Pu-erh Tea Reduces Nitric Oxide Levels in Rats by Inhibiting Inducible Nitric Oxide Synthase Expression through Toll-Like Receptor 4

Abstract
:1. Introduction
2. Results and Discussion
2.1. Main Contents in Pu-erh Tea
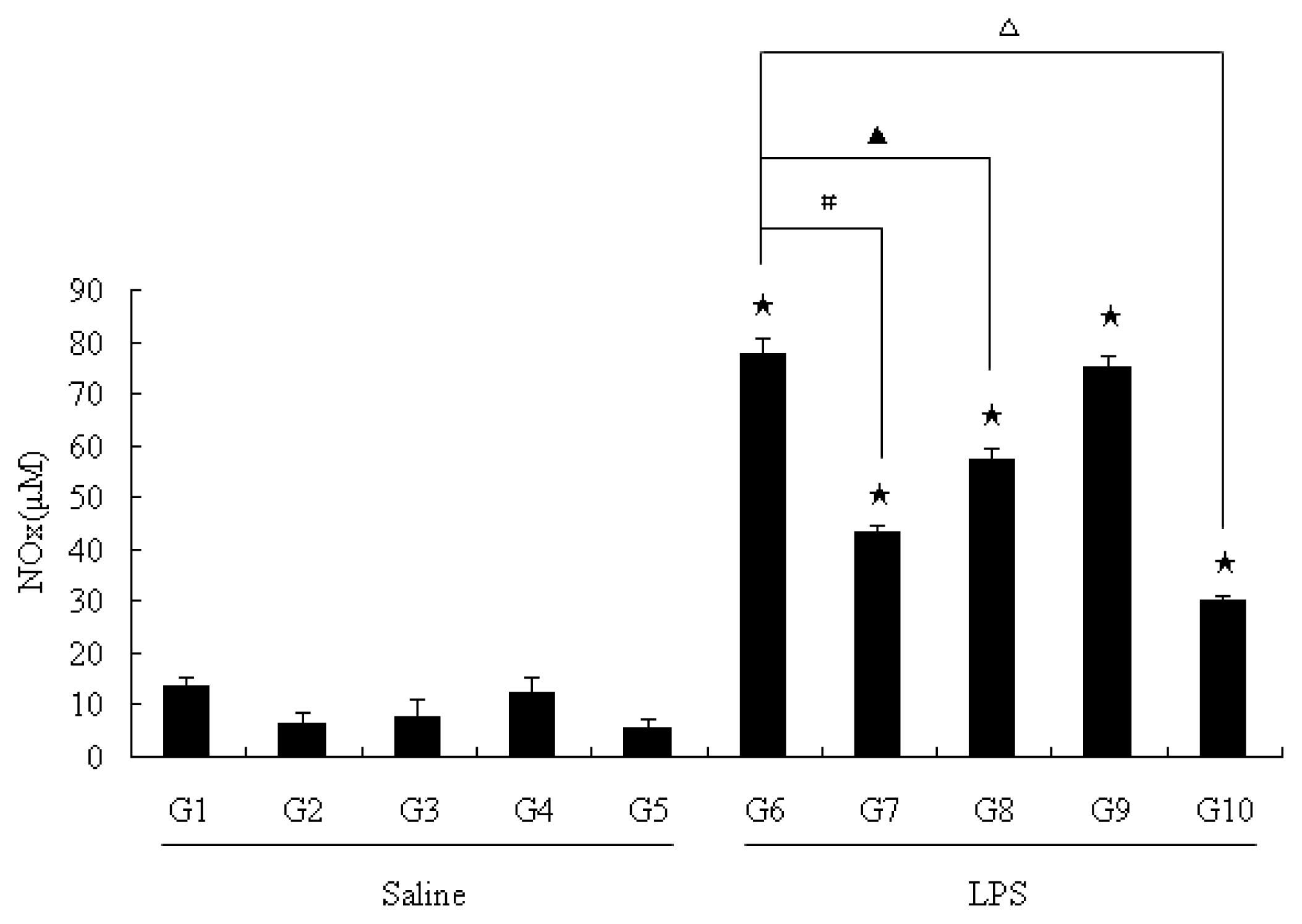
2.2. NO Level in Serum
2.3. NO Level in Liver Homogenates
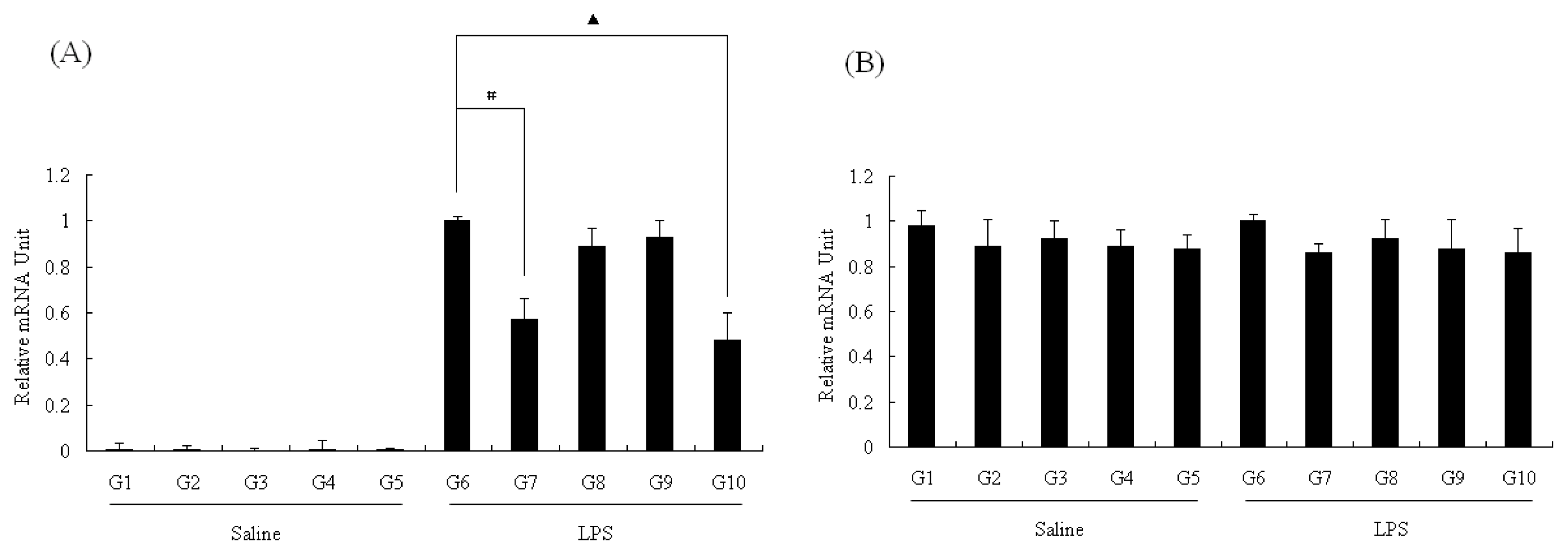
2.4. NOS mRNA Level in Liver
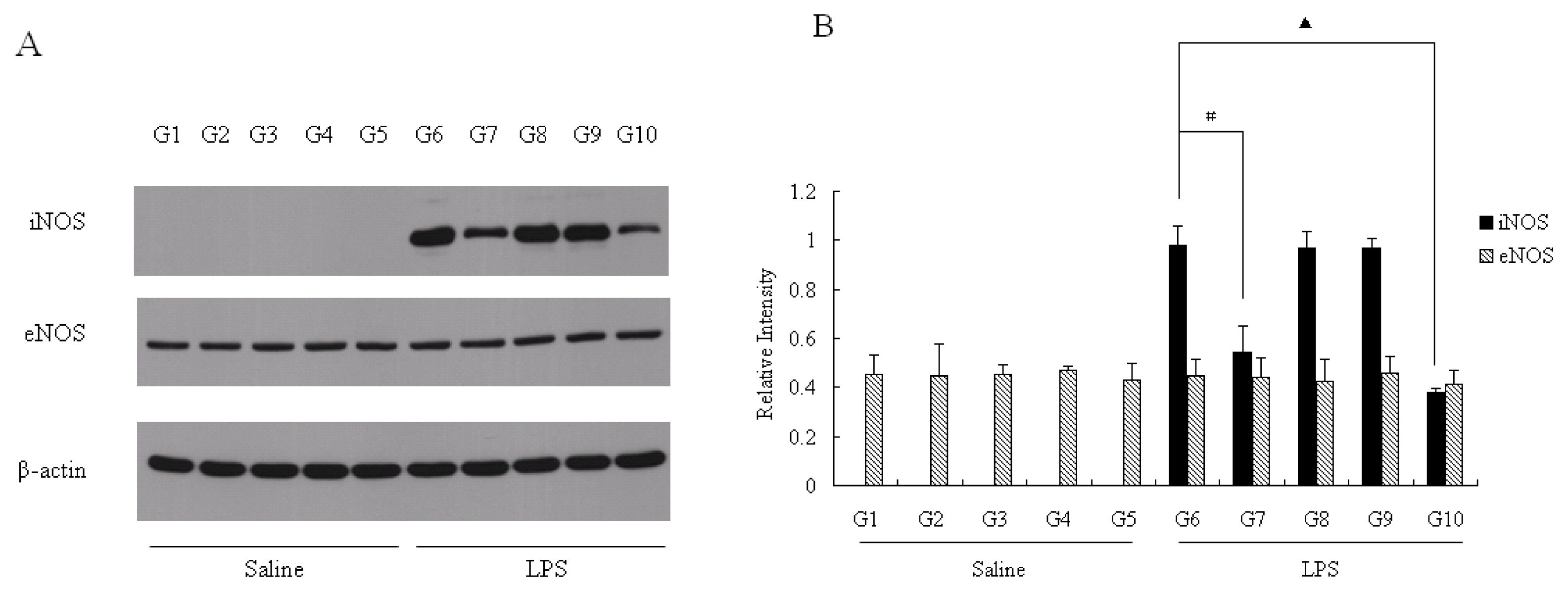
2.5. NOS Protein Expression in Liver
2.6. Expression and Activities of iNOS Expression Signal Pathway
3. Experimental Section
3.1. Materials
3.2. Determination of Polyphenols, Polysaccharides, and Caffeine Content in Concentrated Pu-erh Tea Extracts
3.3. Experimental Animals
- G1 (group1): the group consumed water, which was injected with saline;
- G2 (group2): the group consumed pu-erh tea aqueous extracts (250 mg/kg/day) dissolved in water, which were injected with saline;
- G3 (group3): the group consumed pu-erh tea theabrownins (50 mg/kg/day) dissolved in water, which were injected with saline;
- G4 (group4): the group consumed pu-erh tea caffeine (50 mg/kg/day) dissolved in water, which was injected with saline;
- G5 (group5): the group consumed pu-erh tea polysaccharides (50 mg/kg/day) dissolved in water, which were injected with saline;
- G6 (group6): the group consumed water, which was injected with LPS;
- G7 (group7): the group consumed pu-erh tea aqueous extracts (250 mg/kg/day) dissolved in water, which were injected with LPS;
- G8 (group8): the group consumed pu-erh tea theabrownins (50 mg/kg/day) dissolved in water, which were injected with LPS;
- G9 (group9): the group consumed pu-erh tea caffeine (50 mg/kg/day) dissolved in water, which was injected with LPS;
- G10 (group10): the group consumed pu-erh tea polysaccharides (50 mg/kg/day) dissolved in water, which were injected with LPS.
3.4. NO Assay
3.5. Protein Assay
3.6. RT-PCR
3.7. Western Blot
3.8. Statistical Analysis
4. Conclusions
Acknowledgment
- Conflict of InterestThe authors declare no conflict of interest.
References
- Zhang, Y.Q.; Mao, Z.; Zheng, Y.L.; Han, B.P.; Chen, L.T.; Li, J.; Li, F. Elevation of inducible nitric oxide synthase and cyclooxygenase-2 expression in the mouse brain after chronic nonylphenol exposure. Int. J. Mol. Sci 2008, 9, 1977–1988. [Google Scholar]
- Medvedev, A.E.; Piao, W.; Shoenfelt, J.; Rhee, S.H.; Chen, H.; Basu, S.; Wahl, L.M.; Fenton, M.J.; Vogel, S.N. Role of TLR4 tyrosine phosphorylation in signal transduction and endotoxin tolerance. J. Biol. Chem 2007, 282, 16042–16053. [Google Scholar]
- Chen, T.; Zamora, R.; Zuckerbraun, B.; Billiar, T.R. Role of nitric oxide in liver injury. Curr. Mol. Med 2003, 3, 519–526. [Google Scholar]
- McNaughton, L.; Puttagunta, L.; Martinez-Cuesta, M.A.; Kneteman, N.; Mayers, I.; Moqbel, R.; Hamid, Q.; Radomski, M.W. Distribution of nitric oxide synthase in normal and cirrhotic human liver. Proc. Natl. Acad. Sci. USA 2002, 99, 17161–17166. [Google Scholar]
- Mayer, B.; Hemmens, B. Biosynthesis and action of nitric oxide in mammalian cell. Trends Biochem. Sci 1997, 22, 477–481. [Google Scholar]
- Lin, Y.S.; Tsai, Y.J.; Tsay, J.S.; Lin, J.K. Factors affecting the levels of tea polyphenols and caffeine in tea leaves. J. Agric. Food Chem 2003, 51, 1864–1873. [Google Scholar]
- Beckman, J.S.; Koppenol, W.H. Nitric oxide, superoxide, and peroxynitrite: The good, the bad, and ugly. Am. J. Physiol 1996, 271, 1424–1437. [Google Scholar]
- Cooke, J.P.; Dzau, V.J. Nitric oxide synthase: Role in the genesis of vascular disease. Annu. Rev. Med 1997, 48, 489–509. [Google Scholar]
- Yermilov, V.; Rubio, J.; Becchi, M.; Friesen, M.D.; Pignatelli, B.; Ohshima, H. Formation of 8-nitroguanine by the reaction of guanine with peroxynitrite in vitro. Carcinogenesis 1996, 16, 2045–2050. [Google Scholar]
- Jie, G.; Lin, Z.; Zhang, L.; Lv, H.; He, P.; Zhao, B. Free radical scavenging effect of pu-erh tea extracts and their protective effect on oxidative damage in human fibroblast cells. J. Agric. Food Chem 2006, 54, 8058–8064. [Google Scholar]
- Luis, M.; Irene, C.; Jose, A.B.; Pedro, R. Antioxidant effect of rosemary, borage, green tea, pu-erh tea and ascorbic acid on fresh pork sausages packaged in a modified atmosphere: Influence of the presence of sodium chloride. J. Agric. Food Chem 2006, 86, 1298–1307. [Google Scholar]
- Jeng, K.C.; Chen, C.S.; Fang, Y.P.; Hou, R.C.; Chen, Y.S. Effect of microbial fermentation on content of statin, GABA, and polyphones in pu-erh tea. J. Agric. Food Chem 2007, 55, 8787–8792. [Google Scholar]
- Sano, M.; Takenaka, Y.; Kojima, R.; Saito, S.; Tomita, I.; Katou, M.; Shibuya, S. Effects of pu-erh tea on lipid metabolism in rats. Chem. Pharm. Bull. (Tokyo) 1986, 34, 221–228. [Google Scholar]
- Cao, Z.H.; Gu, D.H.; Lin, Q.Y.; Xu, Z.Q.; Huang, Q.C.; Rao, H.; Liu, E.W.; Jia, J.J.; Ge, C.R. Effect of pu-erh tea on body fat and lipid profiles in rats with diet-induced obesity. Phytother. Res 2011, 25, 234–238. [Google Scholar]
- Wu, S.C.; Yen, C.Y.; Wang, B.S.; Chih, C.K.; Yen, W.J.; Chang, L.W.; Duh, P.D. Antimutagenic and antimicrobial activities of pu-erh tea. Food Sci. Technol 2007, 40, 506–512. [Google Scholar]
- Yao, L.H.; Liu, X.; Jiang, Y.M.; Nola, C.; Bruce, D.; Riantong, S.; Nivedita, D.; Xu, Y. Compositional analysis of teas from Australian supermarkets. Food Chem 2006, 94, 115–122. [Google Scholar]
- Gong, J.; Peng, C.; Chen, T.; Gao, B.; Zhou, H. Effects of theabrownin from pu-erh tea on the metabolism of serum lipids in rats: Mechanism of action. J. Food Sci 2010, 75, 182–189. [Google Scholar]
- Wang, D.; Xiao, R.; Hu, X.; Xu, K.; Hou, Y.; Zhong, Y.; Meng, J.; Fan, B.; Liu, L. Comparative safety evaluation of Chinese pu-erh green tea extract and pu-erh black tea extract in wistar rats. J. Agric. Food Chem 2010, 58, 1350–1358. [Google Scholar]
- Yadav, S.K.; Ahuja, P.S. Towards generating caffeine-free tea by metabolic engineering. Plant Foods Hum. Nutr 2007, 62, 185–191. [Google Scholar]
- Xu, Y.; Zhao, H.; Zhang, M.; Li, C.J.; Lin, X.Z.; Sheng, J.; Shi, W. Variations of antioxidant properties and NO scavenging abilities during fermentation of tea. Int. J. Mol. Sci 2011, 12, 4574–4590. [Google Scholar]
- Wang, B.-S.; Yu, H.M.; Chang, L.-W.; Yen, W.-J.; Duh, P.-D. Protective effects of pu-erh tea on LDL oxidation and nitric oxide generation in macrophage cells. LWT Food Sci. Technol 2008, 41, 1122–1132. [Google Scholar]
- Kumamoto, T.; Togo, S.; Ishibe, A.; Morioka, D.; Watanabe, K.; Takahashi, T.; Shimizu, T.; Matsuo, K.; Kubota, T.; Tanaka, K.; et al. Role of nitricoxide synthesized bynitricoxide synthase 2 in liver regeneration. Liver Int 2008, 28, 865–877. [Google Scholar]
- Mizel, S.B.; Honko, A.N.; Moors, M.A.; Smith, P.S.; West, A.P. Induction of macrophage nitric oxide production by Gram-negative flagellin involves signaling via heteromeric Toll-like receptor 5/Toll-like receptor 4 complexes. J. Immunol 2003, 170, 6217–6223. [Google Scholar]
- Kadowaki, S.; Chikumi, H.; Yamamoto, H.; Yoneda, K.; Yamasaki, A.; Sato, K.; Shimizu, E. Down-regulation of inducible nitric oxide synthase by lysophosphatidic acid in human respiratory epithelial cells. Mol. Cell Biochem 2004, 262, 51–59. [Google Scholar]
- Davis, R.L.; Sanchez, A.C.; Lindley, D.J.; Williams, S.C.; Syapin, P.J. Effects of mechanistically distinct NF-κB inhibitors on glial inducible nitric-oxide synthase expression. Nitric Oxide 2005, 12, 200–209. [Google Scholar]
- Yang, Z.Y.; Tu, Y.Y.; Susanne, B.; Dong, F.; Xu, Y.; Naoharu, W. Isolation and identification of compounds from the ethanolic extract of flowers of the tea (Camellia sinensis) plant and their contribution to the antioxidant capacity. LWT Food Sci. Technol 2009, 42, 1439–1443. [Google Scholar]
- Sava, V.M.; Yang, S.M.; Hong, M.Y.; Yang, P.C.; Guewha, S.H. Isolation and characterization of melanic pigments derived from tea and tea polyphenols. Food Chem 2001, 73, 177–184. [Google Scholar]
- Laurentin, A.; Edwards, C.A. A microtiter modification of the anthrone-sulfuric acid colorimetric assay for glucose-based carbohydrates. Anal. Biochem 2003, 315, 143–145. [Google Scholar]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem 1976, 72, 248–254. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative CT method. Nat. Protoc 2008, 3, 1101–1108. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primers | Tm |
|---|---|---|
| iNOS | Sense: 5′-CAGATCCCGAAACGCTACAC-3′ | 60.05 °C |
| Antisense: 5′-TGCGGCTGGACTTCTCACT-3′ | 59.5 °C | |
| eNOS | Sense: 5′-CTGCTGCCCCAGATATCTTC-3′ | 60.5 °C |
| Antisense: 5′-CAGGTACTGCAGTCCCTCCT-3′ | 62.5 °C | |
| β-actin | Sense: 5′-AACCCTAAGGCCAACCGTGAAAAG-3′ | 59.4 °C |
| Antisense: 5′-TCATGAGGTAGTCTGTCAGGT-3′ | 61.37 °C |
© 2012 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Xu, Y.; Wang, G.; Li, C.; Zhang, M.; Zhao, H.; Sheng, J.; Shi, W. Pu-erh Tea Reduces Nitric Oxide Levels in Rats by Inhibiting Inducible Nitric Oxide Synthase Expression through Toll-Like Receptor 4. Int. J. Mol. Sci. 2012, 13, 7174-7185. https://doi.org/10.3390/ijms13067174
Xu Y, Wang G, Li C, Zhang M, Zhao H, Sheng J, Shi W. Pu-erh Tea Reduces Nitric Oxide Levels in Rats by Inhibiting Inducible Nitric Oxide Synthase Expression through Toll-Like Receptor 4. International Journal of Molecular Sciences. 2012; 13(6):7174-7185. https://doi.org/10.3390/ijms13067174
Chicago/Turabian StyleXu, Yang, Guan Wang, Chunjie Li, Min Zhang, Hang Zhao, Jun Sheng, and Wei Shi. 2012. "Pu-erh Tea Reduces Nitric Oxide Levels in Rats by Inhibiting Inducible Nitric Oxide Synthase Expression through Toll-Like Receptor 4" International Journal of Molecular Sciences 13, no. 6: 7174-7185. https://doi.org/10.3390/ijms13067174
APA StyleXu, Y., Wang, G., Li, C., Zhang, M., Zhao, H., Sheng, J., & Shi, W. (2012). Pu-erh Tea Reduces Nitric Oxide Levels in Rats by Inhibiting Inducible Nitric Oxide Synthase Expression through Toll-Like Receptor 4. International Journal of Molecular Sciences, 13(6), 7174-7185. https://doi.org/10.3390/ijms13067174