Hyaluronan Synthase and Hyaluronidase Expression in Serous Ovarian Carcinoma is Related to Anatomic Site and Chemotherapy Exposure

Abstract
:1. Introduction
2. Results
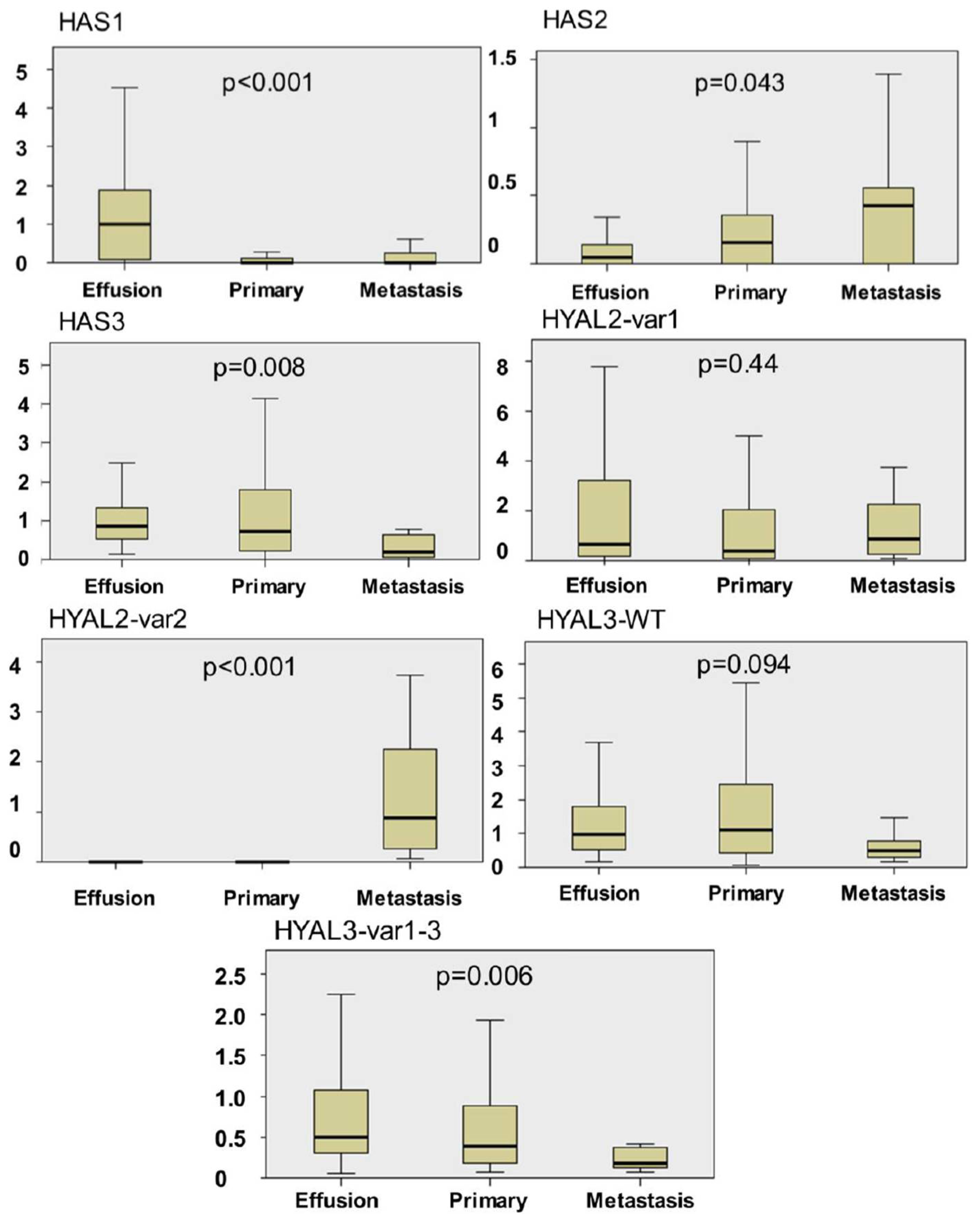
2.1. HAS and HYAL Expression is Significantly Different at Various Anatomic Sites in OC
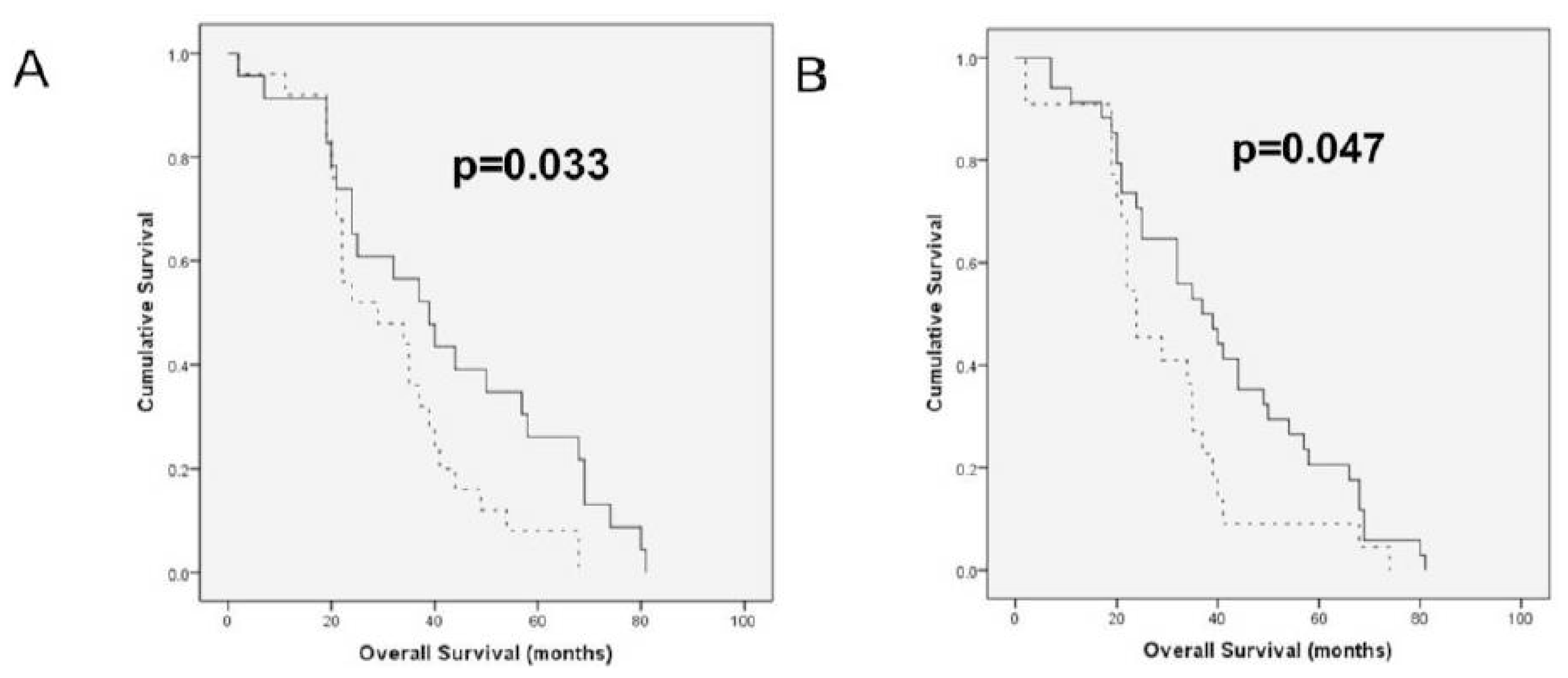
2.2. HAS and HYAL Expression Is Significantly Related to Clinicopathologic Parameters in OC Effusions
3. Discussion
4. Materials and Methods
4.1. Patients and Material
4.2. RT-PCR Analysis
4.3. Quantitative PCR (qPCR)
4.4. Statistical Analysis
5. Conclusion
Supplementary Materials
ijms-13-12925-s001.pdfAcknowledgements
References
- Agarwal, R.; Kaye, S.B. Ovarian cancer: Strategies for overcoming resistance to chemotherapy. Nat. Rev. Cancer 2003, 3, 502–516. [Google Scholar]
- Hennessy, B.T.; Coleman, R.L.; Markman, M. Ovarian cancer. Lancet 2009, 374, 1371–1382. [Google Scholar]
- Sironen, R.K.; Tammi, M.; Tammi, R.; Auvinen, P.K.; Anttila, M.; Kosma, V.-M. Hyaluronan in human malignancies. Exp. Cell Res 2011, 317, 383–391. [Google Scholar]
- Takahashi, E.; Nagano, O.; Ishimoto, T.; Yae, T.; Suzuki, Y.; Shinoda, T.; Nakamura, S.; Niwa, S.; Ikeda, S.; Koga, H.; Tanihara, H.; Saya, H. Tumor necrosis factor-alpha regulates transforming growth factor-beta-dependent epithelial-mesenchymal transition by promoting hyaluronan-CD44-moesin interaction. J. Biol. Chem 2010, 285, 4060–4073. [Google Scholar]
- Ween, M.P.; Oehler, M.K.; Ricciardelli, C. Role of Versican, Hyaluronan and CD44 in Ovarian Cancer Metastasis. Int. J. Mol. Sci. 2011, 12, 1009–1029. [Google Scholar]
- Tammi, R.H.; Passi, A.G.; Rilla, K.; Karousou, E.; Vigetti, D.; Makkonen, K.; Tammi, M.I. Transcriptional and post-translational regulation of hyaluronan synthesis. FEBS J 2011, 278, 1419–1428. [Google Scholar]
- Csoka, A.B.; Frost, G.I.; Stern, R. The six hyaluronidase-like genes in the human and mouse genomes. Matrix Biol 2001, 20, 499–508. [Google Scholar]
- Nykopp, T.K.; Rilla, K.; Sironen, R.; Tammi, M.I.; Tammi, R.H.; Hämäläinen, K.; Heikkinen, A.M.; Komulainen, M.; Kosma, V.M.; Anttila, M. Expression of hyaluronan synthases (HAS1-3) and hyaluronidases (HYAL1-2) in serous ovarian carcinomas: Inverse correlation between HYAL1 and hyaluronan content. BMC Cancer 2009, 9, 143. [Google Scholar]
- Tamakoshi, K.; Kikkawa, F.; Maeda, O.; Suganuma, N.; Yamagata, S.; Yamagata, T.; Tomoda, Y. Hyaluronidase activity in gynaecological cancer tissues with different metastatic forms. Br. J. Cancer 1997, 75, 1807–1811. [Google Scholar]
- Hiltunen, E.L.; Anttila, M.; Kultti, A.; Ropponen, K.; Penttinen, J.; Yliskoski, M.; Kuronen, A.T.; Juhola, M.; Tammi, R.; Tammi, M.; Kosma, V.M. Elevated hyaluronan concentration without hyaluronidase activation in malignant epithelial ovarian tumors. Cancer Res 2002, 62, 6410–6413. [Google Scholar]
- Yabushita, H.; Noguchi, M.; Kishida, T.; Fusano, K.; Noguchi, Y.; Itano, N.; Kimata, K.; Noguchi, M. Hyaluronan synthase expression in ovarian cancer. Oncol. Rep 2004, 12, 739–743. [Google Scholar]
- Davidson, B. Biological characteristics of cancers involving the serosal cavities. Crit. Rev. Oncog 2007, 13, 189–227. [Google Scholar]
- Köbel, M.; Kalloger, S.E.; Boyd, N.; McKinney, S.; Mehl, E.; Palmer, C.; Leung, S.; Bowen, N.J.; Ionescu, D.N.; Rajput, A.; et al. Ovarian carcinoma subtypes are different diseases: Implications for biomarker studies. PLoS Med 2008, 5, e232. [Google Scholar]
- Prat, J. Ovarian carcinomas: Five distinct diseases with different origins, genetic alterations, and clinicopathological features. Virchows Arch 2012, 460, 237–249. [Google Scholar]
- Lokeshwar, V.B.; Schroeder, G.L.; Carey, R.I.; Soloway, M.S.; Iida, N. Regulation of hyaluronidase activity by alternative mRNA splicing. J. Biol. Chem 2002, 277, 33654–33663. [Google Scholar]
- Slomiany, M.G.; Dai, L.; Tolliver, L.B.; Grass, G.D.; Zeng, Y.; Toole, B.P. Inhibition of Functional Hyaluronan-CD44 Interactions in CD133-positive Primary Human Ovarian Carcinoma Cells by Small Hyaluronan Oligosaccharides. Clin. Cancer Res 2009, 15, 7593–7601. [Google Scholar]
- Auzenne, E.; Ghosh, S.C.; Khodadadian, M.; Rivera, B.; Farquhar, D.; Price, R.E.; Ravoori, M.; Kundra, V.; Freedman, R.S.; Klostergaard, J. Hyaluronic acid-paclitaxel: Antitumor efficacy against CD44(+) human ovarian carcinoma xenografts. Neoplasia 2007, 9, 479–486. [Google Scholar]
- Banzato, A.; Bobisse, S.; Rondina, M.; Renier, D.; Bettella, F.; Esposito, G.; Quintieri, L.; Meléndez-Alafort, L.; Mazzi, U.; Zanovello, P.; Rosato, A. A paclitaxel-hyaluronan bioconjugate targeting ovarian cancer affords a potent in vivo therapeutic activity. Clin. Cancer Res 2008, 14, 3598–3606. [Google Scholar]
- De Stefano, I.; Battaglia, A.; Zannoni, G.F.; Prisco, M.G.; Fattorossi, A.; Travaglia, D.; Baroni, S.; Renier, D.; Scambia, G.; Ferlini, C.; Gallo, D. Hyaluronic acid-paclitaxel: Effects of intraperitoneal administration against CD44(+) human ovarian cancer xenografts. Cancer Chemother. Pharmacol. 2011, 68, 107–116. [Google Scholar]
- Anttila, M.A.; Tammi, R.H.; Tammi, M.I.; Syrjänen, K.J.; Saarikoski, S.V.; Kosma, V.M. High levels of stromal hyaluronan predict poor disease outcome in epithelial ovarian cancer. Cancer Res 2000, 60, 150–155. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Parameter | ||
|---|---|---|
| Age | Mean (range) | Number of patients 61 (38–79) |
| FIGO stage | III | 28 |
| IV | 28 | |
| Grade | Low | 4 |
| High | 46 | |
| NA a | 6 | |
| Residual disease | ≤1 cm | 20 |
| >1 cm | 28 | |
| NA b | 8 | |
| Chemotherapy status c | Pre-chemotherapy | 30 |
| Post-chemotherapy | 31 | |
| Chemotherapy response at diagnosis | Complete | 33 |
| Non-complete d | 16 | |
| Other e | 7 | |
| Clinical Parameter | Enzyme | ||||||
|---|---|---|---|---|---|---|---|
| HAS1 | HAS2 | HAS3 | HYAL2v1 | HYAL2v2 | HYAL3wt | HYAL3v1-3 | |
| Effusion site | 0.4 | 0.28 | 0.76 | 0.23 | 0.97 | 0.9 | 0.37 |
| Age | 0.71 | 0.23 | 0.68 | 0.84 | 0.18 | 0.4 | 0.89 |
| FIGO stage | 0.06 | 0.15 | 0.62 | 0.61 | 0.77 | 0.8 | 0.54 |
| RD volume | 0.78 | 0.54 | 0.31 | 0.15 | 0.87 | 0.19 | 0.54 |
| Chemoresponse | 0.99 | 0.59 | 0.3 | 0.94 | 0.23 | 0.35 | 0.06 |
| Previous chemotherapy | <0.001 | 0.23 | 0.5 | 0.016 | 0.79 | 0.024 | 0.34 |
| Previous platinum | <0.001 | 0.4 | 0.83 | 0.17 | 0.71 | 0.06 | 0.82 |
| Previous paclitaxel | 0.003 | 0.24 | 0.72 | 0.43 | 0.58 | 0.005 | 0.54 |
| mRNA | Primers | Product size (bp) |
|---|---|---|
| HAS1 | Sense: 5′-GAGGCCTGGTACAACCAGAA-35′ | 551 |
| Antisense: 55′-GCAGAGGGACGTAGTTAGCG-35′ | ||
| HAS2 | Sense: 55′-AAGGCTAACCTACCCTGGGA-35′ | 523 |
| Antisense: 55′-AATGCACTGAACACACCCAA-35′ | ||
| HAS3 | Sense: 55′-GACGACAGCCCTGCGTGT-35′ | 342 |
| Antisense: 55′-TTGAGGTCAGGGAAGGAGAT-35′ | ||
| HYAL1 wt + variant 1–4 | Sense: 55′-GGTCAGGAAATTTGGAGGAT-35′ | 1154 |
| Antisense: 55′-ACAGGGCTTGACTGCAGAGA-35′ | ||
| HYAL1 variant 5 | Sense: 55′-GTGGACAAAGAACACTCCCT-35′ | 1210 |
| Antisense: 55′-ACAGGGCTTGACTGCAGAGA-35′ | ||
| HYAL3 | Sense: 55′-ACACACCGGAGATCTGGGAG-35′ | 100 + 200 |
| Antisense: 55′-CTGGTCACATTGATCACATA-35′ | ||
| 28S | Sense: 55′-GTTCACCCACTAATAGGGAACGTGA-35′ | 212 |
| Antisense: 55′-GGATTCTGACTTAGAGGCGTTCAGT-35′ |
| Gene | Heating | Denaturation | Annealing | Extension | Cycles |
|---|---|---|---|---|---|
| HAS1 | 94 °C-5 min | 94 °C-15 s | 62 °C-30 s | 72 °C-20 s | 35 |
| HAS2 | 94 °C-5 min | 94 °C-15 s | 60 °C-30 s | 72 °C-20 s | 32 |
| HAS3 | 94 °C-5 min | 94 °C-15 s | 58 °C-30 s | 72 °C-20 s | 35 |
| HYAL1 | 94 °C-5 min | 94 °C-15 s | 59 °C-30 s | 72 °C-40 s | 40 |
| HYAL3 | 94 °C-5 min | 94 °C-15 s | 55.7 °C-30 s | 72 °C-20 s | 33 |
| 28S | 94 °C-5 min | 94 °C-15 s | 63 °C-20 s | 72 °C-10 s | 16 |
| mRNA | Primers | Product size (bp) |
|---|---|---|
| HYAL2 var1 | Sense: 55′-GGACTCCCACACAGTTCCTG-35′ | 157 |
| Antisense: 55′-GTGAAGATGGGTGGTGCTGT-35′ | ||
| HYAL2 var2 | Sense: 55′-GCGCGAGTTCCTGAGCTG-35′ | 154 |
| Antisense: 55′-CAGTGAAGATGGGTGGTGCT-35′ | ||
| 28S | Sense: 55′-GTTCACCCACTAATAGGGAACGTGA-35′ | 212 |
| Antisense: 55′-GGATTCTGACTTAGAGGCGTTCAGT-35′ | ||
| RPLPO | Sense: 55′-CCAACTACTTCCTTAAGATCATCCAACTA-35′ | 108 |
| Antisense: 55′-ACATGCGGATCTGCTGCA-35′ |
| Gene | Heating | Denaturation | Annealing | Extension | Cycles |
|---|---|---|---|---|---|
| HYAL2 | 95 °C-7 min | 95 °C-30 s | 63 °C-1 min | 72 °C-30 s | 40 |
| 28S | 95 °C-7 min | 95 °C-30 s | 60 °C-1 min | 72 °C-30 s | 40 |
| RPLPO | 95 °C-7 min | 95 °C-30 s | 60 °C-1 min | 72 °C-30 s | 40 |
© 2012 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Weiss, I.; Trope, C.G.; Reich, R.; Davidson, B. Hyaluronan Synthase and Hyaluronidase Expression in Serous Ovarian Carcinoma is Related to Anatomic Site and Chemotherapy Exposure. Int. J. Mol. Sci. 2012, 13, 12925-12938. https://doi.org/10.3390/ijms131012925
Weiss I, Trope CG, Reich R, Davidson B. Hyaluronan Synthase and Hyaluronidase Expression in Serous Ovarian Carcinoma is Related to Anatomic Site and Chemotherapy Exposure. International Journal of Molecular Sciences. 2012; 13(10):12925-12938. https://doi.org/10.3390/ijms131012925
Chicago/Turabian StyleWeiss, Ilana, Claes G. Trope, Reuven Reich, and Ben Davidson. 2012. "Hyaluronan Synthase and Hyaluronidase Expression in Serous Ovarian Carcinoma is Related to Anatomic Site and Chemotherapy Exposure" International Journal of Molecular Sciences 13, no. 10: 12925-12938. https://doi.org/10.3390/ijms131012925
APA StyleWeiss, I., Trope, C. G., Reich, R., & Davidson, B. (2012). Hyaluronan Synthase and Hyaluronidase Expression in Serous Ovarian Carcinoma is Related to Anatomic Site and Chemotherapy Exposure. International Journal of Molecular Sciences, 13(10), 12925-12938. https://doi.org/10.3390/ijms131012925