Ultrasmall Peptides Self-Assemble into Diverse Nanostructures: Morphological Evaluation and Potential Implications

Abstract
:1. Introduction
2. Results and Discussion
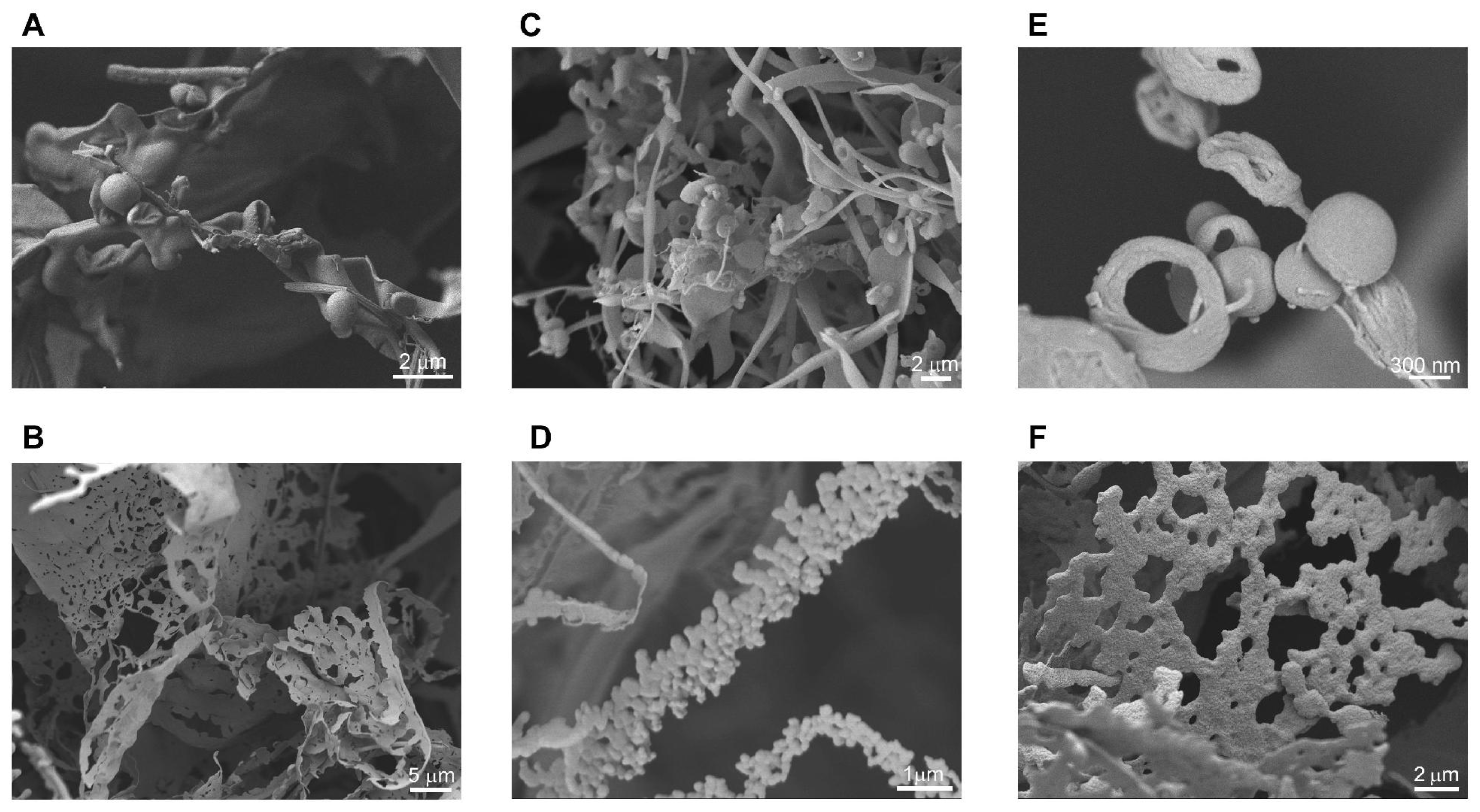
2.1. Nanostructures Formed at Low Concentrations of Aliphatic Peptides
2.2. Long, Helical Fibers Formed by Aliphatic Peptides
2.3. Short, Straight Fibers Formed by Modified Peptides with Aromatic Amino Acids
3. Experimental Section
4. Conclusions
Acknowledgments
References
- Whitesides, GM; Grzybowski, B. Self-assembly at all scales. Science 2002, 295, 2418–2421. [Google Scholar]
- Heckl, WM. Molecular Self-Assembly and the Origin of Life. In Astrobiology the Quest for the Conditions of Life, 1st ed; Horneck, G, Baumstark-Khan, C, Eds.; Springer: Berlin, Germany, 2002; pp. 360–371. [Google Scholar]
- Lehn, JM. Toward self-organization and complex matter. Science 2002, 295, 2400–2403. [Google Scholar]
- Ariga, K; Hill, JP; Lee, MV; Vinu, A; Charvet, R; Acharya, S. Challenges and breakthroughs in recent research on self-assembly. Sci Technol Adv Mater 2008, 9, 014109. [Google Scholar]
- Hauser, CAE; Zhang, S. Designer self-assembling peptide nanofiber biological materials. Chem Soc Rev 2010, 39, 2780–2790. [Google Scholar]
- Aggeli, A; Bell, M; Boden, N; Keen, JN; Knowles, PF; McLeish, TC; Pitkeathly, M; Radford, SE. Responsive gels formed by the spontaneous self-assembly of peptides into polymeric β-sheet tapes. Nature 1997, 386, 259–262. [Google Scholar]
- Hartgerink, JD; Beniash, E; Stupp, SI. Self-assembly and mineralization of peptide-amphiphile nanofibers. Science 2001, 294, 1684–1688. [Google Scholar]
- Vauthey, S; Santoso, S; Gong, H; Watson, N; Zhang, S. Molecular self-assembly of surfactant-like peptides to form nanotubes and nanovesicles. Proc Natl Acad Sci USA 2002, 99, 5355–5360. [Google Scholar]
- Ariga, K; Kikuchi, J; Naito, M; Koyama, E; Yamada, N. Modulated supramolecular assemblies composed of tripeptide derivatives: formation of micrometer-scale rods, nanometer-size needles, and regular patterns with molecular-level flatness from the same compound. Langmuir 2000, 16, 4929–4939. [Google Scholar]
- Place, ES; Evans, ND; Stevens, MM. Complexity in Biomaterials for Tissue Engineering. Nat Mater 2009, 8, 457–470. [Google Scholar]
- Adler-Abramovich, L; Reches, M; Sedman, VL; Allen, S; Tendler, SJB; Gazit, E. Thermal and chemical stability of diphenylalanine peptide nanotubes: Implications for nanotechnological applications. Langmuir 2006, 22, 1313–1320. [Google Scholar]
- Hauser, CAE; Deng, R; Mishra, A; Loo, Y; Khoe, U; Zhuang, F; Cheong, DW; Accardo, A; Sullivan, MB; Riekel, C; et al. Natural tri- to hexapeptides self-assemble in water to amyloid β-type fiber aggregates by unexpected α-helical intermediate structures. Proc Natl Acad Sci USA 2011, 108, 1361–1366. [Google Scholar]
- Mishra, A; Loo, Y; Deng, R; Chuah, YJ; Hee, HT; Ying, JY; Hauser, CAE. Ultrasmall natural peptides self-assemble to strong temperature-resistant helical fibers in scaffolds suitable for tissue engineering. Nano Today 2011, 6, 232–239. [Google Scholar]
- Marek, P; Abedini, A; Song, B; Kanungo, M; Johnson, ME; Gupta, R; Zaman, W; Wong, SS; Raleigh, DP. Aromatic interactions are not required for amyloid fibril formation by islet amyloid polypeptide but do influence the rate of fibril formation and fibril morphology. Biochemistry 2007, 46, 3255–3261. [Google Scholar]
- Oparin, AI. The Origin of Life; Macmillan: New York, NY, USA, 1938. [Google Scholar]
- Miller, SL. A production of amino acids under possible primitive earth conditions. Science 1953, 117, 528–529. [Google Scholar]
- Rode, BM. Peptides and the origin of life. Peptides 1999, 20, 773–786. [Google Scholar]
- Rode, BM; Fitz, D; Jakschitz, T. The first steps of chemical evolution towards the origin of life. Chem Biodivers 2007, 4, 2674–2702. [Google Scholar]
- Schwendinger, MG; Rode, BM. Investigations on the mechanism of the salt-induced peptide formation. Orig Life Evol Biosph 1992, 22, 349–359. [Google Scholar]
- Shepherd, NE; Hoang, HN; Abbenante, G; Fairlie, DP. Single turn peptide alpha helices with exceptional stability in water. J Am Chem Soc 2005, 127, 2974–2983. [Google Scholar]
- Banwell, EF; Abelardo, ES; Adams, DJ; Birchall, MA; Corrigan, A; Donald, AM; Kirkland, M; Serpell, LC; Butler, MF; Woolfson, DN. Rational design and application of responsive α-helical peptide hydrogels. Nat Mater 2009, 8, 596–600. [Google Scholar]
- Kisiday, J; Jin, M; Kurz, B; Hung, H; Semino, C; Zhang, S; Grodzinsky, AJ. Self-assembling peptide hydrogel fosters chondrocyte extracellular matrix production and cell division: implications for cartilage tissue repair. Proc Natl Acad Sci USA 2002, 99, 9996–10001. [Google Scholar]
- Kumada, Y; Hammond, NA; Zhang, S. Functionalized scaffolds of shorter self-assembling peptides containing MMP-2 cleavable motif promote fibroblast proliferation and significantly accelerate 3-D cell migration independent of scaffold stiffness. Soft Matter 2010, 6, 5073–5079. [Google Scholar]
- Tamamis, P; Adler-Abramovich, L; Reches, M; Marshall, K; Sikorski, P; Serpell, L; Gazit, E; Archontis, G. Self-assembly of phenylalanine oligopeptides: Insights from experiments and simulations. Biophys J 2009, 96, 5020–5029. [Google Scholar]
- Azriel, R; Gazit, E. Analysis of the minimal amyloid-forming fragment of the islet amyloid polypeptide. An experimental support for the key role of the phenylalanine residue in amyloid formation. J Biol Chem 2001, 276, 34156–34161. [Google Scholar]
- Gorbitz, CH. The Structure of nanotubes formed by diphenylalanine, the core recognition motif of Alzheimer’s β-amyloid polypeptide. Chem Commun 2006, 2332–2334. [Google Scholar]
- Gazit, EA. Possible role for π-Stacking in the self-assembly of amyloid fibrils. FASEB J 2002, 16, 77–83. [Google Scholar]
- Bemporad, F; Taddei, N; Stefani, M; Chiti, F. Assessing the role of aromatic residues in the amyloid aggregation of human muscle acylphosphatase. Protein Sci 2006, 15, 862–870. [Google Scholar]
- Reches, M; Gazit, E. Casting metal nanowires within discrete self-assembled peptide nanotubes. Science 2003, 300, 625–627. [Google Scholar]
- Yemini, M; Reches, M; Rishpon, J; Gazit, E. Novel electrochemical biosensing platform using self-assembled peptide nanotubes. Nano Lett 2004, 5, 183–186. [Google Scholar]
- Yemini, M; Reches, M; Gazit, E; Rishpon, J. Peptide nanotube-modified electrodes for enzyme—biosensor applications. Anal Chem 2005, 77, 5155–5159. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ultrasmall peptides containing only aliphatic amino acids | Ultrasmall peptides containing an aromatic head group | ||
|---|---|---|---|
| Hexamer | Trimer | Hexamer | Trimer |
| LIVAGD (LD6) | IVD (ID3) | LIVAGF (LF6) | IVF (IF3) |
| AIVAGD (AD6) | LIVAGW (LW6) | IVW (IW3) | |
| LIVAGY (LY6) | IVY (IY3) | ||
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Lakshmanan, A.; Hauser, C.A.E. Ultrasmall Peptides Self-Assemble into Diverse Nanostructures: Morphological Evaluation and Potential Implications. Int. J. Mol. Sci. 2011, 12, 5736-5746. https://doi.org/10.3390/ijms12095736
Lakshmanan A, Hauser CAE. Ultrasmall Peptides Self-Assemble into Diverse Nanostructures: Morphological Evaluation and Potential Implications. International Journal of Molecular Sciences. 2011; 12(9):5736-5746. https://doi.org/10.3390/ijms12095736
Chicago/Turabian StyleLakshmanan, Anupama, and Charlotte A.E. Hauser. 2011. "Ultrasmall Peptides Self-Assemble into Diverse Nanostructures: Morphological Evaluation and Potential Implications" International Journal of Molecular Sciences 12, no. 9: 5736-5746. https://doi.org/10.3390/ijms12095736
APA StyleLakshmanan, A., & Hauser, C. A. E. (2011). Ultrasmall Peptides Self-Assemble into Diverse Nanostructures: Morphological Evaluation and Potential Implications. International Journal of Molecular Sciences, 12(9), 5736-5746. https://doi.org/10.3390/ijms12095736