Alpha7 Nicotinic Acetylcholine Receptor Is a Target in Pharmacology and Toxicology


Abstract
:1. Introduction
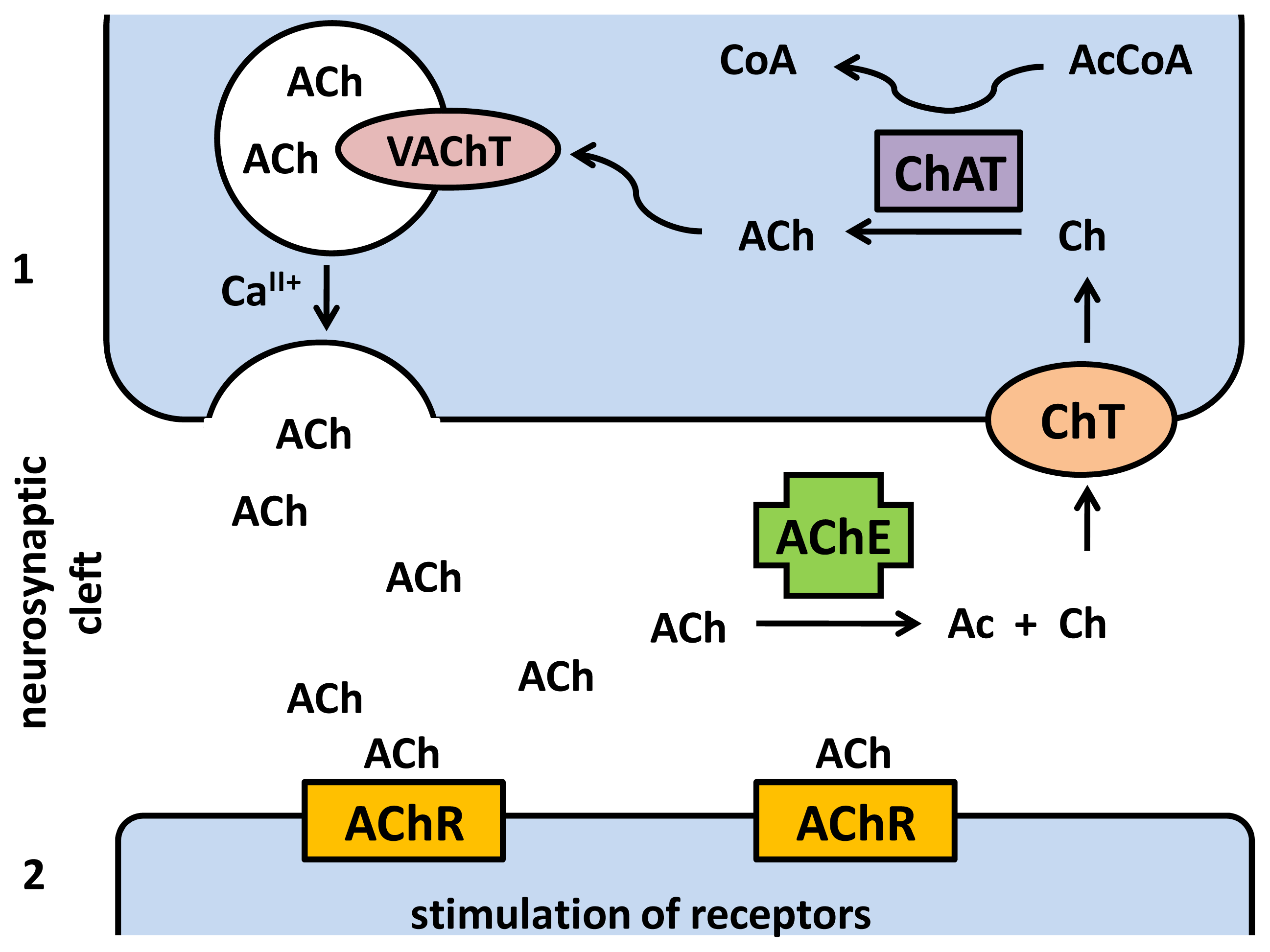
2. Acetylcholine as Neurotransmitter
3. Acetylcholine Receptors
4. α7 Nicotinic Acetylcholine Receptor in Brain
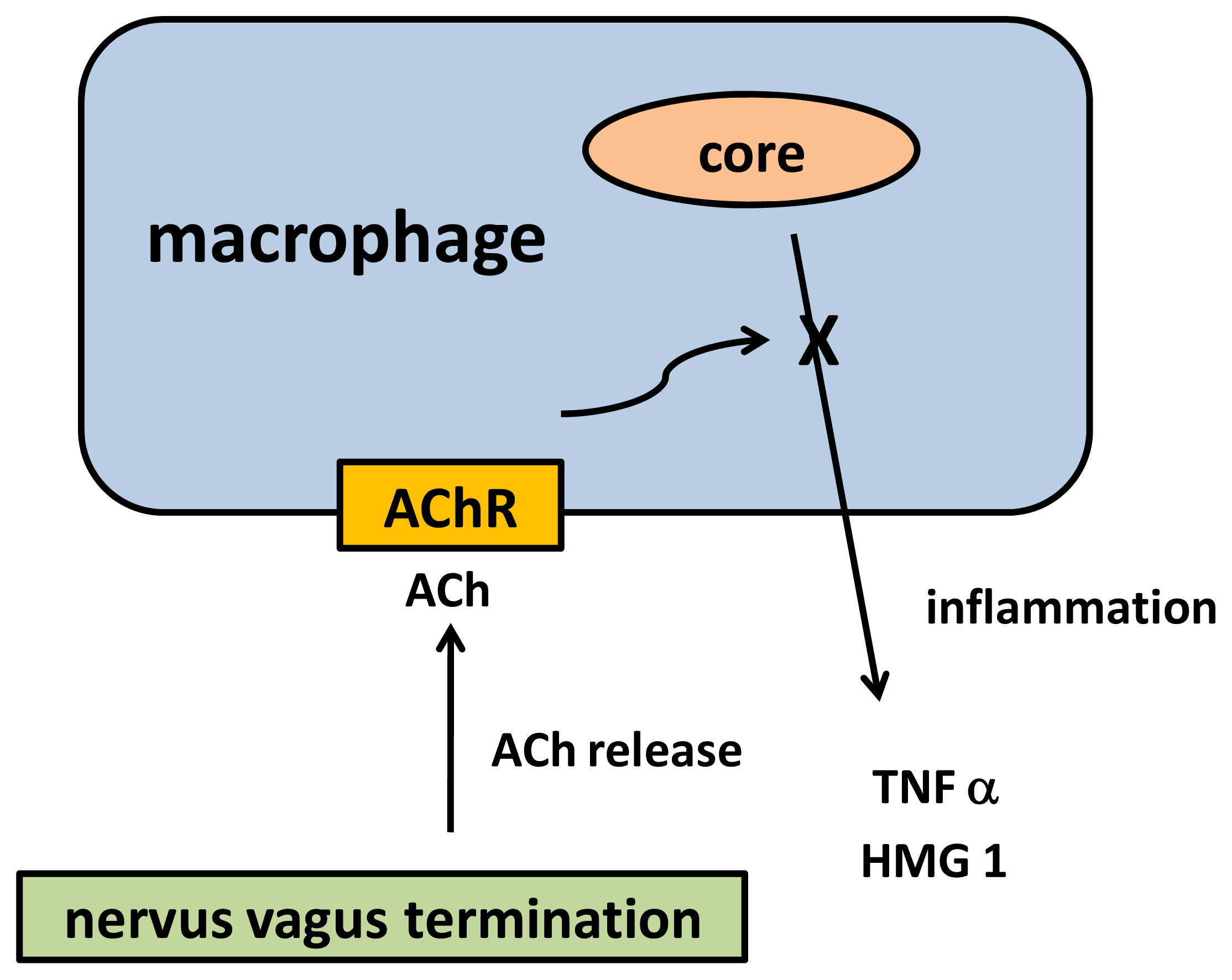
5. Cholinergic Anti-Inflammatory Pathway
6. Antagonists of α7 nAChR
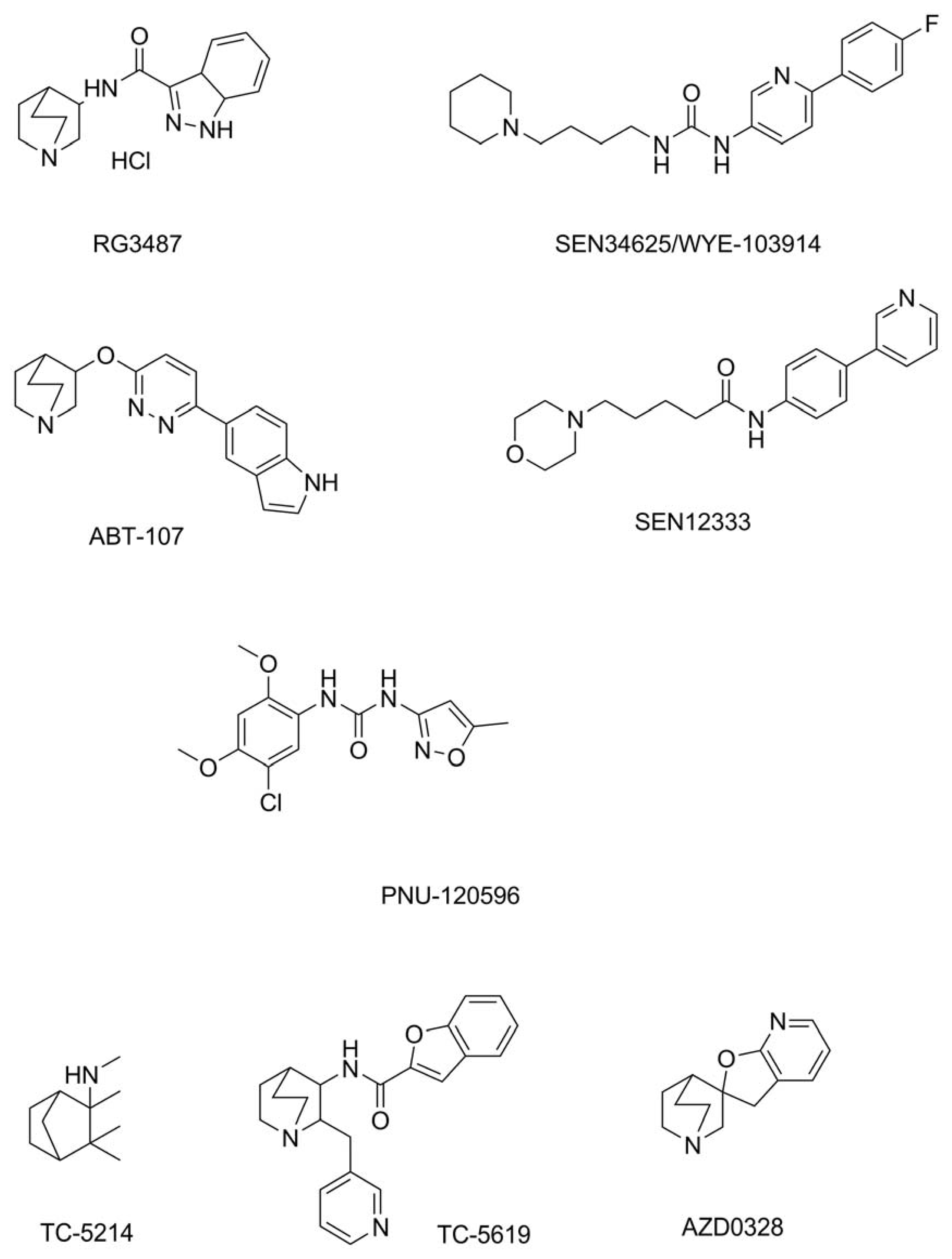
7. Agonists of α7 nAChR
8. Conclusions
Acknowledgments
References
- Rand, J.B. Acetylcholine. WormBook. 2007. http://www.wormbook.org accessed on 17 February 2012. [CrossRef]
- Loewi, O. Uberhumerole ubertragbarkeit der herznervenwirkung. I. Mitt. Pflugers Arch 1921, 189, 239–242. [Google Scholar]
- Wessler, I.; Kirkpatrick, C.J. Acetylcholine beyond neurons: The non-neuronal cholinergic system in humans. Br. J. Pharmacol 2008, 154, 1558–1571. [Google Scholar]
- Gabrielle, P.; Jeana, M.; Lorenza, E.C. Cytosolic choline acetyltransferase binds specifically to cholinergic plasma membrane of rat brain synaptosomes to generate membrane-bound enzyme. Neurochem. Res 2003, 28, 543–549. [Google Scholar]
- Phillis, J.W. Acetylcholine release from the central nervous system: A 50year retrospective. Crit. Rev. Neurobiol 2005, 17, 161–217. [Google Scholar]
- Llona, I. Synaptic like microvesicles: Do they participate in regulated exocytosis? Neurochem. Int 1995, 27, 219–226. [Google Scholar]
- Dun, A.R.; Rickman, C.; Duncan, R.R. The t-SNARE complex: A close up. Cell Mol. Neurobiol 2010, 30, 1321–1326. [Google Scholar]
- Snyder, D.A.; Kelly, M.L.; Woodbury, D.J. SNARE complex regulation by phosphorylation. Cell Biochem. Biophys 2006, 45, 111–123. [Google Scholar]
- Rotundo, R.L. Expression and localization of acetylcholinesterase at the neuromuscular junction. J. Neurocytol 2003, 32, 743–766. [Google Scholar]
- Sarter, M.; Parikh, V. Choline transporters, cholinergic transmission and cognition. Nat. Rev. Neurosci 2005, 6, 48–56. [Google Scholar]
- Hall, J.D.; McCroskey, L.M.; Pincomb, B.J.; Hatheway, C.L. Isolation of an organism resembling Clostridium baratii which produces type F botulinal toxon from an infant with botulism. J. Clin. Microbiol 1985, 21, 654–655. [Google Scholar]
- Aureli, P.; Fenicia, L.; Pasolini, B.; Gianfranceschi, M.; McCroskey, J.M.; Hatheway, C.L. Two cases of type infant botulism caused by neurotoxigenic clostridium butyricum in Italy. J. Infect. Dis 1986, 154, 207–211. [Google Scholar]
- Lacy, D.B.; Tepp, W.; Cohen, A.C.; DasGupta, B.R.; Stevens, R.C. Crystal structure of botulinum neurotoxin type A and implications for toxicity. Nat. Struct. Biol 1998, 5, 898–902. [Google Scholar]
- Singh, B.R. Botulinum neurotoxin structure, engineering, and novel cellular trafficking and targeting. Neurotox. Res 2006, 9, 73–92. [Google Scholar]
- Hirota, S.A. A quick guide to muscarinic acetylcholine receptors. BioPharm. J 2001, 5, 6–8. [Google Scholar]
- Felder, C.C. Muscarinic acetylcholine receptors: Signal transduction through multiple effectors. FASEB J 1995, 9, 619–625. [Google Scholar]
- Tobin, G.; Giglio, D.; Lundgren, O. Muscarinic receptor subtypes in the alimentary track. J. Physiol. Pharmacol 2009, 60, 3–21. [Google Scholar]
- Berstein, G.; Blank, J.L.; Smrcka, A.; Higashijima, T.; Sternweis, P.C.; Exton, J.H.; Ross, E.M. Reconstitution of agonist-stimulated phoshpatidylinostiol 4,5-bisphosphate hydrolysis using purified m1 muscarinic receptor, Gq/11 and phospholipase C-β1. J. Biol. Chem 1992, 267, 8081–8088. [Google Scholar]
- Falkenburger, B.H.; Jensen, J.B.; Hille, B. Kinetics of M1 muscarinic receptor and G protein signaling to phospholipase C in living cells. J. Gen. Physiol 2010, 135, 81–97. [Google Scholar]
- Parker, E.M.; Kameyama, K.; Higashijima, T.; Ross, E.M. Reconstitutively active G protein-coupled receptors purified from baculovirus-infected insect cells. J. Biol. Chem 1991, 266, 519–527. [Google Scholar]
- Alfonzo, M.J.; de Becemberg, I.L.; de Villaroel, S.S.; de Herrerea, V.N.; Misle, A.J.; de Alfonzo, R.G. Two opposite signal transducting mechanisms regulate a G-protein-coupled guanylyl cyclase. Arch. Biochem. Biophys 1998, 350, 19–25. [Google Scholar]
- Horng, H.C.; Chen, F.C.; Ho, C.C.; Kuo, C.P.; Wu, C.T.; Wong, C.S. Bradycardia and hypotension refractory to ephedrine and atropine treatment: Severe autonomic dysfunction with abnormal heart rate variability. Acta Anaesthesiol. Taiwan 2006, 44, 109–112. [Google Scholar]
- Bryant, S.M.; Rhee, J.W.; Thompson, T.M.; Aks, S.E. Pretreating rats with parenteral ophthalmic antimuscarinic agents decreases mortality from lethal organophosphate poisoning. Acad. Emerg. Med 2007, 14, 370–372. [Google Scholar]
- Nachum, Z.; Shupak, A.; Gordon, C.R. Transdermal scopolamine for prevention of motion sickness: Clinical pharmacokinetics and therapeutic applications. Clin. Pharmacokinet 2006, 45, 543–566. [Google Scholar]
- Bridges, T.M.; Lebois, E.P.; Hopkins, C.R.; Wood, M.R.; Jones, C.K.; Conn, P.J.; Lindsley, C.W. The antipsychotic potential of muscarinic allosteric modulation. Drug News Perspect 2010, 23, 229–240. [Google Scholar]
- Woolley, M.L.; Carter, H.J.; Gartlon, J.E.; Watson, J.M.; Dawson, L.A. Attenuation of amphetamine-induced activity by the non-selective muscarinic receptor agonist, xanomeline, is absent in muscarinic M4 receptor knockout mice and attenuated in muscarinic M1 redceptor knockout mice. Eur. J. Pharmacol 2009, 603, 147–149. [Google Scholar]
- Sellin, A.K.; Shad, M.; Tamminga, C. Muscarinic agonists for the treatment of cognition in schizophrenia. CNS Spectr 2008, 13, 985–996. [Google Scholar]
- Nissen, C.; Nofzinger, E.A.; Feige, B.; Waldheim, B.; Radosa, M.P.; Riemann, D.; Berger, M. Differential effects of the muscarinic M1 receptor agonist RS-86 and the acetylcholine-esterase inhibitor donepezil on REM sleep regulation in healthy volunteers. Neuropsychopharmacology 2006, 31, 1294–1300. [Google Scholar]
- Winterer, G. Why do patients with schizophrenia smoke. Curr. Opin. Psychiatry 2010, 23, 112–119. [Google Scholar]
- Williams, J.M.; Gandhi, K.K. Use of caffeine and nicotine in people with schizophrenia. Curr. Drug Abuse Rev 2008, 1, 155–161. [Google Scholar]
- Rommelspacher, H.; Meier-Henco, M.; Smolka, M.; Kloft, C. The levels of norharman are high enough after smoking to affect monoamineoxidase B in platelets. Eur. J. Pharmacol 2002, 441, 115–125. [Google Scholar]
- Nasiripourdori, A.; Taly, V.; Grutter, T.; Taly, A. From toxins targeting ligand gated ion channels to therapeutic molecules. Toxins 2011, 3, 260–293. [Google Scholar]
- Tracey, K.J. Physiology and immunolgy of the cholinergic antiinflammatory pathway. J. Clin. Invest 2007, 117, 289–296. [Google Scholar]
- Millar, N.S. A review of experimental techniques used for the heterologous expression of nicotinic acetylcholine receptors. Biochem. Pharmacol 2009, 78, 766–776. [Google Scholar]
- Lohmann, T.H.; Torrao, A.S.; Britto, L.R.; Lindstrom, J.; Hamassaki-Britto, D.E. A comparative non-radioactive in situ hybridization and immunohistochemical study of the distribution of alpha7 and alpha8 subunits of the nicotinic acetylcholine receptors in visual areas of the chick brain. Brain Res 2000, 852, 463–469. [Google Scholar]
- Unwin, N. Refined Structure of the nicotinic acetylcholine receptor at 4 A resolution. J. Mol. Biol 2005, 346, 967–989. [Google Scholar]
- Albuquerque, E.X.; Pereira, E.F.; Alkondon, M.; Rogers, S.W. Mammalian nicotinic acetylcholine receptors: From structure to function. Physiol. Rev 2009, 89, 73–120. [Google Scholar]
- Alkondon, M.; Pereira, E.F.R.; Cortes, W.S.; Maelicke, A.; Albuquerque, E.X. Choline is a selective agonist of alpha7 nicotnic acetylcholine receptors in the rat brain neurons. Eur. J. Neurosci 1997, 9, 2734–2742. [Google Scholar]
- Zhou, Y.; Nelson, M.E.; Kuryatov, A.; Choi, C.; Cooper, J.; Lindstrom, J. Human α4β2 acetylcholine receptors formed from linked subunits. J. Neurosci 2003, 23, 9004–9015. [Google Scholar]
- Yang, J.J.; Wang, Y.T.; Cheng, P.C.; Kuo, Y.J.; Huang, R.C. Cholinergic modulation of neuronal excitability in the rat suprachiasmatic nucleus. J. Neurophysiol 2010, 103, 1397–1409. [Google Scholar]
- Corringer, P.J.; Bertrand, S.; Galzi, J.L.; Devillers-Thiery, A.; Changeux, J.P.; Bertrand, D. Mutational analysis of the charge selectivity filter of the α7 nicotinic acetylcholine receptor. Neuron 1999, 22, 831–843. [Google Scholar]
- Doley, R.; Kini, R.M. Protein complexes in snake venom. Cell. Mol. Life Sci 2009, 66, 2851–2871. [Google Scholar]
- Gurnaney, H.; Brown, A.; Litman, R.S. Malignant hyperthermia and muscular dystrophies. Anesth. Anal 2009, 109, 1043–1048. [Google Scholar]
- Langeron, O.; Birenbaum, A.; Amour, J. Airway management in trauma. Minerva Anestesiol 2009, 75, 307–311. [Google Scholar]
- Gatke, M.R.; Bundgaard, J.R.; Viby-Mogensen, J. Two novel mutations in the BChE gene in patients with prolonged duration of action of mivacurium or succinylcholine during anaesthesia. Pharmacogenet. Genomics 2007, 17, 995–999. [Google Scholar]
- Leiser, S.C.; Bowlby, M.R.; Comery, T.A.; Dunlop, J. A cog in cognition: How the alpha 7 nicotinic acetylcholine receptor is geared towards improving cognitive deficits. Pharmacol. Ther 2009, 122, 302–311. [Google Scholar]
- Soderman, A.; Mikkelsen, J.D.; West, M.J.; Christensen, D.Z.; Jensen, M.S. Activation of nicotinic α(7) acetylcholine receptor enhances long term potentation in wild type mice but not in APP (swe)/PS1ΔE9 mice. Neurosci. Lett 2011, 487, 325–329. [Google Scholar]
- Thomsen, M.S.; Hansen, H.H.; Timmerman, D.B.; Kikkelsen, J.D. Cognitive improvement by activation of alpha7 nicotinic acetylcholine receptors: From animal models to human pathophysiology. Curr. Pharm. Des 2010, 16, 323–343. [Google Scholar]
- Feher, A.; Juhasz, A.; Rimanoczy, A.; Csibri, E.; Kalman, J.; Janka, Z. Association between a genetic variant of the alpha-7 nicotinic acetylcholine receptor subunit and four types of dementia. Dement. Geriatr. Cogn. Disord 2009, 28, 56–62. [Google Scholar]
- Chu, L.W.; Ma, E.S.; Lam, K.K.; Chan, M.F.; Lee, D.H. Increased alpha 7 nicotinic acetylcholine receptor protein levels in Alzheimer’s disease patients. Dement. Geriatr. Cogn. Disord 2005, 19, 106–112. [Google Scholar]
- Tregellas, J.R.; Tanabe, J.; Rojas, D.C.; Shatti, S.; Olincy, A.; Johnson, L.; Martin, L.F.; Soti, F.; Kem, W.R.; Leonard, S.; et al. Effects of an alpha 7-nicotinic agonist on default network activity in schizophrenia. Biol. Psychiatry 2011, 69, 7–11. [Google Scholar]
- Livingston, P.D.; Srinivasan, J.; Kew, J.N.; Dawson, L.A.; Gotti, C.; Moretti, M.; Shoaib, M.; Wonnacott, S. Alpha7 and non-alpha7 nicotinic acetylcholine receptors modulate dopamine release in vitro and in vivo in the rat prefrontal cortex. Eur. J. Neurosci 2009, 29, 539–550. [Google Scholar]
- Martin, L.F.; Kem, W.R.; Freedman, R. Alpha-7 nicotinic receptor agonists: Potential new candidates for the treatment of schizophrenia. Psychopharmacology 2004, 174, 54–64. [Google Scholar]
- Picciotto, M.R.; Caldarone, B.J.; Brunzell, D.H.; Zachariou, V.; Stevens, T.R.; King, S.L. Neuronal nicotinic acetylcholine receptor subunit knockout mice: Physiological and behavioral phenotypes and possible clinical inplications. Pharmacol. Ther 2001, 92, 89–108. [Google Scholar]
- Dziewczapolski, G.; Glogowski, C.M.; Masliah, E.; Heinemann, S.F. Deletion of the alpha 7 nicotinic acetylcholine receptor gene improves cognitive deficits and synaptic pathology in a mouse model of Alzheimer’s disease. J. Neurosci 2009, 29, 8805–8815. [Google Scholar]
- Lester, H.A.; Fonck, C.; Tapper, A.R.; McKinney, S.; Damaj, M.I.; Balogh, S.; Owens, J.; Wehner, J.M.; Collins, A.C.; Labarca, C. Hypersensitive knockin mouse strains identify receptors and pathways for nicotine action. Curr. Opin. Drug Discov. Devel 2003, 6, 633–639. [Google Scholar]
- Tracey, K.J. The inflammatory reflex. Nature 2002, 420, 853–859. [Google Scholar]
- Borovikova, L.V.; Ivanova, S.; Zhang, M.; Yang, H.; Botchkina, G.I.; Watkins, L.R.; Wang, H.; Abumrad, N.; Eaton, J.W.; Tracey, K.J. Vagus nerve stimulation attenuates the systemic inflammatory response to endotoxin. Nature 2000, 405, 458–462. [Google Scholar]
- Tracey, K.J. Reflex control of immunity. Nat. Rev. Immunol 2009, 9, 418–428. [Google Scholar]
- Tracey, K.J. Fat meets the cholinergic antiinflammatory pathway. J. Exp. Med 2005, 202, 1071–1021. [Google Scholar]
- Casserly, B.; Baram, M.; Walsh, P.; Sucov, A.; Ward, N.S.; Levy, M.M. Implementing a collaborative protocol in a sepsis intervention program: Lessons learned. Lung 2011, 189, 11–19. [Google Scholar]
- Babaev, V.R.; Patel, M.B.; Semenkovich, C.F.; Fazio, S.; Linton, M.F. Macrophage lipoprotein lipase promotes foam cell formation and atherosclerosis in low density lipoprotein receptor-deficient mice. J. Biol. Chem 2000, 275, 26293–26299. [Google Scholar]
- Herbein, G.; Gras, G.; Khan, K.A.; Abbas, W. Macrophage signaling in HIV-1 infection. Retrovirology 2010, 7, 34:1–34:13. [Google Scholar]
- Pohanka, M.; Snopkova, S.; Havlickova, K.; Bostik, P.; Sinkorova, Z.; Fusek, J.; Kuca, K.; Pikula, J. Macrophage-assisted inflammation and pharmacological regulation of the cholinergic anti-inflammatory pathway. Curr. Med. Chem 2011, 18, 539–551. [Google Scholar]
- Parrish, W.R.; Gallowitsch-Puerta, M.; Czura, C.J.; Tracey, K.J. Experimental therapeutic strategies for severe sepsis: Mediators and mechanisms. Ann. N. Y. Acad. Sci 2008, 1144, 210–236. [Google Scholar]
- Xiong, J.; Xue, F.S.; Yuan, Y.J.; Wang, Q.; Liao, X.; Wang, W.L. Cholinergic anti-inflammatory pathway: A possible approach to protect against myocardial ischemia reperfusion injury. Chin. Med. J. (Engl.) 2010, 123, 2720–2726. [Google Scholar]
- Zhou, Y.; Zuo, X.; Li, Y.; Wang, Y.; Zhao, H.; Xiao, X. Nicotine inhibits tumor necrosis factor-alpha induced IL-6 and IL-8 secretion in fibroblast-like synoviocytes from patients with rheumatoid arthritis. Rheumatol. Int 2012, 32, 97–104. [Google Scholar]
- Azam, L.; Mcintosh, J.M. Alpha-conotoxins as pharmacological probes of nicotinic acetylholine receptors. Acta Pharmacol. Sin 2009, 30, 771–783. [Google Scholar]
- Millard, E.L.; Daly, N.L.; Craik, D.J. Structure-activity relationships of alpha-conotoxins targeting neuronal nicotinic acetylcholine receptors. Eur. J. Biochem 2004, 271, 2320–2326. [Google Scholar]
- Favreau, P.; Krimm, I.; Le Gall, F.; Bobenrieth, M.J.; Lamthanh, H.; Bouet, F.; Servent, D.; Molgo, J.; Menez, A.; Letouneux, Y.; et al. Biochemical characterization and nuclear magnetic resonance structure of novel alpha-conotoxins isolated from the venom of conus consors. Biochemistry 1999, 38, 6317–6326. [Google Scholar]
- Hogg, R.C.; Miranda, L.P.; Craik, D.J.; Lewis, R.J.; Alewood, P.F.; Adams, D.J. Single amino acid substitutions in alpha-conotoxin PnIA shift selectivity for subtypes of the mammalian neuronal nicotinic acetylcholine receptor. J. Biol. Chem 1999, 274, 36559–36564. [Google Scholar]
- Dutertre, S.; Nicke, A.; Lewis, R.J. Beta2 subunit contribution to 4/7 alpha-conotoxin binding to the nicotinic acetylchline receptor. J. Biol. Chem 2005, 280, 30460–30468. [Google Scholar]
- Blanchfield, J.T.; Gallagher, O.P.; Cros, C.; Lewis, R.J.; Alewood, P.F.; Toth, I. Oral absorption and in vivo biodistribution of alpha-conotoxin MII and a lipidic analogue. Biochem. Biophys. Res. Commun 2007, 361, 97–102. [Google Scholar]
- Whiteaker, P.; Mcintosh, J.M.; Luo, S.; Collins, A.C.; Marks, M.J. 125I-alpha-conotoxin MII identifies a novel nicotinic acetylcholine receptor population in mouse brain. Mol. Pharmacol 2000, 57, 913–925. [Google Scholar]
- Arias, H.R. Localization of agonist and competitive antagonist binding sites on nicotinic acetylcholine receptors. Neurochem. Int 2000, 36, 595–645. [Google Scholar]
- McCann, C.M.; Bracamontes, J.; Steinbach, J.H.; Sanes, J.R. The cholinergic antagonist alpha-bungarotoxin also binds and blocks a subset of GABA receptors. Proc. Natl. Acad. Sci. USA 2006, 103, 5149–5154. [Google Scholar]
- Chang, C.C. Looking back on the discovery of alpha-bungarotoxin. J. Biomed. Sci 1999, 6, 368–375. [Google Scholar]
- Hawgood, B.J. Professor Chen-Yuan Lee, MD (1915–2001), pharmacologist: Snake venom research at the Institute of Pharmacology, National Taiwan University. Toxicon 2002, 40, 1065–1072. [Google Scholar]
- Chu, N.S. Contribution of a snake venom toxin to myasthenia gravis: The discovery of alpha-bungarotoxin in Taiwan. J. Hist. Neurosci 2005, 14, 138–148. [Google Scholar]
- Aracava, Y.; Pereira, E.F.; Maelicke, A.; Albuquerque, E.X. Memantine blocks alpha7* nicotnic acetylcholine receptors more potently than N-methyl-D-aspartate receptors in rat hippocampal neurons. J. Pharmacol. Exp. Ther 2005, 312, 1195–1205. [Google Scholar]
- Banerjee, P.; Samoriski, G.; Gupta, S. Comments on “Memantine blocks alpha7* nicotnic acetylcholine receptors more potently than N-methyl-D-aspartate receptors in rat hippocampal neurons”. J. Pharmacol. Exp. Ther 2005, 313, 928–929. [Google Scholar]
- Taly, A.; Corringer, P.J.; Guedin, D.; Lestage, P.; Changeux, J.P. Nicotinic receptors: Allosteric transitions and therapeutic targets in the nervous system. Nat. Rev. Drug Discov 2009, 8, 733–750. [Google Scholar]
- Santos, M.D.; Alkondon, M.; Pereira, E.F.; Aracava, Y.; Eisenberg, H.M.; Maelicke, A.; Albuquerque, E.X. The nicotinic allosteric potentiating ligand galantamine facilitates synaptic transmission in the mammalian central nervous system. Mol. Pharmacol 2002, 61, 1222–1234. [Google Scholar]
- Schedel, A.; Thornton, S.; Schloss, P.; Kluter, H.; Bugert, P. Human platelets express functional alpha 7-nicotinic acetylcholine receptors. Arterioscler. Thromb. Vasc. Biol 2011, 31, 928–934. [Google Scholar]
- Kassa, J. Review of oximes in the antidotal treatment of poisoning by organophoshorus nerve agents. J. Toxicol. Clin. Toxicol 2002, 40, 803–816. [Google Scholar]
- Pohanka, M.; Jun, D.; Kuca, K. Amperometric biosensor for evaluation of competitive cholinesterase inhibition by the reactivation HI-6. Anal. Lett 2007, 40, 2351–2359. [Google Scholar]
- Soukup, O.; Pohanka, M.; Tobin, G.; Jun, D.; Fusek, J.; Musilek, K.; Marek, J.; Kassa, J.; Kuca, K. The effect of HI-6 on cholinesterases and on the cholinergic system of the rat bladder. Neuroendocrinol. Lett 2008, 29, 759–762. [Google Scholar]
- Pohanka, M.; Pavlis, O.; Pikula, J.; Treml, F.; Kuca, K. Modulation of tularemia disease progress by the bisquaternary pyridinium oxime HI-6. Acta Vet. (Brno) 2010, 79, 443–448. [Google Scholar]
- Pohanka, M.; Pejchal, J.; Horackova, S.; Kuca, K.; Bandouchova, H.; Damkova, V.; Pikula, J. Modulation of ionising radiation generated oxidative stress by HI-6 (asoxime) in a laboratory rat model. Neuroendocrinol. Lett 2010, 31, 62–68. [Google Scholar]
- Tsuji, A. Small molecular drug transfer across the blood-brain barrier via carrier-mediated transport system. NeuroRx 2005, 2, 54–62. [Google Scholar]
- Liu, X.; Testa, B.; Fahr, A. Lipophilicity and its relationship with passive drug permeation. Pharm. Res 2011, 28, 962–977. [Google Scholar]
- Ohtsuki, S.; Terasaki, T. Contribution of carrier-mediated transport system to the blood-brain barrier as a supporting and protecting interface for the brain, importance for CNS drug discovery and development. Pharm. Res 2007, 24, 1745–1758. [Google Scholar]
- Wallace, T.L.; Callahan, P.M.; Tehim, A.; Bertrand, D.; Tombaugh, G.; Wang, S.; Xie, W.; Rowe, W.B.; Ong, V.; Graham, E.; et al. RG3487, a novel nicotinic alpha7 receptor partial agonist, improves cognition and sensorimotor gating in rodents. J. Pharmacol. Exp. Ther 2011, 336, 242–253. [Google Scholar]
- Chiron, C.; Haydar, S.N.; Aschmies, S.; Bothmann, H.; Castaldo, C.; Cocconcelli, G.; Comery, T.A.; Di, L.; Dunlop, J.; Lock, T.; et al. Novel alpha-7 nicotinic acetylcholine receptor agonists containing a urea moiety: Identification and characterization of the potent, selective, and orally efficacious agonist 1-[6-(4-fluorophenyl)pyridin-3-yl]-3-(4-piperidin-1-ylbutyl) urea (SEN34625/WYE-103914). J. Med. Chem 2010, 53, 4379–7389. [Google Scholar]
- Roncarati, R.; Scali, C.; Comery, T.A.; Grauer, S.M.; Aschmi, S.; Bothmann, H.; Jow, B.; Kowal, D.; Gianfriddo, M.; Kelley, C.; et al. Procognitive and neuroprotective activity of novel alpha 7 nicotinic acetylcholine receptor agonist for treatment of neurodegenerative and cognitive disorders. J. Pharmacol. Exp. Ther 2009, 329, 459–468. [Google Scholar]
- Malysz, J.; Anderson, D.J.; Gronlien, J.H.; Ji, J.; Bunnelle, W.H.; Hakerud, M.; Thorin-Hagene, K.; Ween, H.; Helfrich, R.; Hu, M.; et al. In vitro phramacological characterization of a novel selective alpha7 neuronal nicotinic acetylcholine receptor agonist ABT-107. J. Pharmacol. Exp. Ther 2010, 334, 863–874. [Google Scholar]
- Asenjo Lobos, C.; Komossa, K.; Rummel-Kluge, C.; Hunger, H.; Schmid, F.; Schwarz, S.; Leucht, S. Clozapine versus other atypical antipsychotics for schizophrenia. Cochrane Database Syst. Rev 2010, 83. [Google Scholar] [CrossRef]
- Bertrand, D.; Gopalakrishnan, M. Allosteric modulation of nicotinic acetlycholine receptors. Biochem. Pharmacol 2007, 74, 1155–1163. [Google Scholar]
- Kalappa, B.I.; Gusev, A.G.; Uteshev, V.V. Activation of functional α7-containing nAChRs in hippocampal CA1 pyrmidal neurons by physiologica levels of choline in the presence of PNU-120596. PLoS One 2010, 5. [Google Scholar] [CrossRef]
- Lippiello, P.M.; Beaver, J.S.; Gatto, G.J.; James, J.W.; Jordan, K.G.; Traina, V.M.; Xie, J.; Benchrif, M. TC-5214 (S-(+)-mecamylamine): A neuronal nicotinic receptor modulator with antidepressant activity. CNS Neurosci. Ther 2008, 14, 266–277. [Google Scholar]
- Sydserff, S.; Sutton, E.J.; Song, D.; Quirk, M.C.; Maciag, C.; Li, C.; Jonak, G.; Gurley, D.; Gordon, J.C.; Christian, E.P.; et al. Selective alpha7 nicotinic receptor activation by AZD0328 enhances cortical dopamine release and improves learning and attentional processes. Biochem. Pharmacol 2009, 78, 880–888. [Google Scholar]
- Hauser, T.A.; Kucinski, A.; Jordan, K.G.; Gatto, G.J.; Wersinger, S.R.; Hesse, R.A.; Stachowiak, E.K.; Stachowiak, M.K.; Papke, R.L.; Lippiello, P.M.; et al. TC-5619: An alpha7 neuronal nicotinic receptor-selective agonist that demonstrates efficacy in animal models of the positive and negative symptoms and cognitive dysfunction of schizophrenia. Biochem. Pharmacol 2009, 78, 803–812. [Google Scholar]
- Jonnala, R.R.; Grahama, J.H.; Terry, A.V.; Beach, J.W.; Young, J.A.; Buccafusco, J.J. Relative level of cytoprotection produced by analogs of choline and the role of alpha 7-nicotinic acetylcholine receptors. Synapse 2003, 47, 262–269. [Google Scholar]
- Pohanka, M. Cholinesterases, a target of pharmacology and toxicology. Biomed. Pap. Olomouc 2011, 155, 219–223. [Google Scholar]
- Minutoli, L.; Squadrito, F.; Nicotina, P.A.; Giuliani, D.; Ottani, A.; Polito, F.; Bitto, A.; Irrera, N.; Guzzo, G.; Spaccapelo, L.; et al. Melanocortin 4 receptor stimulation decreases pancreatitis severity in rats by activation of the cholinergic anti-inflammatory pathway. Crit. Care Med 2011, 39, 1089–1096. [Google Scholar]
- Rosas-Ballina, M.; Tracey, K.J. Cholinergic control of inflammation. J. Intern. Med 2009, 265, 663–679. [Google Scholar]
- Rosas-Ballina, M.; Goldstein, R.S.; Gallowitsch-Puerta, M.; Yang, L.; Valdes-Ferrer, S.I.; Patel, N.B.; Chavan, S.; Al-Abed, Y.; Yang, H.; Tracey, K.J. The selective alpha7 agonist GTS-21 attenuates cytokine production in human whole blood and human monocytes activated by ligands for TLR2, TLR3, TLR4, TLR9, and RAGE. Mol. Med 2009, 15, 195–202. [Google Scholar]
- Pavlov, V.A.; Ochani, M.; Yang, L.H.; Gallowitsch-Puerta, M.; Ochani, K.; Lin, X.; Levi, J.; Parrish, W.R.; Rosas-Ballina, M.; Czura, C.J.; et al. Selective alpha7-nicotinic acetylcholine receptor agonist GTS-21 improves survival in murine endotoxemia and severe sepsis. Crit. Care Med 2007, 35, 1139–1144. [Google Scholar]
- Kox, M.; Pompe, J.C.; Peters, E.; VAneker, M.; van der Laak, J.W.; van der Hoeven, J.G.; Scheffer, G.J.; Hoedemaekers, C.W.; Pickkers, P. α7 nicotinic acetylcholine receptor agonist GTS-21 attenuates ventilator-induced tumour necrosis factor-α production and lung injury. Br. J. Anaesth 2011, 107, 559–566. [Google Scholar]
- Tregellas, J.R.; Tanabe, J.; Rojas, D.C.; Shatti, S.; Olincy, A.; Johnson, L.; Martin, L.F.; Soti, F.; Kem, W.R.; Leonard, S.; Freedman, R. Effects of an alpha 7-nicotinic agonist on default network activity in schizophrenia. Biol. Psychiatry 2011, 69, 7–11. [Google Scholar]
- Bowman, G.; Bonneau, R.H.; Chinchilli, V.M.; Tracey, K.J.; Cockroft, K.M. A novel inhibitor of inflammatory cytokine production (CNI-1493) reduces rodent post-hemorrhagic vasospasm. Neurocit. Care 2006, 5, 222–229. [Google Scholar]
- Oke, S.L.; Tracey, K.J. From CNI-1493 to the immunological homunculus: Physiology of the inflammatory reflex. J. Leukoc. Biol 2008, 83, 512–517. [Google Scholar]
- Bacher, M.; Dodel, R.; Aljabari, B.; Keyvani, K.; Marambaud, P.; Kayed, R.; Glabe, C.; Goertz, N.; Hoppmann, A.; Sachser, N.; et al. CNI-1493 inhibits Abeta production, and cognitive deterioration in an animal model of Alzheimer’s disease. J. Exp. Med 2008, 205, 1593–1599. [Google Scholar]
- Oettinger, C.W.; D’Souza, M.J. Synergism in survival to endotoxic shock in rats given microencapsulated CNI-1493 and antisense oligomers to NF-kappaB. J. Microencapsul 2010, 27, 372–376. [Google Scholar]
- Dotan, I.; RAchmilewitz, D.; Schreiber, S.; Eliakim, R.; van der Woude, C.J.; Kornbluth, A.; Buchman, A.L.; Bar-Meir, S.; Bokemeyer, B.; Goldin, E.; et al. A randomised placebo-controlled multicentre trial of intravenous semapimod HCl for moderate to severe Crohn’s disease. Gut 2010, 59, 760–766. [Google Scholar]
- The, F.O.; Boeckxstaens, G.E.; Snoek, S.A.; Cash, J.L.; Bennink, R.; Larosa, G.J.; van den Wijngaard, R.M.; Greaves, D.R.; de Jonge, W.J. Activation of the cholinergic anti-inflammatory pathway ameliorates postoperative ileus in mice. Gastroenterology 2007, 133, 1219–1228. [Google Scholar]
- Van Kampen, M.; Selbach, K.; Schneider, R.; Schiegel, E.; Boess, F.; Schreiber, R. AR-R 17779 improves social recognition in rats by activation of nicotinic alpha7 receptors. Psychopharmacology 2004, 172, 375–383. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Structure | Source | Use | Reference |
|---|---|---|---|---|
| α-bungarotoxin | 8 kDa globular protein | Taiwanese krait Bungarus multicinctus | not used in therapy, natural toxin | [77–79] |
| CnIA, PnIA | polypeptide | α-conotoxins from cone snail Conus consors | natural toxin, use in research for distinguishing of acetylcholine receptors types | [70–75] |
| HI-6 (also known as asoxime) | 1-[(4-carbamoylpyridin-1-ium- 1-yl)methoxymethyl]pyridin-1- ium-4- carboxamide dichloride CAS: 34433-31-3 | byspyridinium oxime derivative | therapy of nerve agents poisoning via reactivation of AChE | [85–89] |
| memantine | 3,5-Dimethyltricyclo[ 3.3.1.13,7]decan-1- amine hydrochloride CAS: 19982-08-2 | adamantane derivative | Alzheimer’s disease drug antagonizing NMDA receptor, antagonism of α7 nAChR is a side pathway | [80–82] |
| methylcaconitine | 683 Da alkaloid CAS: 21019-30-7 | Consolida flowers | not used in therapy, natural toxin | [83–84] |
| Name | Structure | Use | Reference |
|---|---|---|---|
| ABT-107 | 5-(6-[(3R)-1- azabicyclo[2.2.2]oct-3- yloxy]pyridazin-3-yl)-1H-indole | Treatment of Alzheimer’s disease and cognitive deficits associated with schizophrenia, under testing, not comercially available | [96] |
| SEN12333 | 5-morpholin-4-yl-pentanoic acid (4-pyridin-3-yl-phenyl)-amide | [95] | |
| TC-5619 | N-[(2S,3S)-2-(pyridin-3- ylmethyl)-1-azabicyclo[2.2.2]oct- 3-yl]-1-benzofuran-2- carboxamide | [102] | |
| Clozapine | 8-chloro-11-(4-methylpiperazin- 1-yl)-5Hdibenzo[ b,e][1,4]diazepine | Commercially available drug (trade names Azaleptin, Clozaril, FazaClo, Leonex and others), used as antipsychotics in paranoid disorders and schizophrenia | [51,53] |
| CNI-1493 | N,N′-bis[3,5-bis[N- (diaminomethylideneamino)-Cmethylcarbonimidoyl] phenyl] decanediamide tetrahydrochloride | Inhibitor of inflammation and NO production, antagonist of α7 nAChR via cholinergic anti-inflammatory pahtway, known as a drug Semapimod, under clinical trials | [112–115] |
| GTS-21 (or DMXB-A) | 3-[(3E)-3-[(2,4- dimethoxyphenyl)methylidene]-5,6-dihydro-4H-pyridin-2-yl] pyridine | Treatment of Alzheimer’s disease and cognitive deficits associated with schizophrenia, experimental testing for anti-inflammatory potency, under clinical testing | [107–110] |
© 2012 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Pohanka, M. Alpha7 Nicotinic Acetylcholine Receptor Is a Target in Pharmacology and Toxicology. Int. J. Mol. Sci. 2012, 13, 2219-2238. https://doi.org/10.3390/ijms13022219
Pohanka M. Alpha7 Nicotinic Acetylcholine Receptor Is a Target in Pharmacology and Toxicology. International Journal of Molecular Sciences. 2012; 13(2):2219-2238. https://doi.org/10.3390/ijms13022219
Chicago/Turabian StylePohanka, Miroslav. 2012. "Alpha7 Nicotinic Acetylcholine Receptor Is a Target in Pharmacology and Toxicology" International Journal of Molecular Sciences 13, no. 2: 2219-2238. https://doi.org/10.3390/ijms13022219
APA StylePohanka, M. (2012). Alpha7 Nicotinic Acetylcholine Receptor Is a Target in Pharmacology and Toxicology. International Journal of Molecular Sciences, 13(2), 2219-2238. https://doi.org/10.3390/ijms13022219