Improvement of Carbon Tetrachloride-Induced Acute Hepatic Failure by Transplantation of Induced Pluripotent Stem Cells without Reprogramming Factor c-Myc

Abstract
:1. Introduction
2. Results and Discussion
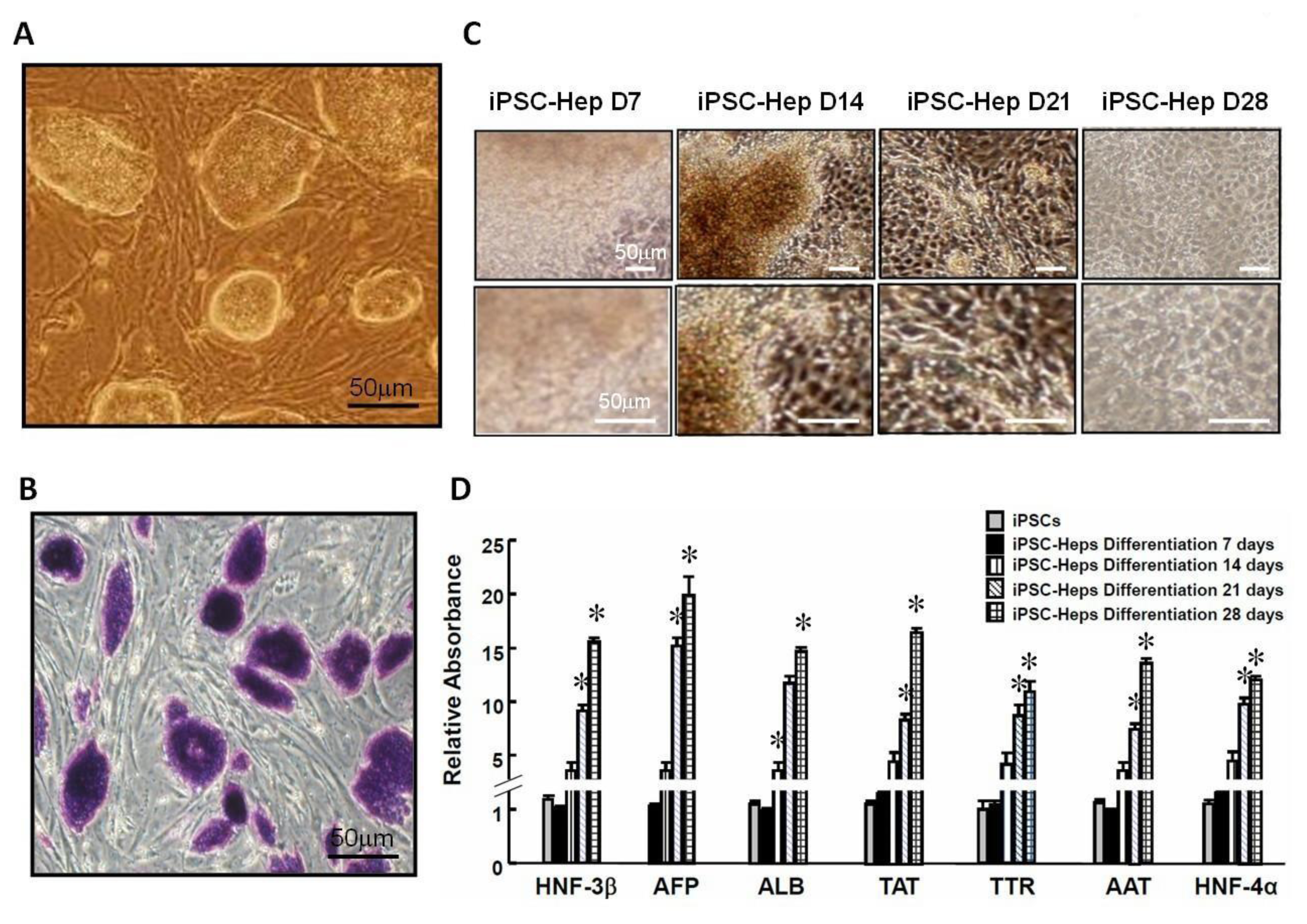
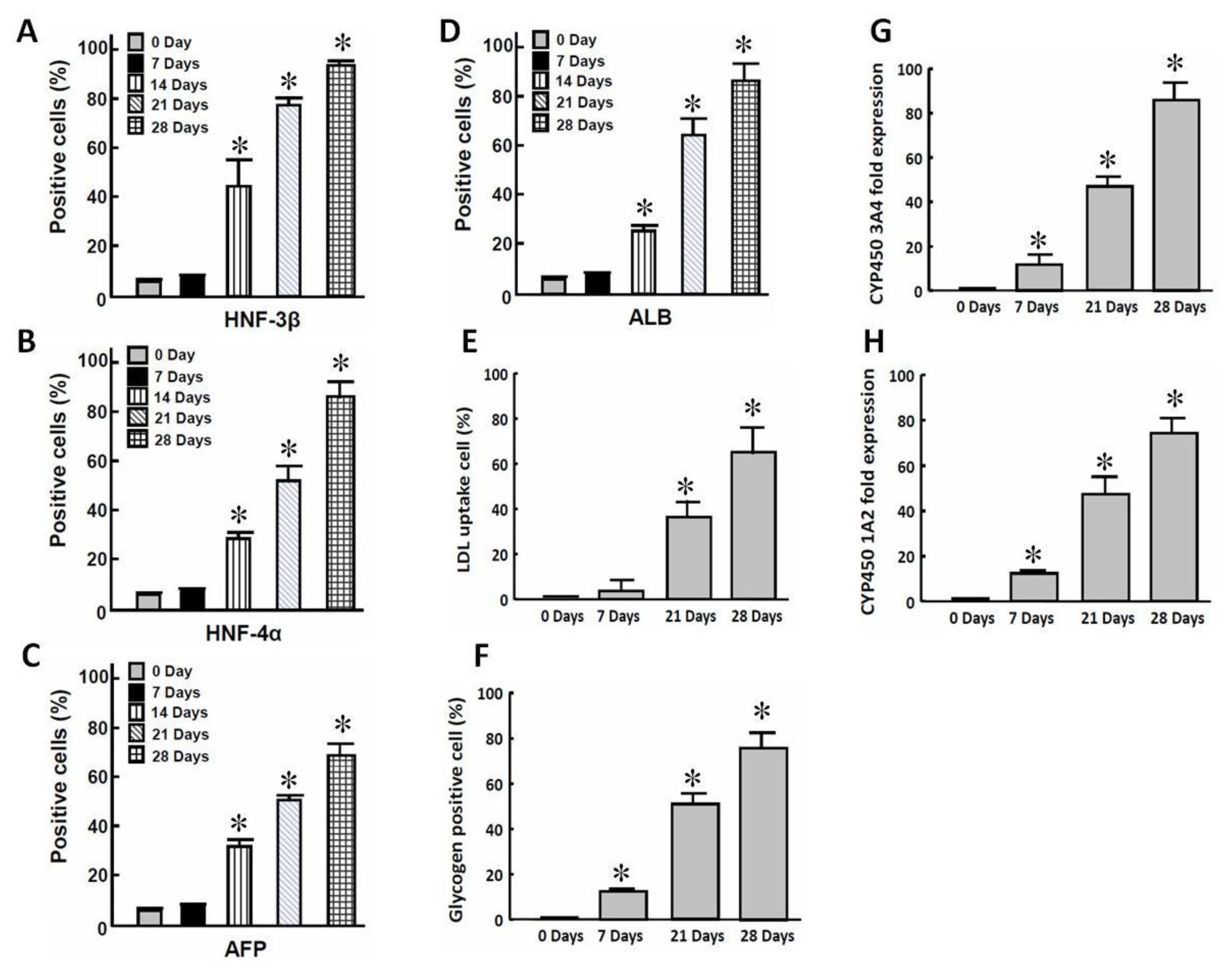
2.1. In Vitro Differentiation of iPSCs into iPSC-Heps
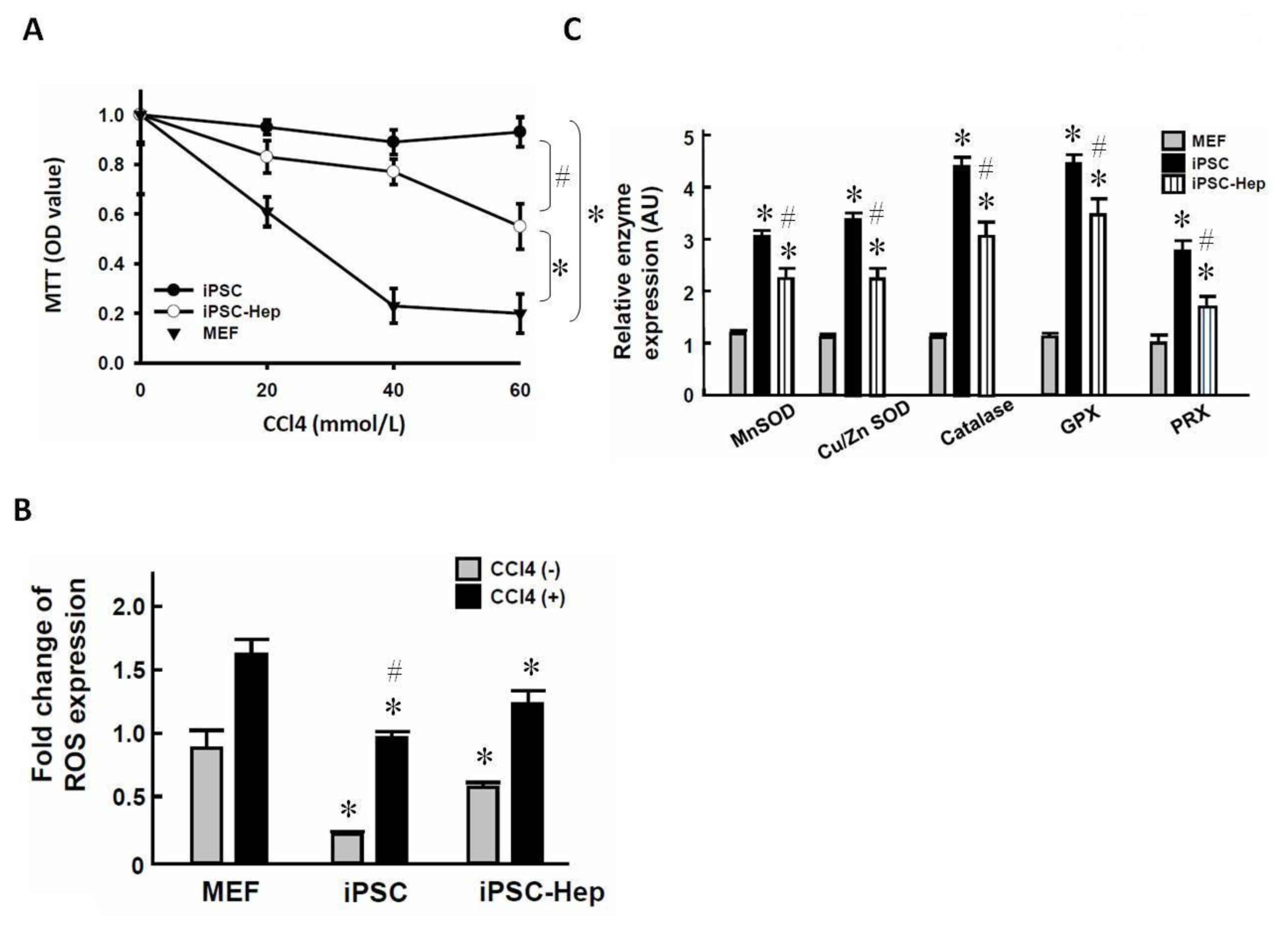
2.2. 3-Genes iPSCs Possess an Antioxidant System Against CCl4-Induced Cell Death in Vitro
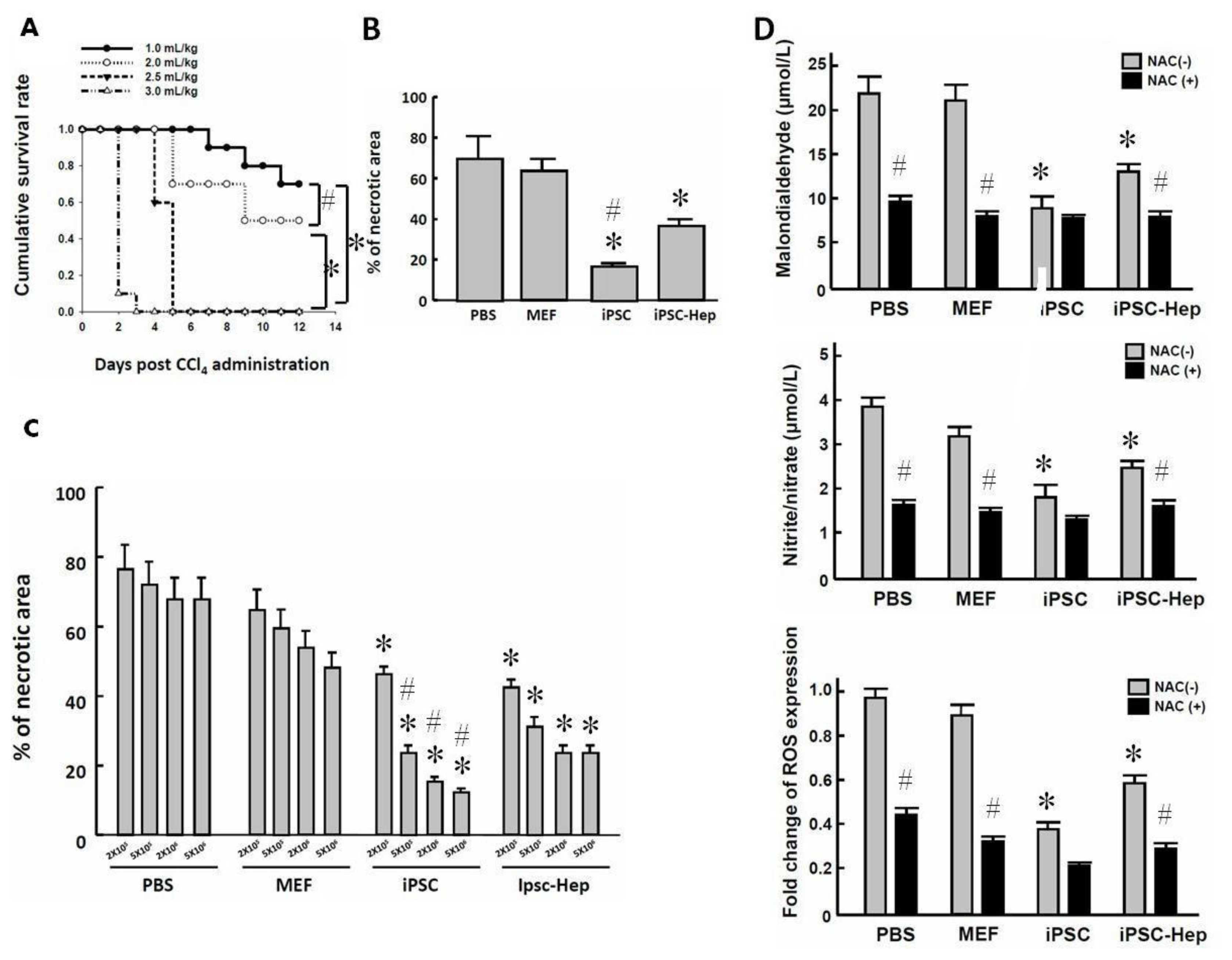
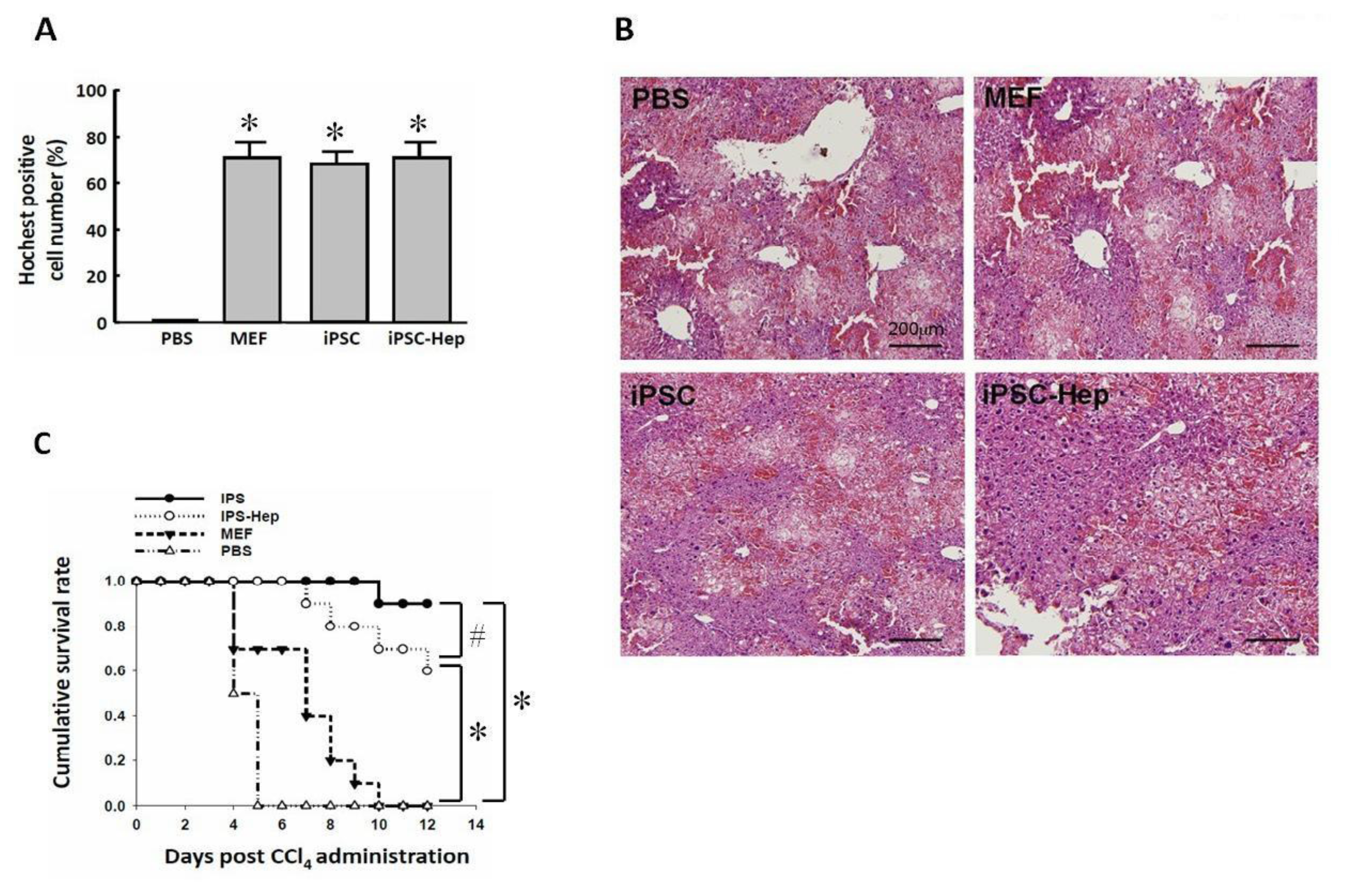
2.3. Intraperitoneal Transplantation of 3-Genes iPSCs Reduced Hepatic Necrotic Area and Oxidative Stress
2.4. 3-Genes iPSCs and 3-Genes iPSC-Heps Rescued the Survival of CCl4-Induced AHF
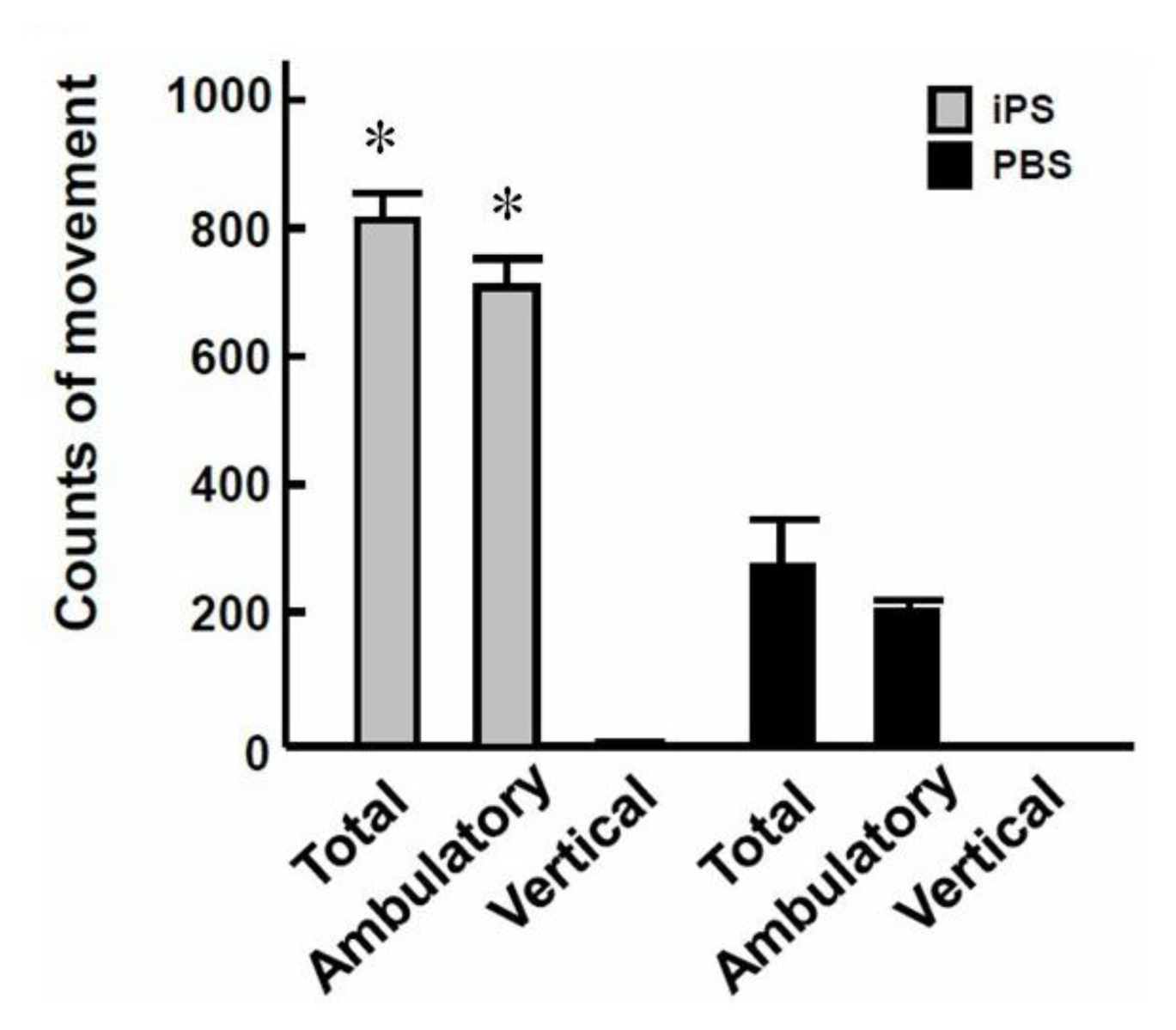
2.5. 3-Genes iPSCs Improved CCl4-Induced Hepatic Encephalopathy and Reduced Tumorigenesis
2.6. Discussion
3. Materials and Methods
3.1. 3-Genes iPSC Culture and in Vitro Hepatic Differentiation
3.2. Cellular Uptake Assay of Low-Density Lipoprotein (LDL)
3.3. Periodic Acid-Schiff (PAS) Stain for Glycogen
3.4. ELISA Analysis
3.5. Real-Time Reverse Transcription-Polymerase Chain Reaction
3.6. Immunofluorescence Staining
3.7. Cell Viability Assay
3.8. Cell-Labeling Protocol
3.9. Animal Model of Liver Injury
3.10. Histological Quantification of Liver Injury
3.11. Liver Functional Tests
3.12. Determination of Intracellular Reactive Oxygen Species (ROS) Production
3.13. Malondialdehyde Assay
3.14. Nitrate/Nitrite Concentration
3.15. Measurement of Motor Activity
3.16. Statistical Analysis
4. Conclusions
Acknowledgments
References
- Tunon, M.J.; Alvarez, M.; Culebras, J.M.; Gonzalez-Gallego, J. An overview of animal models for investigating the pathogenesis and therapeutic strategies in acute hepatic failure. World J. Gastroenterol 2009, 15, 3086–3098. [Google Scholar]
- Fiegel, H.C.; Kaufmann, P.M.; Bruns, H.; Kluth, D.; Horch, R.E.; Vacanti, J.P.; Kneser, U. Hepatic tissue engineering: From transplantation to customized cell-based liver directed therapies from the laboratory. J. Cell. Mol. Med 2008, 12, 56–66. [Google Scholar]
- Heo, J.; Factor, V.M.; Uren, T.; Takahama, Y.; Lee, J.S.; Major, M.; Feinstone, S.M.; Thorgeirsson, S.S. Hepatic precursors derived from murine embryonic stem cells contribute to regeneration of injured liver. Hepatology 2006, 44, 1478–1486. [Google Scholar]
- Takahashi, K.; Tanabe, K.; Ohnuki, M.; Narita, M.; Ichisaka, T.; Tomoda, K.; Yamanaka, S. Induction of pluripotent stem cells from adult human fibroblasts by defined factors. Cell 2007, 131, 861–872. [Google Scholar]
- Okita, K.; Ichisaka, T.; Yamanaka, S. Generation of germline-competent induced pluripotent stem cells. Nature 2007, 448, 313–317. [Google Scholar]
- Yu, J.; Vodyanik, M.A.; Smuga-Otto, K.; Antosiewicz-Bourget, J.; Frane, J.L.; Tian, S.; Nie, J.; Jonsdottir, G.A.; Ruotti, V.; Stewart, R.; et al. Induced pluripotent stem cell lines derived from human somatic cells. Science 2007, 318, 1917–1920. [Google Scholar]
- Si-Tayeb, K.; Noto, F.K.; Nagaoka, M.; Li, J.; Battle, M.A.; Duris, C.; North, P.E.; Dalton, S.; Duncan, S.A. Highly efficient generation of human hepatocyte-like cells from induced pluripotent stem cells. Hepatology 2010, 51, 297–305. [Google Scholar]
- Sancho-Bru, P.; Roelandt, P.; Narain, N.; Pauwelyn, K.; Notelaers, T.; Shimizu, T.; Ott, M.; Verfaillie, C. Directed differentiation of murine-induced pluripotent stem cells to functional hepatocyte-like cells. J. Hepatol 2010, 54, 98–107. [Google Scholar]
- Rashid, S.T.; Corbineau, S.; Hannan, N.; Marciniak, S.J.; Miranda, E.; Alexander, G.; Huang-Doran, I.; Griffin, J.; Ahrlund-Richter, L.; Skepper, J.; et al. Modeling inherited metabolic disorders of the liver using human induced pluripotent stem cells. J. Clin. Invest 2010, 120, 3127–3136. [Google Scholar]
- Seminatore, C.; Polentes, J.; Ellman, D.; Kozubenko, N.; Itier, V.; Tine, S.; Tritschler, L.; Brenot, M.; Guidou, E.; Blondeau, J.; et al. The postischemic environment differentially impacts teratoma or tumor formation after transplantation of human embryonic stem cell-derived neural progenitors. Stroke 2010, 41, 153–159. [Google Scholar]
- Chen, S.J.; Chang, C.M.; Tsai, S.K.; Chang, Y.L.; Chou, S.J.; Huang, S.S.; Tai, L.K.; Chen, Y.C.; Ku, H.H.; Li, H.Y.; et al. Functional improvement of focal cerebral ischemia injury by subdural transplantation of induced pluripotent stem cells with fibrin glue. Stem Cells Dev 2010, 19, 1757–1767. [Google Scholar]
- Li, H.Y.; Chien, Y.; Chen, Y.J.; Chen, S.F.; Chang, Y.L.; Chiang, C.H.; Jeng, S.Y.; Chang, C.M.; Wang, M.L.; Chen, L.K.; et al. Reprogramming induced pluripotent stem cells in the absence of c-myc for differentiation into hepatocyte-like cells. Biomaterials 2011, 32, 5994–6005. [Google Scholar]
- Reinke, L.A.; Lai, E.K.; McCay, P.B. Ethanol feeding stimulates trichloromethyl radical formation from carbon tetrachloride in liver. Xenobiotica 1988, 18, 1311–1318. [Google Scholar]
- Armstrong, L.; Tilgner, K.; Saretzki, G.; Atkinson, S.P.; Stojkovic, M.; Moreno, R.; Przyborski, S.; Lako, M. Human induced pluripotent stem cell lines show stress defense mechanisms and mitochondrial regulation similar to those of human embryonic stem cells. Stem Cells 2010, 28, 661–673. [Google Scholar]
- Kakinuma, S.; Nakauchi, H.; Watanabe, M. Hepatic stem/progenitor cells and stem-cell transplantation for the treatment of liver disease. J. Gastroenterol 2009, 44, 167–172. [Google Scholar]
- Yoshimi, A.; Nannya, Y.; Ueda, K.; Asano, D.; Yamamoto, G.; Kumano, K.; Hangaishi, A.; Matsui, Y.; Takahashi, T.; Imai, Y.; et al. Successful hematopoietic stem cell transplantation from an hla-identical sibling in a patient with aplastic anemia after hla-haploidentical living-related liver transplantation for fulminant hepatitis. Biol. Blood Marrow Transplant 2009, 15, 389–390. [Google Scholar]
- Navarro-Alvarez, N.; Soto-Gutierrez, A.; Kobayashi, N. Stem cell research and therapy for liver disease. Curr. Stem Cell Res. Ther 2009, 4, 141–146. [Google Scholar]
- Nakagawa, M.; Koyanagi, M.; Tanabe, K.; Takahashi, K.; Ichisaka, T.; Aoi, T.; Okita, K.; Mochiduki, Y.; Takizawa, N.; Yamanaka, S. Generation of induced pluripotent stem cells without myc from mouse and human fibroblasts. Nat. Biotechnol 2008, 26, 101–106. [Google Scholar]
- Lee, J.W.; Shin, K.D.; Lee, M.; Kim, E.J.; Han, S.S.; Han, M.Y.; Ha, H.; Jeong, T.C.; Koh, W.S. Role of metabolism by flavin-containing monooxygenase in thioacetamide-induced immunosuppression. Toxicol. Lett 2003, 136, 163–172. [Google Scholar]
- Okuyama, H.; Shimahara, Y.; Nakamura, H.; Araya, S.; Kawada, N.; Yamaoka, Y.; Yodoi, J. Thioredoxin prevents thioacetamide-induced acute hepatitis. Comp. Hepatol 2004, 3, S1–S6. [Google Scholar]
- Bruck, R.; Aeed, H.; Avni, Y.; Shirin, H.; Matas, Z.; Shahmurov, M.; Avinoach, I.; Zozulya, G.; Weizman, N.; Hochman, A. Melatonin inhibits nuclear factor kappa b activation and oxidative stress and protects against thioacetamide induced liver damage in rats. J. Hepatol 2004, 40, 86–93. [Google Scholar]
- Abul, H.; Mathew, T.C.; Dashti, H.M.; Al-Bader, A. Level of superoxide dismutase, glutathione peroxidase and uric acid in thioacetamide-induced cirrhotic rats. Anat. Histol. Embryol 2002, 31, 66–71. [Google Scholar]
- Lu, S.C.; Huang, Z.Z.; Yang, H.; Tsukamoto, H. Effect of thioacetamide on the hepatic expression of gamma-glutamylcysteine synthetase subunits in the rat. Toxicol. Appl. Pharmacol 1999, 159, 161–168. [Google Scholar]
- Rahman, T.M.; Hodgson, H.J. The effects of early and late administration of inhibitors of inducible nitric oxide synthase in a thioacetamide-induced model of acute hepatic failure in the rat. J. Hepatol 2003, 38, 583–590. [Google Scholar]
- Moriya, K.; Yoshikawa, M.; Saito, K.; Ouji, Y.; Nishiofuku, M.; Hayashi, N.; Ishizaka, S.; Fukui, H. Embryonic stem cells develop into hepatocytes after intrasplenic transplantation in ccl4-treated mice. World J. Gastroenterol 2007, 13, 866–873. [Google Scholar]
- Kuo, T.K.; Hung, S.P.; Chuang, C.H.; Chen, C.T.; Shih, Y.R.; Fang, S.C.; Yang, V.W.; Lee, O.K. Stem cell therapy for liver disease: Parameters governing the success of using bone marrow mesenchymal stem cells. Gastroenterology 2008, 134. [Google Scholar]
- Fong, E.L.; Chan, C.K.; Goodman, S.B. Stem cell homing in musculoskeletal injury. Biomaterials 2011, 32, 395–409. [Google Scholar]
- Chen, Y.; Xiang, L.X.; Shao, J.Z.; Pan, R.L.; Wang, Y.X.; Dong, X.J.; Zhang, G.R. Recruitment of endogenous bone marrow mesenchymal stem cells towards injured liver. J. Cell. Mol. Med 2010, 14, 1494–1508. [Google Scholar]
- Lukas, G.; Brindle, S.D.; Greengard, P. The route of absorption of intraperitoneally administered compounds. J. Pharmacol. Exp. Ther 1971, 178, 562–564. [Google Scholar]
- Cash, W.J.; McConville, P.; McDermott, E.; McCormick, P.A.; Callender, M.E.; McDougall, N.I. Current concepts in the assessment and treatment of hepatic encephalopathy. QJM 2010, 103, 9–16. [Google Scholar]
- Kao, C.L.; Tai, L.K.; Chiou, S.H.; Chen, Y.J.; Lee, K.H.; Chou, S.J.; Chang, Y.L.; Chang, C.M.; Chen, S.J.; Ku, H.H.; et al. Resveratrol promotes osteogenic differentiation and protects against dexamethasone damage in murine induced pluripotent stem cells. Stem Cells Dev 2010, 19, 247–258. [Google Scholar]
- Song, Z.; Cai, J.; Liu, Y.; Zhao, D.; Yong, J.; Duo, S.; Song, X.; Guo, Y.; Zhao, Y.; Qin, H.; et al. Efficient generation of hepatocyte-like cells from human induced pluripotent stem cells. Cell Res 2009, 19, 1233–1242. [Google Scholar]
- Chen, Y.C.; Hsu, H.S.; Chen, Y.W.; Tsai, T.H.; How, C.K.; Wang, C.Y.; Hung, S.C.; Chang, Y.L.; Tsai, M.L.; Lee, Y.Y.; et al. Oct-4 expression maintained cancer stem-like properties in lung cancer-derived cd133-positive cells. PLoS One 2008, 3, e2637. [Google Scholar]
- Byrne, J.A.; Grieve, D.J.; Bendall, J.K.; Li, J.M.; Gove, C.; Lambeth, J.D.; Cave, A.C.; Shah, A.M. Contrasting roles of nadph oxidase isoforms in pressure-overload versus angiotensin ii-induced cardiac hypertrophy. Circ. Res 2003, 93, 802–805. [Google Scholar]
- Kerksick, C.; Willoughby, D. The antioxidant role of glutathione and n-acetyl-cysteine supplements and exercise-induced oxidative stress. J. Int. Soc. Sports Nutr 2005, 2, 38–44. [Google Scholar]
- Ribeiro, J.; Nordlinger, B.; Ballet, F.; Cynober, L.; Coudray-Lucas, C.; Baudrimont, M.; Legendre, C.; Delelo, R.; Panis, Y. Intrasplenic hepatocellular transplantation corrects hepatic encephalopathy in portacaval-shunted rats. Hepatology 1992, 15, 12–18. [Google Scholar]
- Chu, C.J.; Lee, F.Y.; Wang, S.S.; Chang, F.Y.; Lin, H.C.; Wu, S.L.; Chan, C.C.; Tsai, Y.T.; Lee, S.D. Establishment of an animal model of hepatic encephalopathy. Zhonghua Yi Xue Za Zhi (Taipei) 2000, 63, 263–269. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Alanine aminotransferase (ALT; IU/L) | Aspartate aminotransferase (AST; IU/L) | Total bilirubin (TBIL; mg/dL) | |
|---|---|---|---|
| BALB/c nude mice | 11.1 ± 3.5 | 55.2 ± 10.8 | 0.45 ± 0.4 |
| Recipient day 1 Post CCl4 | |||
| PBS | 10125.5 ± 680.5 | 6610.4 ± 800.5 | 4.40 ± 0.42 |
| MEFs | 9912.2 ± 620.0 | 5618.9 ± 428.2 | 4.24 ± 0.12 |
| iPSC-Heps | 5084.5 ± 432.8 * | 2359.5 ± 307.1 * | 3.24 ± 0.22 * |
| iPSCs | 2339.3 ± 349.8 *,# | 1369.3 ± 210.0 *,# | 2.28 ± 0.20 *,# |
| Recipient day 2 Post CCl4 | |||
| PBS | 3135.1 ± 451.4 | 2660.6 ± 286.5 | 3.11 ± 0.35 |
| MEFs | 2679.8 ± 424.7 | 2241.2 ± 249.4 | 2.22 ± 0.21 |
| iPSC-Heps | 1807.6 ± 305.5 * | 1527.5 ± 179.8 * | 1.90 ± 0.28 * |
| iPSCs | 865.6 ± 171.2 *,# | 767.5 ± 227.4 *,# | 1.29 ± 0.15 *,# |
| Recipient day 3 Post CCl4 | |||
| PBS | 598.0 ± 60.2 | 412.1 ± 28.5 | 2.18 ± 0.17 |
| MEFs | 459.2 ± 56.1 | 362.8 ± 8.5 | 1.32 ± 0.12 |
| iPSC-Heps | 139.1 ± 10.9 * | 104.5 ± 8.8 * | 1.22 ± 0.18 * |
| iPSCs | 95.6 ± 5.5 *,# | 60.3 ± 6.7 *,# | 1.13 ± 0.17 *,# |
| Group tumor formation | 1 month | 2 months | 3 months | 4 months |
|---|---|---|---|---|
| 4-genes iPSCs | 0/6 | 1/6 | 2/6 | 3/6 |
| 3-genes iPSCs | 0/6 | 0/6 | 0/6 | 0/6 |
| 4-genes iPSC-Heps | 0/6 | 0/6 | 0/6 | 0/6 |
© 2012 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Chang, H.-M.; Liao, Y.-W.; Chiang, C.-H.; Chen, Y.-J.; Lai, Y.-H.; Chang, Y.-L.; Chen, H.-L.; Jeng, S.-Y.; Hsieh, J.-H.; Peng, C.-H.; et al. Improvement of Carbon Tetrachloride-Induced Acute Hepatic Failure by Transplantation of Induced Pluripotent Stem Cells without Reprogramming Factor c-Myc. Int. J. Mol. Sci. 2012, 13, 3598-3617. https://doi.org/10.3390/ijms13033598
Chang H-M, Liao Y-W, Chiang C-H, Chen Y-J, Lai Y-H, Chang Y-L, Chen H-L, Jeng S-Y, Hsieh J-H, Peng C-H, et al. Improvement of Carbon Tetrachloride-Induced Acute Hepatic Failure by Transplantation of Induced Pluripotent Stem Cells without Reprogramming Factor c-Myc. International Journal of Molecular Sciences. 2012; 13(3):3598-3617. https://doi.org/10.3390/ijms13033598
Chicago/Turabian StyleChang, Hua-Ming, Yi-Wen Liao, Chih-Hung Chiang, Yi-Jen Chen, Ying-Hsiu Lai, Yuh-Lih Chang, Hen-Li Chen, Shaw-Yeu Jeng, Jung-Hung Hsieh, Chi-Hsien Peng, and et al. 2012. "Improvement of Carbon Tetrachloride-Induced Acute Hepatic Failure by Transplantation of Induced Pluripotent Stem Cells without Reprogramming Factor c-Myc" International Journal of Molecular Sciences 13, no. 3: 3598-3617. https://doi.org/10.3390/ijms13033598
APA StyleChang, H. -M., Liao, Y. -W., Chiang, C. -H., Chen, Y. -J., Lai, Y. -H., Chang, Y. -L., Chen, H. -L., Jeng, S. -Y., Hsieh, J. -H., Peng, C. -H., Li, H. -Y., Chien, Y., Chen, S. -Y., Chen, L. -K., & Huo, T. -I. (2012). Improvement of Carbon Tetrachloride-Induced Acute Hepatic Failure by Transplantation of Induced Pluripotent Stem Cells without Reprogramming Factor c-Myc. International Journal of Molecular Sciences, 13(3), 3598-3617. https://doi.org/10.3390/ijms13033598