular biology. Chromatin higher order folding--wrapping up transcription. Science 2002, 297, 1824–1827. [Google Scholar]
- Nemeth, A; Langst, G. Chromatin higher order structure: Opening up chromatin for transcription. Brief. Funct. Genomic Proteomic 2004, 2, 334–343. [Google Scholar]
- Tremethick, DJ. Higher-order structures of chromatin: The elusive 30 nm fiber. Cell 2007, 128, 651–654. [Google Scholar]
- Luger, K; Richmond, TJ. The histone tails of the nucleosome. Curr. Opin. Genet. Dev 1998, 8, 140–146. [Google Scholar]
- Jenuwein, T; Allis, CD. Translating the histone code. Science 2001, 293, 1074–1080. [Google Scholar]
- Peterson, CL; Laniel, MA. Histones and histone modifications. Curr. Biol 2004, 14, R546–R551. [Google Scholar]
- Lee, JS; Smith, E; Shilatifard, A. The language of histone crosstalk. Cell 2010, 142, 682–685. [Google Scholar]
- Eisen, JA; Sweder, KS; Hanawalt, PC. Evolution of the SNF2 family of proteins: Subfamilies with distinct sequences and functions. Nucleic Acids Res 1995, 23, 2715–2723. [Google Scholar]
- Muchardt, C; Yaniv, M. ATP-dependent chromatin remodelling: SWI/SNF and Co. are on the job. J. Mol. Biol 1999, 293, 187–198. [Google Scholar]
- Havas, K; Whitehouse, I; Owen-Hughes, T. ATP-dependent chromatin remodeling activities. Cell. Mol. Life Sci 2001, 58, 673–682. [Google Scholar]
- Johnson, AB; Barton, MC. Hypoxia-induced and stress-specific changes in chromatin structure and function. Mutat. Res 2007, 618, 149–162. [Google Scholar]
- Arany, Z; Huang, LE; Eckner, R; Bhattacharya, S; Jiang, C; Goldberg, MA; Bunn, HF; Livingston, DM. An essential role for p300/CBP in the cellular response to hypoxia. Proc. Natl. Acad. Sci. USA 1996, 93, 12969–12973. [Google Scholar]
- Ema, M; Hirota, K; Mimura, J; Abe, H; Yodoi, J; Sogawa, K; Poellinger, L; Fujii-Kuriyama, Y. Molecular mechanisms of transcription activation by HLF and HIF1alpha in response to hypoxia: Their stabilization and redox signal-induced interaction with CBP/p300. EMBO J 1999, 18, 1905–1914. [Google Scholar]
- Carrero, P; Okamoto, K; Coumailleau, P; O'Brien, S; Tanaka, H; Poellinger, L. Redoxregulated recruitment of the transcriptional coactivators CREB-binding protein and SRC-1 to hypoxia-inducible factor 1alpha. Mol. Cell Biol 2000, 20, 402–415. [Google Scholar]
- Ruas, JL; Poellinger, L; Pereira, T. Functional analysis of hypoxia-inducible factor-1 alphamediated transactivation. Identification of amino acid residues critical for transcriptional activation and/or interaction with CREB-binding protein. J. Biol. Chem 2002, 277, 38723–38730. [Google Scholar]
- Ruas, JL; Poellinger, L; Pereira, T. Role of CBP in regulating HIF-1-mediated activation of transcription. J. Cell Sci 2005, 118, 301–311. [Google Scholar]
- Kato, H; Tamamizu-Kato, S; Shibasaki, F. Histone deacetylase 7 associates with hypoxia-inducible factor 1alpha and increases transcriptional activity. J. Biol. Chem 2004, 279, 41966–41974. [Google Scholar]
- Seo, HW; Kim, EJ; Na, H; Lee, MO. Transcriptional activation of hypoxia-inducible factor-1alpha by HDAC4 and HDAC5 involves differential recruitment of p300 and FIH-1. FEBS Lett 2009, 583, 55–60. [Google Scholar]
- Wang, F; Zhang, R; Beischlag, TV; Muchardt, C; Yaniv, M; Hankinson, O. Roles of Brahma and Brahma/SWI2-related gene 1 in hypoxic induction of the erythropoietin gene. J. Biol. Chem 2004, 279, 46733–46741. [Google Scholar]
- Peserico, A; Simone, C. Physical and functional HAT/HDAC interplay regulates protein acetylation balance. J. Biomed. Biotechnol 2011, 2011, 371832. [Google Scholar]
- Fischle, W; Wang, Y; Allis, CD. Histone and chromatin cross-talk. Curr. Opin. Cell. Biol 2003, 15, 172–183. [Google Scholar]
- Shogren-Knaak, M; Ishii, H; Sun, JM; Pazin, MJ; Davie, JR; Peterson, CL. Histone H4-K16 acetylation controls chromatin structure and protein interactions. Science 2006, 311, 844–847. [Google Scholar]
- Kalkhoven, E. CBP and p300: HATs for different occasions. Biochem. Pharmacol 2004, 68, 1145–1155. [Google Scholar]
- Bannister, AJ; Kouzarides, T. The CBP co-activator is a histone acetyltransferase. Nature 1996, 384, 641–643. [Google Scholar]
- Yuan, W; Condorelli, G; Caruso, M; Felsani, A; Giordano, A. Human p300 protein is a coactivator for the transcription factor MyoD. J. Biol. Chem 1996, 271, 9009–9013. [Google Scholar]
- Dames, SA; Martinez-Yamout, M; De Guzman, RN; Dyson, HJ; Wright, PE. Structural basis for Hif-1 alpha /CBP recognition in the cellular hypoxic response. Proc. Natl. Acad. Sci. USA 2002, 99, 5271–5276. [Google Scholar]
- Freedman, SJ; Sun, ZY; Poy, F; Kung, AL; Livingston, DM; Wagner, G; Eck, MJ. Structural basis for recruitment of CBP/p300 by hypoxia-inducible factor-1 alpha. Proc. Natl. Acad. Sci. USA 2002, 99, 5367–5372. [Google Scholar]
- Ruas, JL; Berchner-Pfannschmidt, U; Malik, S; Gradin, K; Fandrey, J; Roeder, RG; Pereira, T; Poellinger, L. Complex regulation of the transactivation function of hypoxia-inducible factor-1 alpha by direct interaction with two distinct domains of the CREB-binding protein/p300. J. Biol. Chem 2010, 285, 2601–2609. [Google Scholar]
- Lando, D; Peet, DJ; Whelan, DA; Gorman, JJ; Whitelaw, ML. Asparagine hydroxylation of the HIF transactivation domain a hypoxic switch. Science 2002, 295, 858–861. [Google Scholar]
- Mahon, PC; Hirota, K; Semenza, GL. FIH-1: A novel protein that interacts with HIF-1alpha and VHL to mediate repression of HIF-1 transcriptional activity. Genes Dev 2001, 15, 2675–2686. [Google Scholar]
- Webb, JD; Coleman, ML; Pugh, CW. Hypoxia, hypoxia-inducible factors (HIF), HIF hydroxylases and oxygen sensing. Cell. Mol. Life Sci 2009, 66, 3539–3554. [Google Scholar]
- Pereira, T; Zheng, X; Ruas, JL; Tanimoto, K; Poellinger, L. Identification of residues critical for regulation of protein stability and the transactivation function of the hypoxia-inducible factor-1alpha by the von Hippel-Lindau tumor suppressor gene product. J. Biol. Chem 2003, 278, 6816–6823. [Google Scholar]
- Lisy, K; Peet, DJ. Turn me on: Regulating HIF transcriptional activity. Cell Death Differ 2008, 15, 642–649. [Google Scholar]
- Lim, JH; Lee, YM; Chun, YS; Chen, J; Kim, JE; Park, JW. Sirtuin 1 modulates cellular responses to hypoxia by deacetylating hypoxia-inducible factor 1alpha. Mol. Cell 2010, 38, 864–878. [Google Scholar]
- Dioum, EM; Chen, R; Alexander, MS; Zhang, Q; Hogg, RT; Gerard, RD; Garcia, JA. Regulation of hypoxia-inducible factor 2alpha signaling by the stress-responsive deacetylase sirtuin 1. Science 2009, 324, 1289–1293. [Google Scholar]
- Ebert, BL; Bunn, HF. Regulation of transcription by hypoxia requires a multiprotein complex that includes hypoxia-inducible factor 1, an adjacent transcription factor, and p300/CREB binding protein. Mol. Cell. Biol 1998, 18, 4089–4096. [Google Scholar]
- Jung, JE; Lee, HG; Cho, IH; Chung, DH; Yoon, SH; Yang, YM; Lee, JW; Choi, S; Park, JW; Ye, SK; Chung, MH. STAT3 is a potential modulator of HIF-1-mediated VEGF expression in human renal carcinoma cells. FASEB J 2005, 19, 1296–1298. [Google Scholar]
- Wang, F; Zhang, R; Wu, X; Hankinson, O. Roles of coactivators in hypoxic induction of the erythropoietin gene. PLoS One 2010, 5, e10002. [Google Scholar]
- Kasper, LH; Boussouar, F; Boyd, K; Xu, W; Biesen, M; Rehg, J; Baudino, TA; Cleveland, JL; Brindle, PK. Two transactivation mechanisms cooperate for the bulk of HIF-1-responsive gene expression. EMBO J 2005, 24, 3846–3858. [Google Scholar]
- Krieg, AJ; Rankin, EB; Chan, D; Razorenova, O; Fernandez, S; Giaccia, AJ. Regulation of the histone demethylase JMJD1A by hypoxia-inducible factor 1 alpha enhances hypoxic gene expression and tumor growth. Mol. Cell. Biol 2010, 30, 344–353. [Google Scholar]
- Zhou, X; Sun, H; Chen, H; Zavadil, J; Kluz, T; Arita, A; Costa, M. Hypoxia induces trimethylated H3 lysine 4 by inhibition of JARID1A demethylase. Cancer Res 2010, 70, 4214–4221. [Google Scholar]
- Islam, KN; Mendelson, CR. Permissive effects of oxygen on cyclic AMP and interleukin-1 stimulation of surfactant protein A gene expression are mediated by epigenetic mechanisms. Mol. Cell. Biol 2006, 26, 2901–2912. [Google Scholar]
- Chen, H; Yan, Y; Davidson, TL; Shinkai, Y; Costa, M. Hypoxic stress induces dimethylated histone H3 lysine 9 through histone methyltransferase G9a in mammalian cells. Cancer Res 2006, 66, 9009–9016. [Google Scholar]
- Tausendschon, M; Dehne, N; Brune, B. Hypoxia causes epigenetic gene regulation in macrophages by attenuating Jumonji histone demethylase activity. Cytokine 2011, 53, 256–262. [Google Scholar]
- Yang, XJ; Seto, E. Collaborative spirit of histone deacetylases in regulating chromatin structure and gene expression. Curr. Opin. Genet. Dev 2003, 13, 143–153. [Google Scholar]
- Ahringer, J. NuRD and SIN3 histone deacetylase complexes in development. Trends Genet 2000, 16, 351–356. [Google Scholar]
- Blander, G; Guarente, L. The Sir2 family of protein deacetylases. Annu. Rev. Biochem 2004, 73, 417–435. [Google Scholar]
- Yang, XJ; Gregoire, S. Class II histone deacetylases: From sequence to function, regulation, and clinical implication. Mol. Cell. Biol 2005, 25, 2873–2884. [Google Scholar]
- Vaquero, A; Sternglanz, R; Reinberg, D. NAD+-dependent deacetylation of H4 lysine 16 by class III HDACs. Oncogene 2007, 26, 5505–5520. [Google Scholar]
- Chen, S; Sang, N. Histone deacetylase inhibitors: The epigenetic therapeutics that repress hypoxia-inducible factors. J. Biomed. Biotechnol 2011, 2011, 197946. [Google Scholar]
- Lee, KJ; Lee, KY; Lee, YM. Downregulation of a tumor suppressor RECK by hypoxia through recruitment of HDAC1 and HIF-1alpha to reverse HRE site in the promoter. Biochim. Biophys. Acta 2010, 1803, 608–616. [Google Scholar]
- Lee, JS; Kim, Y; Kim, IS; Kim, B; Choi, HJ; Lee, JM; Shin, HJ; Kim, JH; Kim, JY; Seo, SB; et al. Negative regulation of hypoxic responses via induced Reptin methylation. Mol. Cell 2010, 39, 71–85. [Google Scholar]
- Fath, DM; Kong, X; Liang, D; Lin, Z; Chou, A; Jiang, Y; Fang, J; Caro, J; Sang, N. Histone deacetylase inhibitors repress the transactivation potential of hypoxia-inducible factors independently of direct acetylation of HIF-alpha. J. Biol. Chem 2006, 281, 13612–13619. [Google Scholar]
- Marks, PA; Richon, VM; Breslow, R; Rifkind, RA. Histone deacetylase inhibitors as new cancer drugs. Curr. Opin. Oncol 2001, 13, 477–483. [Google Scholar]
- Johnstone, RW; Licht, JD. Histone deacetylase inhibitors in cancer therapy: Is transcription the primary target? Cancer Cell 2003, 4, 13–18. [Google Scholar]
- Drummond, DC; Noble, CO; Kirpotin, DB; Guo, Z; Scott, GK; Benz, CC. Clinical development of histone deacetylase inhibitors as anticancer agents. Annu. Rev. Pharmacol. Toxicol 2005, 45, 495–528. [Google Scholar]
- Kenneth, NS; Mudie, S; van Uden, P; Rocha, S. SWI/SNF regulates the cellular response to hypoxia. J. Biol. Chem 2009, 284, 4123–4131. [Google Scholar]
- Tang, L; Nogales, E; Ciferri, C. Structure and function of SWI/SNF chromatin remodeling complexes and mechanistic implications for transcription. Prog. Biophys. Mol. Biol 2010, 102, 122–128. [Google Scholar]
- Dekanty, A; Romero, NM; Bertolin, AP; Thomas, MG; Leishman, CC; Perez-Perri, JI; Boccaccio, GL; Wappner, P. Drosophila genome-wide RNAi screen identifies multiple regulators of HIF-dependent transcription in hypoxia. PLoS Genet 2010, 6, e1000994. [Google Scholar]
- Klose, RJ; Kallin, EM; Zhang, Y. JmjC-domain-containing proteins and histone demethylation. Nat. Rev. Genet 2006, 7, 715–727. [Google Scholar]
- Shi, Y. Histone lysine demethylases: Emerging roles in development, physiology and disease. Nat. Rev. Genet 2007, 8, 829–833. [Google Scholar]
- Shi, Y; Whetstine, JR. Dynamic regulation of histone lysine methylation by demethylases. Mol. Cell 2007, 25, 1–14. [Google Scholar]
- Loenarz, C; Schofield, CJ. Expanding chemical biology of 2-oxoglutarate oxygenases. Nat. Chem. Biol 2008, 4, 152–156. [Google Scholar]
- Berger, SL. The complex language of chromatin regulation during transcription. Nature 2007, 447, 407–412. [Google Scholar]
- Xia, X; Lemieux, ME; Li, W; Carroll, JS; Brown, M; Liu, XS; Kung, AL. Integrative analysis of HIF binding and transactivation reveals its role in maintaining histone methylation homeostasis. Proc. Natl. Acad. Sci. USA 2009, 106, 4260–4265. [Google Scholar]
- Yang, J; Ledaki, I; Turley, H; Gatter, KC; Montero, JC; Li, JL; Harris, AL. Role of hypoxia-inducible factors in epigenetic regulation via histone demethylases. Ann. N. Y. Acad. Sci 2009, 1177, 185–197. [Google Scholar]
- Beyer, S; Kristensen, MM; Jensen, KS; Johansen, JV; Staller, P. The histone demethylases JMJD1A and JMJD2B are transcriptional targets of hypoxia-inducible factor HIF. J. Biol. Chem 2008, 283, 36542–36552. [Google Scholar]
- Pollard, PJ; Loenarz, C; Mole, DR; McDonough, MA; Gleadle, JM; Schofield, CJ; Ratcliffe, PJ. Regulation of Jumonji-domain-containing histone demethylases by hypoxiainducible factor (HIF)-1alpha. Biochem. J 2008, 416, 387–394. [Google Scholar]
- Wellmann, S; Bettkober, M; Zelmer, A; Seeger, K; Faigle, M; Eltzschig, HK; Buhrer, C. Hypoxia upregulates the histone demethylase JMJD1A via HIF-1. Biochem. Biophys. Res. Commun 2008, 372, 892–897. [Google Scholar]
- Costa, M; Davidson, TL; Chen, H; Ke, Q; Zhang, P; Yan, Y; Huang, C; Kluz, T. Nickel carcinogenesis: Epigenetics and hypoxia signaling. Mutat. Res 2005, 592, 79–88. [Google Scholar]
- Watson, JA; Watson, CJ; McCrohan, AM; Woodfine, K; Tosetto, M; McDaid, J; Gallagher, E; Betts, D; Baugh, J; O'Sullivan, J; Murrell, A; Watson, RW; McCann, A. Generation of an epigenetic signature by chronic hypoxia in prostate cells. Hum. Mol. Genet 2009, 18, 3594–3604. [Google Scholar]
- Watson, JA; Watson, CJ; McCann, A; Baugh, J. Epigenetics the epicenter of the hypoxic response. Epigenetics 2010, 5, 293–296. [Google Scholar]
- Xia, X; Kung, AL. Preferential binding of HIF-1 to transcriptionally active loci determines cell-type specific response to hypoxia. Genome Biol 2009, 10. [Google Scholar] [CrossRef]


{kind=link}
| Gene | Event | References |
|---|---|---|
| Hypoxia-induced genes: | ||
| VEGF A | ↑ H3ac ↑ H3K4me3 ↓ H3K27me3 | [22,61] |
| EPO | ↑ H3ac ↑ H4ac | [43,62] |
| EGR1 | ↑ H3ac ↑ H3K4me3 ↓ H3K9/27me2 ↓ H3K27me3 | [22] |
| ADM GDF15 | ↑ H3K9me2 | [64] |
| HMOX1 DAF | ↑ H3K4me3 | [65] |
| SP-A | ↓ H3ac ↑ H3K9me2 | [66] |
| AFP Albumin | ↓ H3ac ↑ H3K9/27me2 | [22] |
| ↑ H3K4me3 ↓ H3K27me3 | ||
| Mlh1 Dhfr | ↑ H3K9me2 | [67] |
| Ccl2 Ccr1 Ccr5 | ↑ H3K9me2 ↑ H3K9me3 | [68] |
| Histone | Residue | Modification | Event | References |
|---|---|---|---|---|
| H3 | K4 | me1 | ↑ | [22] |
| me2 | ↑ | [22,89] | ||
| me3 | ↑ | [22,65,89] | ||
| K9 | ac | ↓ | [22,26,66–67] | |
| me1 | ↓ | [67,94] | ||
| me2 | ↑ | [22,66–68,89,94] | ||
| me3 | ↑ | [67–68,94] | ||
| K14 | ac | ↑ | [22] | |
| K27 | me3 | ↑ | [22] | |
| K36 | me3 | ↑ | [68,89] | |
| K79 | me2 | ↑ | [22] | |
| H4 | R3 | me2 | ↑ | [22] |
| K5,8,12,16 | ac | ↓ | [22,94] |
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Perez-Perri, J.I.; Acevedo, J.M.; Wappner, P. Epigenetics: New Questions on the Response to Hypoxia. Int. J. Mol. Sci. 2011, 12, 4705-4721. https://doi.org/10.3390/ijms12074705
Perez-Perri JI, Acevedo JM, Wappner P. Epigenetics: New Questions on the Response to Hypoxia. International Journal of Molecular Sciences. 2011; 12(7):4705-4721. https://doi.org/10.3390/ijms12074705
Chicago/Turabian StylePerez-Perri, Joel I., Julieta M. Acevedo, and Pablo Wappner. 2011. "Epigenetics: New Questions on the Response to Hypoxia" International Journal of Molecular Sciences 12, no. 7: 4705-4721. https://doi.org/10.3390/ijms12074705
APA StylePerez-Perri, J. I., Acevedo, J. M., & Wappner, P. (2011). Epigenetics: New Questions on the Response to Hypoxia. International Journal of Molecular Sciences, 12(7), 4705-4721. https://doi.org/10.3390/ijms12074705