Effects of Inflorescence Stem Structure and Cell Wall Components on the Mechanical Strength of Inflorescence Stem in Herbaceous Peony

Abstract
:1. Introduction
2. Results
2.1. Mechanical Strength and Morphological Indices
2.2. Microstructure
2.3. Cellulose and Lignin Contents
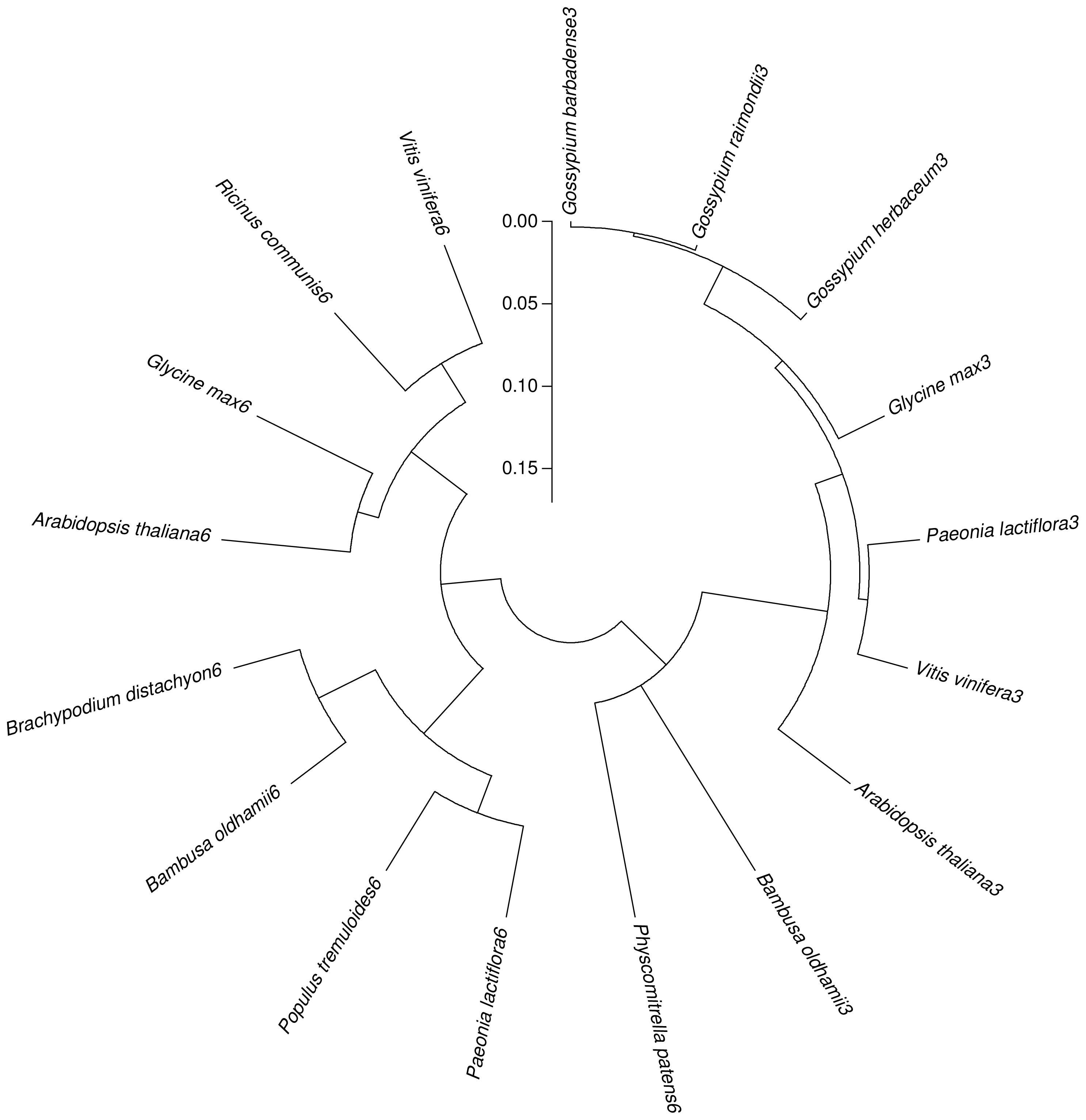
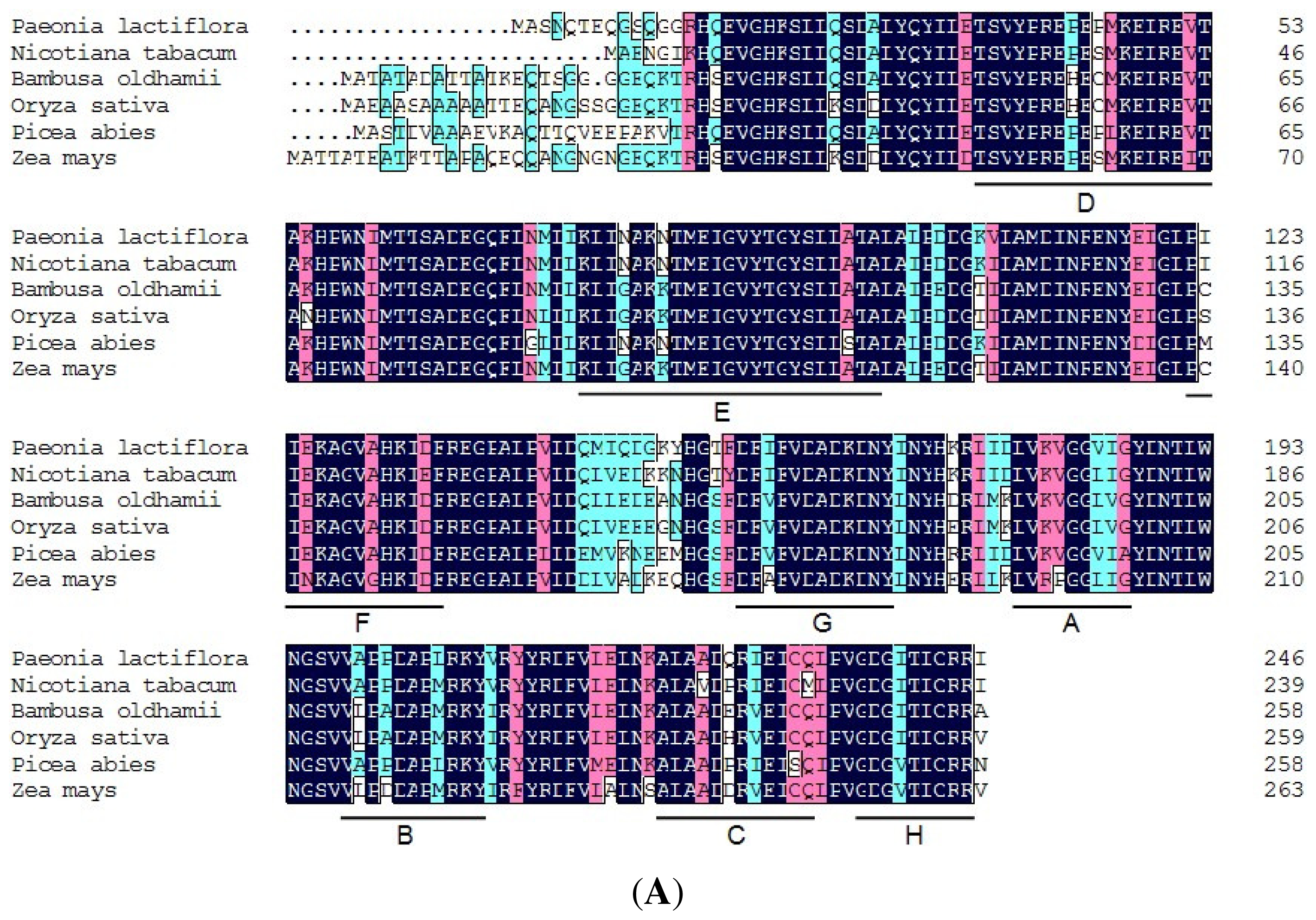
2.4. Isolation and Sequence Analysis
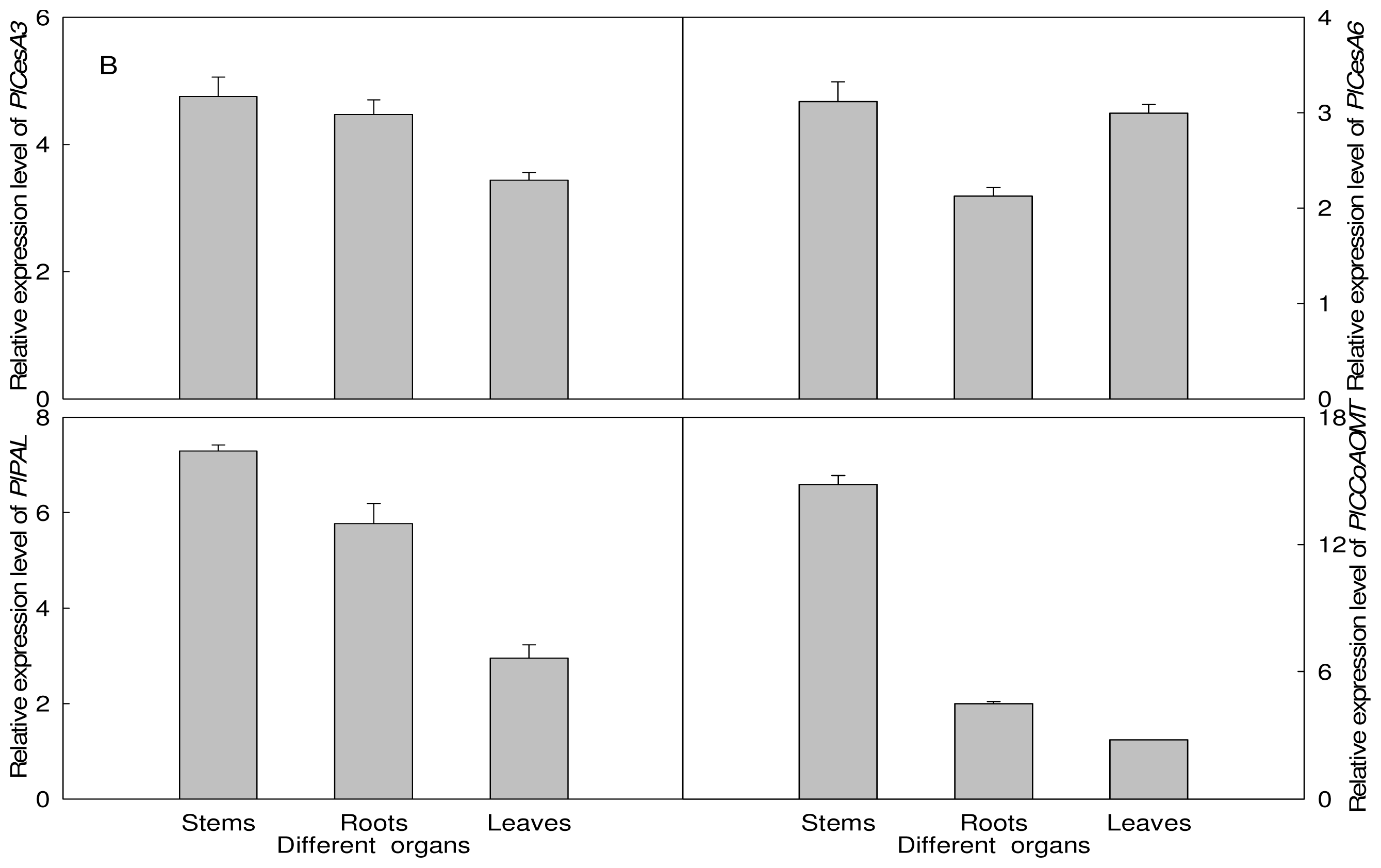
2.5. Expression Analysis
3. Discussion
4. Experimental Section
4.1. Plant Materials
4.2. Morphological Indices and Mechanical Strength Determination
4.3. Microstructure Observation
4.4. Cell Wall Materials Fractionation, Cellulose and Lignin Contents Determination
4.5. RNA Extraction and Primers Design
4.6. Isolation of the Full-length cDNA Sequence
4.7. Gene Expression Analysis
4.8. Sequence and Statistical Analysis
Acknowledgments
References
- Qing, K.J. The Origin and History. In Illustration of One Hundred Ornamental Flowers Bonsai—The Herbaceous Peony; Qing, K.J., Ed.; China Forestry Publishing House: Beijing, China, 2004; pp. 12–19. [Google Scholar]
- Stevens, S.; Stevens, A.B.; Gast, K.L.B.; O’Mara, J.A.; Tisserat, N.A.; Bauernfeind, R. Commercial specialty cut flower production, peonies. Cooperative Extension Service; Kansas State University: Manhattan, Kansas, 1993. Available online: http://www.oznet.ksu.edu/library/hort2/mf1083.pdf accessed on 1 March 2012.
- Yu, X.; Guo, P.; Lu, G.; Zhang, Q. Optimum harvesting time of herbaceous peony buds for cutting flowers. J. For. Res 2011, 22, 137–140. [Google Scholar]
- Wang, J.; Zhu, J.; Lin, Q.; Li, X.; Teng, N.; Li, Z.; Li, B.; Zhang, A.; Lin, J. Effects of stem structure and cell wall components on bending strength in wheat. Chin. Sci. Bull 2006, 51, 815–823. [Google Scholar]
- Li, X.; Yang, Y.; Yao, J.; Chen, G.; Li, X.; Zhang, Q.; Wu, C. FLEXIBLE CULM 1 encoding a cinnamyl-alcohol dehydrogenase controls culm mechanical strength in rice. Plant Mol. Biol 2009, 69, 685–697. [Google Scholar]
- Li, Y.; Qian, Q.; Zhou, Y.; Yan, M.; Sun, L.; Zhang, M.; Fu, Z.; Wang, Y.; Han, B.; Pang, X.; et al. BRITTLE CULM1, which encodes a COBRA-like protein, affects the mechanical properties of rice plants. Plant Cell 2003, 15, 2020–2031. [Google Scholar]
- Gilbert, H.J. The biochemistry and structural biology of plant cell wall deconstruction. Plant Physiol 2010, 153, 444–455. [Google Scholar]
- Taylor, N.G.; Scheible, W.; Cutler, S.; Somerville, C.R.; Turner, S.R. The irregular xylem3 locus of Arabidopsis encodes a cellulose synthase required for secondary cell wall synthesis. Plant Cell 1999, 11, 769–779. [Google Scholar]
- Hirano, K.; Kotake, T.; Kamihara, K.; Tsuna, K.; Aohara, T; Kaneko, Y.; Takatsuji, H.; Tsumuraya, Y.; Kawasaki, S. Rice BRITTLE CULM 3 (BC3) encodes a classical dynamin OsDRP2B essential for proper secondary cell wall synthesis. Planta 2010, 232, 95–108. [Google Scholar]
- Aohara, T.; Kotake, T.; Kaneko, Y.; Takatsuji, H.; Tsumuraya, Y.; Kawasaki, S. Rice BRITTLE CULM 5 (BRITTLE NODE) is involved in secondary cell wall formation in the sclerenchyma tissue of nodes. Plant Cell Physiol 2009, 50, 1886–1897. [Google Scholar]
- Yan, C.; Yan, S.; Zeng, X.; Zhang, Z.; Gu, M. Fine mapping and isolation of Bc7(t), allelic to OsCesA4. J. Genet. Genomics 2007, 34, 1019–1027. [Google Scholar]
- Zhou, Y.; Li, S.; Qian, Q.; Zeng, D.; Zhang, M.; Guo, L.; Liu, X.; Zhang, B.; Deng, L.; Liu, X.; et al. BC10, a DUF266-containing and Golgi-located type II membrane protein, is required for cell-wall biosynthesis in rice (Oryza sativa L.). Plant J 2009, 57, 446–462. [Google Scholar]
- Kokubo, A.; Sakurai, N.; Kuraishi, S.; Takeda, K. Culm brittleness of barley (Hordeum vulgare L.) mutants is caused by smaller number of cellulose molecules in cell wall. Plant Physiol 1991, 97, 509–514. [Google Scholar]
- Burton, R.A.; Ma, G.; Baumann, U.; Harvey, A.J.; Shirley, N.J.; Taylor, J.; Pettolino, F.; Bacic, A.; Beatty, M.; Simmons, C.R.; et al. A customized gene expression microarray reveals that the brittle stem phenotype fs2 of barley is attributable to a retroelement in the HvCesA4 cellulose synthase gene. Plant Physiol 2010, 153, 1716–1728. [Google Scholar]
- Brown, R.M.J.; Montezinos, D. Cellulose microfibrils: Visualization of biosynthetic and orienting complexes in association with the plasma membrane. Proc. Natl. Acad. Sci. USA 1976, 73, 143–147. [Google Scholar]
- Pear, J.R.; Kawagoe, Y.; Schreckengost, W.E.; Delmer, D.P.; Stalker, D.M. Higher plants contain homologs of the bacterial celA genes encoding the catalytic subunit of cellulose synthase. Proc. Natl. Acad. Sci. USA 1996, 93, 12637–12642. [Google Scholar]
- Richmond, T.A.; Somerville, C.R. The cellulose synthase superfamily. Plant Physiol 2000, 124, 495–498. [Google Scholar]
- Hazen, S.P.; Scott-Craig, J.S.; Walton, J.D. Cellulose synthase-like genes of rice. Plant Physiol 2002, 12, 336–340. [Google Scholar]
- Burton, R.A.; Shirley, N.J.; King, B.J.; Harvey, A.J.; Fincher, G.B. The CesA gene family of barley. Quantitative analysis of transcripts reveals two groups of co-expressed genes. Plant Physiol 2004, 134, 224–236. [Google Scholar]
- Wise, H.Z.; Saxena, I.M.; Brown, R.M.J. Isolation and characterization of the cellulose synthase genes PpCesA6 and PpCesA7 in Physcomitrella patens. Cellulose 2011, 18, 371–384. [Google Scholar]
- Kokubo, A.; Kuraishi, S.; Sakurai, N. Culm strength of barley: Correlation among maximum bending stress, cell wall dimensions, and cellulose content. Plant Physiol 1989, 91, 876–882. [Google Scholar]
- Howles, P.A.; Sewalt, V.J.H.; Paiva, N.L.; Elkind, Y.; Bate, N.J.; Lamb, C.; Dixon, R.A. Overexpression of l-phenylalanine ammonia-lyase in transgenic tobacco plants reveals control points for flux into phenylpropanoid biosynthesis. Plant Physiol 1996, 112, 1617–1624. [Google Scholar]
- Zhong, R.; Morrison, W.H.; Negrel, J.; Ye, Z.H. Dual methylation pathways in lignin biosynthesis. Plant Cell 1998, 10, 2033–2045. [Google Scholar]
- Wagner, A.; Tobimatsu, Y.; Phillips, L.; Heather, F.; Kirk, T.; Lloyd, D.; Lana, P.; John, R. CCoAOMT suppression modifies lignin composition in Pinus radiata. Plant J 2011, 67, 119–129. [Google Scholar]
- Qi, J.; Yu, S.; Zhang, F.; Shen, X.; Zhao, X.; Yu, Y.; Zhang, D. Reference gene selection for real-time quantitative polymerase chain reaction of mRNA transcript levels in Chinese cabbage (Brassica rapa L. ssp. pekinensis). Plant Mol. Biol. Report 2010, 28, 597–604. [Google Scholar]
- Yang, H.; Yang, R.; Li, Y.; Jiang, Z.; Zheng, J. Relationship between culm traits and lodging resistance of rice cultivars. Fujian J. Agric. Sci 2000, 15, 1–7. [Google Scholar]
- Shen, G.; Shi, Y.; Huang, Y.; Shi, Y.; Wang, W.; Zhang, C.; Chen, D. Study on rice lodging resistance character and correlation between the culm traits and lodging resistance traits. Chin. Agric. Sci. Bull 2007, 23, 58–62. [Google Scholar]
- Mo, Y.S.; Yang, Q.Q.; Liu, P.Q.; He, L.S.; Wang, A.Q.; Fan, Y.S.; Zhao, X.Y. Relationship of tall and tough rice resistance and stalk structure. Crops 2007, 2, 40–42. [Google Scholar]
- Su, Y.; Yan, C.; Wang, X.; Zhao, G. Analysis of cell wall components and ultrastructure for brittleness mutation rice. Acta Agrestia Sin 2008, 16, 594–599. [Google Scholar]
- Wei, C.; Xie, P.; Zhou, W.; Chen, Y.; Yan, C. Anatomical structure and chemical features of leaf in brittle mutant of rice. Acta Agron. Sin 2008, 34, 1417–1423. [Google Scholar]
- Xu, S.X.; Xu, X.B. Morphology and Anatomy of Rice; Agriculture Press: Beijing, China, 1984; pp. 12–30. [Google Scholar]
- Baucher, M.; Monties, B.; van Montagu, M.; Boerjan, W. Biosynthesis and genetic engineer in lignin. Crit. Rev. Plant Sci 1998, 17, 125–197. [Google Scholar]
- Zhang, J.; Chen, J.; Tang, Z.; Wang, R. Study on the physico-chemical properties of stem as related to lodging in rape. J. Southwest Agric. Univ. (Nat. Sci. ) 2006, 28, 763–765. [Google Scholar]
- Sindhu, A.; Langewisch, T.; Olek, A.; Multani, D.S.; McCann, M.C.; Vermerris, W.; Carpita, N.C.; Johal, G. Maize brittle stalk2 encodes a COBRA-Like protein expressed in early organ development but required for tissue flexibility at maturity. Plant Physiol 2007, 145, 1444–1459. [Google Scholar]
- Wu, N.; Zhang, T.T.; Zhou, H.P.; Fan, J.H. Impact of inter-planting different rice varieties on the chemical components of rice stem. J. Yunnan Agric. Univ 2008, 23. [Google Scholar]
- Ookawa, T.; Ishihara, K.V. Difference of the cell wall components affecting the ding stress of the culm in relating to the lodging resistance in paddy rice. Jpn. J. Crop Sci 1993, 62, 378–384. [Google Scholar]
- Saxena, I.M.; Lin, F.C.; Brown, R.M.J. Identification of a new gene in an operon for cellulose biosynthesis in Acetobacter xylinum. Plant Mol. Biol 1991, 16, 947–954. [Google Scholar]
- Robert, S.; Mouille, G.; Höfte, H. The mechanism and regulation of cellulose synthesis in primary walls: Lessons from cellulose deficient Arabidopsis mutants. Cellulose 2004, 11, 351–364. [Google Scholar]
- Taylor, N.G.; Gardiner, J.C.; Whiteman, R.; Turner, S.R. Cellulose synthesis in the Arabidopsis secondary cell wall. Cellulose: Molecular and Structural Biology. Cellulose 2004, 11, 329–338. [Google Scholar]
- Somerville, C. Cellulose synthesis in higher plants. Annu. Rev. Cell Dev. Biol 2006, 22, 53–78. [Google Scholar]
- Xie, X.M.; Zhao, Y.H.; Huo, S.; Chen, H. Dynamic changes of enzyme activities related to lignin biosynthesis for elephant grass cultivars. Acta Agrestia Sin 2010, 18, 523–527. [Google Scholar]
- Rose, J.K.; Hadfield, K.A.; Labavitch, J.M.; Bennet, A.B. Temporal sequence of cell wall disassembly in rapidly ripening melon fruit. Plant Physiol 1998, 117, 345–361. [Google Scholar]
- Updegraff, D.M. Semimicro determination of cellulose inbiological materials. Anal. Biochem 1969, 32, 420–424. [Google Scholar]
- Müsel, G.; Schindler, T.; Bergfeld, R.; Ruel, K.; Jacquet, G.; Lapierre, C.; Speth, V.; Schopfer, P. Structure and distribution of lignin in primary and secondary cell walls of maize coleoptiles analyzed by chemical and immunological probes. Planta 1997, 201, 146–159. [Google Scholar]
- Zhao, D.Q.; Zhou, C.H.; Tao, J. Carotenoid accumulation and carotenogenic genes expression during two types of persimmon fruit (Diospyros kaki L.) development. Plant Mol. Biol. Report 2011, 29, 646–654. [Google Scholar]
- NCBI/BLAST-Basic Logic Alignment Search Tool. National Center for Biotechnology Information (NCBI): Bethesda, MD, USA, 2011. Available online: http://blast.ncbi.nlm.nih.gov/Blast.cgi accessed on 1 March 2012.
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol 2011, 28, 2731–2739. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Development Stages | S1 | S2 | S3 | S4 |
|---|---|---|---|---|
| Plant height (cm) | 49.63 ± 0.40 d | 59.34 ± 0.13 c | 64.93 ± 0.09 b | 67.64 ± 0.09 a |
| Diameter of inflorescence stem (cm) | 0.28 ± 0.01 c | 0.47 ± 0.00 b | 0.63 ± 0.00 a | 0.64 ± 0.00 a |
| Fresh weight of inflorescence stem (g) | 0.49 ± 0.08 c | 0.78 ± 0.00 b | 0.86 ± 0.00 a | 0.86 ± 0.01 a |
| Diameter of flower (cm) | 1.19 ± 0.02 d | 2.00 ± 0.01 c | 9.47 ± 0.10 b | 12.61 ± 0.05 a |
| Fresh weight of flower (g) | 0.69 ± 0.03 d | 3.45 ± 0.08 c | 18.65 ± 0.20 b | 25.82 ± 0.04 a |
| Morphological Indices | R (Correlation Coefficient) |
|---|---|
| Plant height (cm) | 0.95 * |
| Diameter of inflorescence stem (cm) | 0.98 * |
| Fresh weight of inflorescence stem (g) | 0.88 |
| Diameter of flower (cm) | 0.96 * |
| Fresh weight of flower (g) | 0.96 * |
| Development Stages | S1 | S2 | S3 | S4 |
|---|---|---|---|---|
| Number of vascular bundle | 14.67 ± 1.53 c | 25.33 ± 1.15 b | 36.67 ± 1.53 a | 38.33 ± 1.53 a |
| Vascular bundle (%) | 22.40 ± 0.75 b | 32.19 ± 0.48 a | 39.09 ± 1.39 a | 41.35 ± 2.33 a |
| Pith (%) | 6.95 ± 1.07 b | 7.82 ± 0.18 b | 12.06 ± 1.26 a | 14.91 ± 2.13 a |
| Primer | Oligonucleotide Sequence (5′-3′) | Application |
|---|---|---|
| CA3-1 | GAGCTGCTATGTGTCTGATGA | 1st of 3′ RACE |
| CA3-2 | GAGTTTGCAAGGAGATGGGT | 2nd of 3′ RACE |
| CA3-3 | CCCCTCCTCTATATCCTCCAGGCTGAA | 5′ RACE |
| CA6-1 | TTGTGAAAGAACGGAGGG | 1st of 3′ RACE |
| CA6-2 | TGCCAAGGCTCAAAAGGT | 2nd of 3′ RACE |
| CA6-3 | TTCTTGTTGGTGGCTTCT | 1st of 5′ RACE |
| CA6-4 | TCATACTCTCTCTTCATTGCCC | 2nd of 5′ RACE |
| CCoAOMT1 | TTGTGAAAGAACGGAGGG | 1st of 3′ RACE |
| CCoAOMT2 | TGCCAAGGCTCAAAAGGT | 2nd of 3′ RACE |
| CCoAOMT3 | CCCATTCCATAGGGTGTTGTCGTAGCC | 5′ RACE |
| ActinF | GCAGTGTTCCCCAGTATT | expression pattern analysis |
| ActinR | TCTTTTCCATGTCATCCC | |
| CA3F | GTTGCCTCTACGCTTATG | |
| CA3R | CACTTCCCCACTCTGATT | |
| CA6F | GGGTTATTGAAGGTTTTAGC | |
| CA6R | TATCAGCAGTGTAGTCGGA | |
| PALF | ACATTCTCGCCACTACCA | |
| PALR | CTTCCGAAATTCCTCCAC | |
| CCoAOMTF | GCGTGAAGTAACAGCAAAAC | |
| CCoAOMTR | AGAGCAGTAGCAAGGAGAGA |
© 2012 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Zhao, D.; Han, C.; Tao, J.; Wang, J.; Hao, Z.; Geng, Q.; Du, B. Effects of Inflorescence Stem Structure and Cell Wall Components on the Mechanical Strength of Inflorescence Stem in Herbaceous Peony. Int. J. Mol. Sci. 2012, 13, 4993-5009. https://doi.org/10.3390/ijms13044993
Zhao D, Han C, Tao J, Wang J, Hao Z, Geng Q, Du B. Effects of Inflorescence Stem Structure and Cell Wall Components on the Mechanical Strength of Inflorescence Stem in Herbaceous Peony. International Journal of Molecular Sciences. 2012; 13(4):4993-5009. https://doi.org/10.3390/ijms13044993
Chicago/Turabian StyleZhao, Daqiu, Chenxia Han, Jun Tao, Jing Wang, Zhaojun Hao, Qingping Geng, and Bei Du. 2012. "Effects of Inflorescence Stem Structure and Cell Wall Components on the Mechanical Strength of Inflorescence Stem in Herbaceous Peony" International Journal of Molecular Sciences 13, no. 4: 4993-5009. https://doi.org/10.3390/ijms13044993
APA StyleZhao, D., Han, C., Tao, J., Wang, J., Hao, Z., Geng, Q., & Du, B. (2012). Effects of Inflorescence Stem Structure and Cell Wall Components on the Mechanical Strength of Inflorescence Stem in Herbaceous Peony. International Journal of Molecular Sciences, 13(4), 4993-5009. https://doi.org/10.3390/ijms13044993