Screening of Crude Plant Extracts with Anti-Obesity Activity


Abstract
:1. Introduction
2. Experimental Methods
2.1. Chemicals
2.2. Preparation of Natural Extracts
2.3. Pancreatic Lipase Inhibition Assay
2.4. Cell Culture and Differentiation
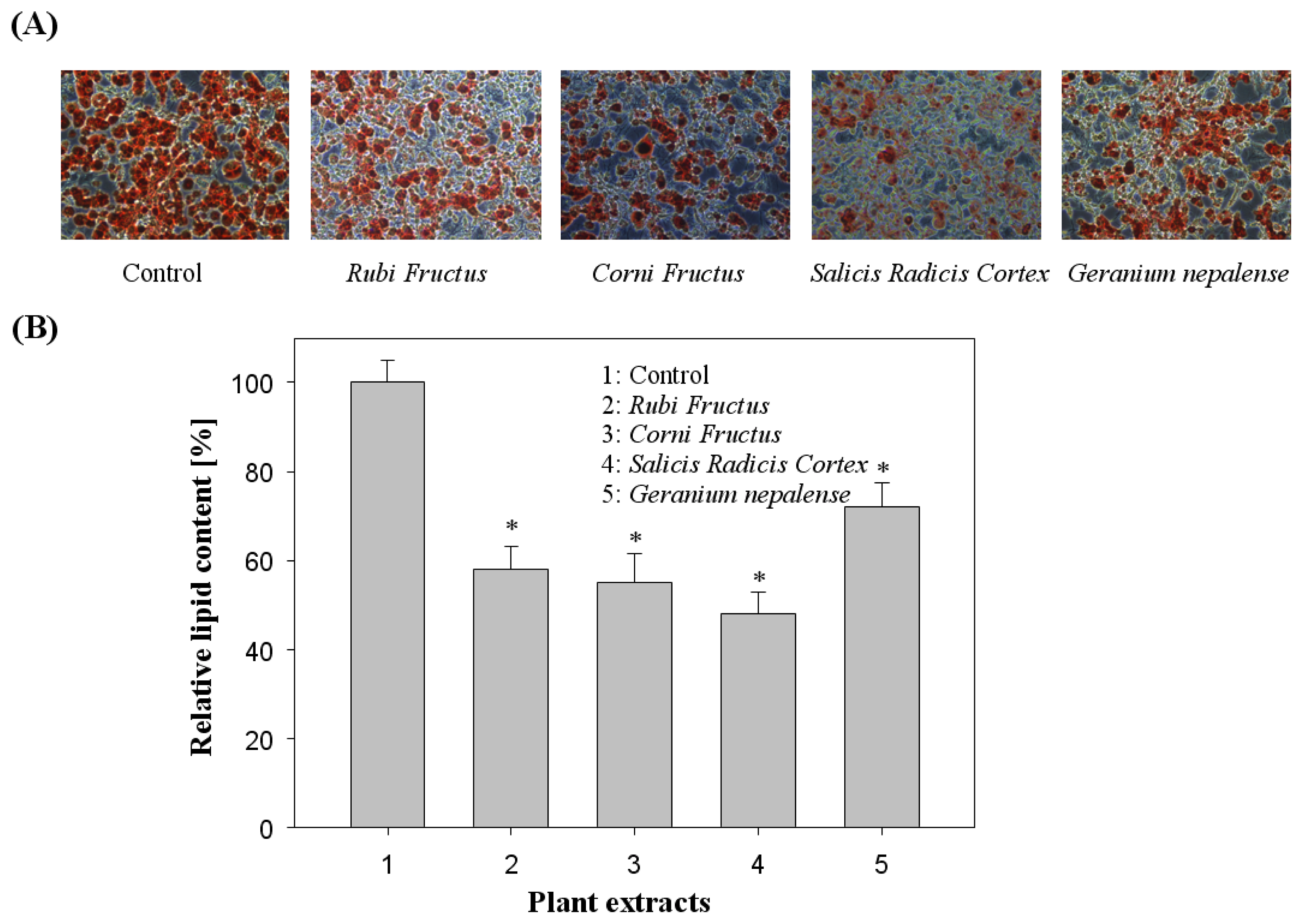
2.5. Cell Viability and Oil Red O Staining Intracellular Triglycerides
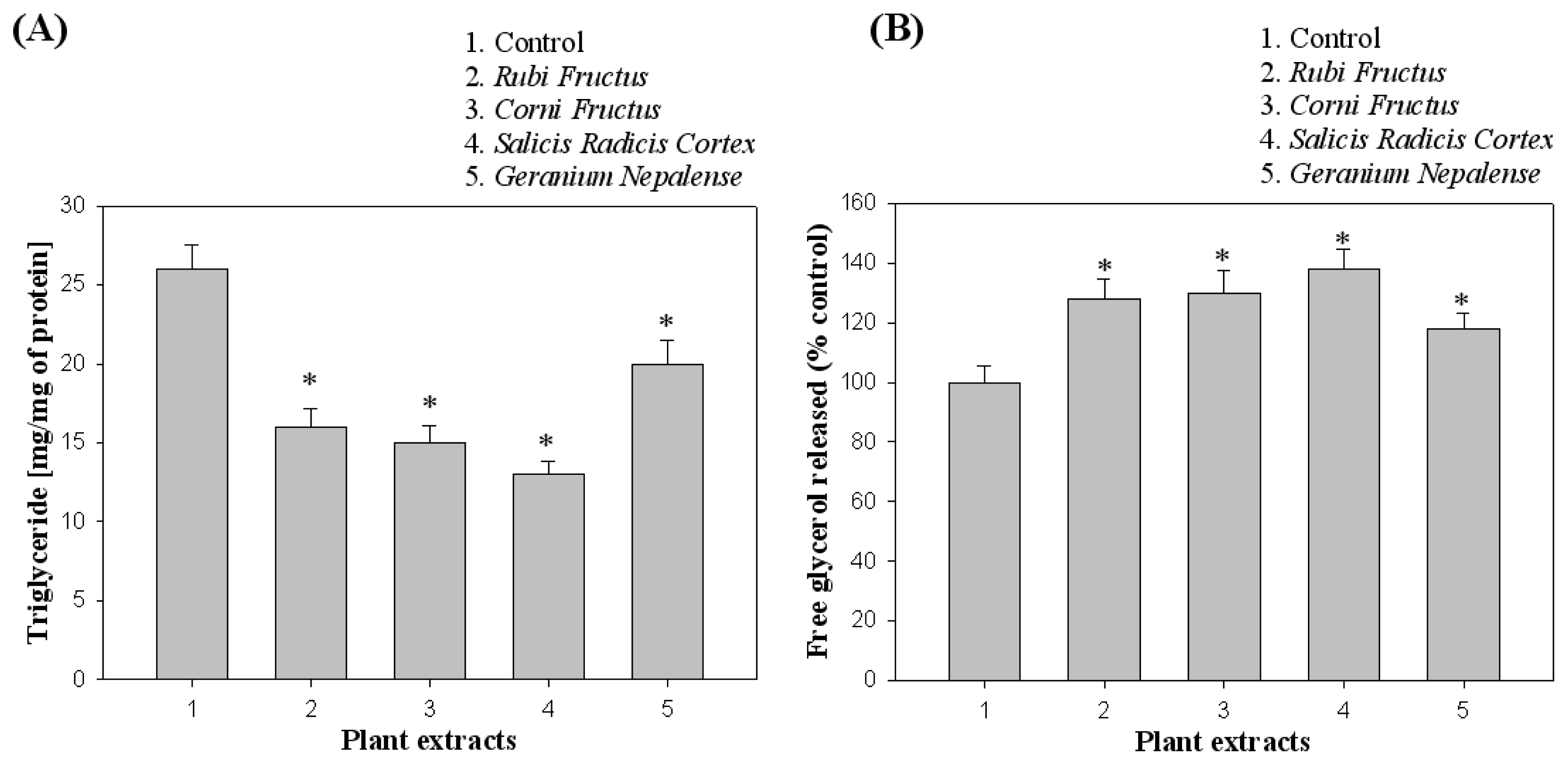
2.6. Measurement of Triglyceride (TG) and Glycerol
2.7. Statistics
3. Results and Discussion
3.1. Anti-Lipase Activity of Crude Natural Source Extracts
3.2. Effect of Cell Viability and Lipid Inhibition in 3T3-L1 Cells
4. Conclusions
Acknowledgments
References
- Spiegelman, B.M.; Flier, J.S. Obesity and the regulation of energy balance. Cell 2001, 104, 531–543. [Google Scholar]
- Kopelman, P.G. Obesity as a medical problem. Nature 2000, 404, 635–643. [Google Scholar]
- Panico, S.; Lannuzzi, A. Dietary fat composition and the metabolic syndrome. Eur. J. Lipid Sci. Technol. 2004, 106, 61–67. [Google Scholar]
- Wakil, S.J.; Abu-Elheiga, L.A. Fatty acid metabolism: Target for metabolic syndrome. J. Lipid Res 2009, S139–143. [Google Scholar]
- Bray, G.A.; Tartaglia, L.A. Medicinal strategies in the treatment of obesity. Nature 2000, 404, 672–677. [Google Scholar]
- Sharma, N.; Sharma, V.K.; Seo, S.Y. Screening of some medicinal plants for anti-lipase activity. J. Ethnopharmacol 2005, 97, 453–456. [Google Scholar]
- Hino, K.; Nagata, H.; Shimonishi, M.; Ido, M. High-throughput screening for small-molecule adiponectin secretion modulators. J. Biomol. Screen 2011, 16, 628–636. [Google Scholar]
- Vázquez-Vela, M.E.F.; Torres, N.; Tovar, A.R. White adipose tissue as endocrine organ and its role in obesity. Arch. Med. Res 2008, 39, 715–728. [Google Scholar]
- Tanzi, M.C.; Fare, S. Adipose tissue engineering: State of the art, recent advances and innovative approaches. Expert Rev. Med. Devices 2009, 6, 533–551. [Google Scholar]
- Miyoshi, H.; Perfield, J.W., II; Obin, M.S.; Greenberg, A.S. Adipose triglyceride lipase regulates basal lipolysis and lipid droplet size in adipocytes. J. Cell. Biochem. 2008, 105, 1430–1436. [Google Scholar]
- Shi, Y.; Burn, P. Lipid metabolism enzymes: Emerging drug targets for the treatment of obesity. Nat. Rev. Drug Discov 2004, 3, 695–710. [Google Scholar]
- Hong, Y.H.; Hishikawa, D.; Miyahara, H.; Tsuzuki, H.; Nishimura, Y.; Gotoh, C.; Choi, K.C.; Hokari, Y.; Takagi, Y.; Lee, H.G.; et al. Up-regulation of adipogenin, an adipocyte plasma transmembrane protein, during adipogenesis. Mol. Cell. Biochem 2005, 276, 133–141. [Google Scholar]
- Reilly, S.M.; Lee, C.H. PPARδ as a therapeutic target in metabolic disease. FEBS Lett 2008, 582, 26–31. [Google Scholar]
- Kos, K.; Wilding, J.P.H. SPARC: A key player in the pathologies associated with obesity and diabetes. Nat. Rev. Endocrinol 2010, 6, 225–235. [Google Scholar]
- Thomson, A.B.; de Pover, A.; Keelan, M.; Jarocka-Cyrta, E.; Clandinin, M.T. Inhibition of lipid absorption as an approach to the treatment of obesity. Meth. Enzymol 1997, 286, 3–41. [Google Scholar]
- Tsujita, T.; Ninomiya, H.; Okuda, H. p-Nitrophenyl butyrate hydrolyzing activity of hormone-sensitive lipase from bovine adipose tissue. J. Lipid Res 1989, 30, 997–1004. [Google Scholar]
- Shizhen, L.; Xiwen, L. Compendium of Materia Medica; Foreign Languages Press: Beijing, China, 2003; Volume 6. [Google Scholar]
- Kim, Y.S.; Lee, Y.M.; Kim, H.; Kim, J.; Jang, D.K.; Kim, J.H.; Kim, J.S. Anti-obesity effect of Morus bombycis root extract: Anti-lipase activity and lipolytic effect. J. Ethnopharmacol 2010, 130, 621–624. [Google Scholar]
- Zheng, C.D.; Duan, Y.Q.; Gao, J.M.; Ruan, Z.G. Screening of anti-lipase properties of 37 traditional Chinese medicinal herbs. J. Chin. Med. Assoc 2010, 73, 319–324. [Google Scholar]
- Yang, Z.; Tu, Y.; Xia, H.; Jie, G.; Chen, X.; He, P. Suppression of free-radicals and protection against H2O2-induced oxidative damage in HPF-1 cell by oxidized phenolic compounds present in black tea. Food Chem 2007, 105, 1349–1356. [Google Scholar]
- Mcgowan, M.W.; Artiss, J.D.; Strandbergh, D.R.; Zak, B. A peroxidase-coupled method for the colorimetric determination of serum triglycerides. Clin. Chem 1983, 29, 538–542. [Google Scholar]
- SPSS, version 19.0; IBM: Chicago, IL, USA, 2010.
- Kim, J.S.; Jeon, W.J.; You, H.J.; Park, M.S.; Ji, G.E. Inhibitory activities of Rubi fructus on digestive enzymes. Food Sci. Biotechnol 2010, 19, 1165–1170. [Google Scholar]
- Park, C.H.; Cho, E.J.; Yokozawa, T. Protection against hypercholesterolemia by Corni fructus extract and its related protective mechanism. J. Med. Food 2009, 12, 973–981. [Google Scholar]
- Bonfili, L.; Cecarini, V.; Amici, M.; Cuccioloni, M.; Angeletti, M.; Keller, J.N.; Eleuteri, A.M. Natural polyphenols as proteasome modulators and their role as anti-cancer compounds. FEBS J 2008, 275, 5512–5526. [Google Scholar]
- Tunon, M.J.; Garcia-Mediavilla, M.V.; Sanchez-Campos, S.; Gonzalez-Gallego, J. Potential of flavonoids as antiinflammatory agents: Modulation of pro-inflammatory gene expression and signal transduction pathways. Curr. Drug Metab 2009, 10, 256–271. [Google Scholar]
- Lugasi, A. Foods fortified with phytosterins: Their role in decreasing serum cholesterol level, their European Community authorization and requirements for placing them on the market. Orv. Hetil 2009, 150, 483–496. [Google Scholar]
- Ovesna, Z.; Vachalkova, A.; Horvathova, K. Taraxasterol and beta-sitosterol: New naturally compounds with chemoprotective/chemopreventive effects. Neoplasma 2004, 51, 407–414. [Google Scholar]
- Plat, J.; Mensink, R.P. Effects of plant sterols and stanols on lipid metabolism and cardiovascular risk. Nutr. Metab. Cardiovasc. Dis 2001, 11, 31–40. [Google Scholar]
- Uprety, Y.; Asselin, H.; Boon, E.K.; Yadav, S.; Shrestha, K.K. Indigenous use and bio-efficacy of medicinal plants in the Rasuwa District, Central Nepal. J. Ethnobiol. Ethnomed 2010, 6, 3–9. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| No. | Latin Name | Scientific Name | Family Name | Plant Part | Inhibition (%) * |
|---|---|---|---|---|---|
| 1 | Platycodi Radix | Platycodon grandiflorum A. De Candolle | Campanulaceae | Root | 3.5 ± 0.7 |
| 2 | Aconiti Tuber | Aconitum carmichaeli Debeaux | Ranunculaceae | Root | 12.1 ± 1.1 |
| 3 | Cannabis Semen | Cannabis sativa Linne | Moraceae | Seed | 7.7 ± 0.2 |
| 4 | Chaenomelis Fructus | Chaenomeles sinensis (Thouin) Koehne | Rosaceae | Fruit | 7.1 ± 0.7 |
| 5 | Actinidiae Fructus | Actinidia chinensis | Actinidiaceae | Fruit | 3.2 ± 0.2 |
| 6 | Tribuli Semen | Tribulus terrestris | Zygophyllaceae | Seed | 2.5 ± 0.6 |
| 7 | Lilie Bulbus | Lilium brownii var. viridulun Baker | Liliaceae | Stem | 6.5 ± 1.2 |
| 8 | Luffae Fructus Retinervus | Luffa cylindrica Roemer | Cucurbitaceae | Fruit | 8.2 ± 1.4 |
| 9 | Crataegi Fructus | Crataegus pinnatifida Bunge var. typica Schneider | Rosaceae | Fruit | 5.5 ± 1.2 |
| 10 | Puerariae Radix | Pueraria thunbergiana Bentham | Leguminosae | Root | 3.2 ± 0.1 |
| 11 | Nardostachyos Rhizoma | Nardostachys chinensis Batal | Valerianaceae | Rhizoma | 6.4 ± 0.5 |
| 12 | Zizyphi Fructus | Zizyphus jujuba Miller var. inermis Rehder | Rhamnaceae | Fruit | 6.6 ± 1.5 |
| 13 | Akebiae Caulis | Akebia quinata Decaisne | Lardizabalaceae | Stem | 11.5 ± 0.1 |
| 14 | Quisqalis Fructus | Quisqualis indica Linné | Combretaceae | Fruit | 11.8 ± 0.3 |
| 15 | Loranthi Ramulus | Loranthus parasticus Merr. | Loranthaceae | Whole grass | 13.5 ± 0.7 |
| 16 | Schizandrae Fructus | Schizandra chinensis Baillon | Schizandraceae | Fruit | 5.8 ± 0.2 |
| 17 | Lonicerae Folium | Lonicera japonica Thunberg | Caprifoliaceae | Stem | 6.4 ± 0.5 |
| 18 | Rehmaniae Radix Preparata | Rehmannia glutinosa | Scrophulariaceae | Root | 6.8 ± 1.1 |
| 19 | Dipsaci Radix | Dipsacus asperoides C. Y. Cheng et T. M. Ai | Dipsacaceae | Root | 12.5 ± 0.3 |
| 20 | Morindae Radix | Morinda officinalis How | Rubiaceae | Root | 3.9 ± 0.7 |
| 21 | Perillae Semen | Perilla sikokiana Nakai | Labiatae | Seed | 8.4 ± 0.3 |
| 22 | Pruni Nakaii Semen | Prunus nakaii Leveille | Rosaceae | Seed | 7.7 ± 0.6 |
| 23 | Meliae Fructus | Melia azedarach Linné var. japonica Makino | Meliaceae | Fruit | 8.6 ± 0.8 |
| 24 | Bletillae Rhizoma | Bletilla striata (Thunberg) Reichenbach fil. | Orchidaceae | Rhizoma | 12.1 ± 0.4 |
| 25 | Hedyotidis Diffusae Herba | Oldenlandia diffusa (Willd.) Roxburgh | Rubiaceae | Whole grass | 1.80 ± 0.4 |
| 26 | Hoelen rubra | Poria cocos wolf | Polyporaceae | Bark | 12.7 ± 0.5 |
| 27 | Gastrodiae Rhizoma | Gastrodia Blume | Orochidaceae | Rhizoma | 8.4 ± 0.8 |
| 28 | Gentianae Scabrae Radix | Gentiana scabra | Sympetalae | Root | 7.5 ± 1.1 |
| 29 | Cuscutae Semen crudus | Cuscuta chinensis Lamark | Convolvulaceae | Seed | 5.8 ± 0.4 |
| 30 | Tetrapanacis Medulla | Tetrapanax papyriferus K. Koch | Araliaceae | Stem | 6.6 ± 0.7 |
| 31 | Fritillariae Bulbus | Fritillaria thunbergii Miquel | Liliaceae | Stem | 8.7 ± 0.5 |
| 32 | Patriniae Radix | Patrinia villosa Jussieu | Valerianaceae | Root | 7.7 ± 1.4 |
| 33 | Scutellariae Radix | Scutellaria baicalenis Georgi | Labiatae | Root | 8.7 ± 0.4 |
| 34 | Astragali Radix | Astragalus membranaceus Bunge | Leguminosae | Root | 7.4 ± 1.4 |
| 35 | Phellodendri Cortex | Phellodendron amurense Ruprecht | Rutaceae | Bark | 11.5 ± 0.2 |
| 36 | Drynariae Rhizoma | Drynaria fortunei Smith. | Polypodiaceae | Rhizoma | 10.5 ± 0.4 |
| 37 | Rubi Fructus | Rubus coreanus Miquel | Rosaceae | Fruit | 32.5 ± 1.1 |
| 38 | Eriobotriae Folium | Eriobotrya japonica Lindley | Rosaceae | Leaf | 9.8 ± 0.3 |
| 39 | Corni Fructus | Cornus officinalis Siebold et Zuccarini | Cornaceae | Fruit | 34.8 ± 2.3 |
| 40 | Forsythiae Fructus | Forsythia koreana Nakai | Oleaceae | Fruit | 5.7 ± 1.2 |
| 41 | Salicis Radicis Cortex | Ulmus darvidian for. Suberose | Ulmaceae | Bark | 38.0 ± 1.9 |
| 42 | Ammomi Tsao-ko Frucuts | Amomum tsao-ko Crevost et Lemaire | Zingiberaceae | Fruit | 7.5 ± 0.6 |
| 43 | Polygoni Avicularis Herba | Polygonum aviculare Linné | Polygonaceae | Whole grass | 21.2 ± 1.7 |
| 44 | Geranium Nepalense | Geranium thunbergii Siebold et Zuccarini | Geraniaceae | Whole grass | 31.4 ± 0.7 |
| 45 | Orlistat | 42.0 ± 2.5 |
© 2012 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Roh, C.; Jung, U. Screening of Crude Plant Extracts with Anti-Obesity Activity. Int. J. Mol. Sci. 2012, 13, 1710-1719. https://doi.org/10.3390/ijms13021710
Roh C, Jung U. Screening of Crude Plant Extracts with Anti-Obesity Activity. International Journal of Molecular Sciences. 2012; 13(2):1710-1719. https://doi.org/10.3390/ijms13021710
Chicago/Turabian StyleRoh, Changhyun, and Uhee Jung. 2012. "Screening of Crude Plant Extracts with Anti-Obesity Activity" International Journal of Molecular Sciences 13, no. 2: 1710-1719. https://doi.org/10.3390/ijms13021710
APA StyleRoh, C., & Jung, U. (2012). Screening of Crude Plant Extracts with Anti-Obesity Activity. International Journal of Molecular Sciences, 13(2), 1710-1719. https://doi.org/10.3390/ijms13021710