The Opuntia streptacantha OpsHSP18 Gene Confers Salt and Osmotic Stress Tolerance in Arabidopsis thaliana


Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
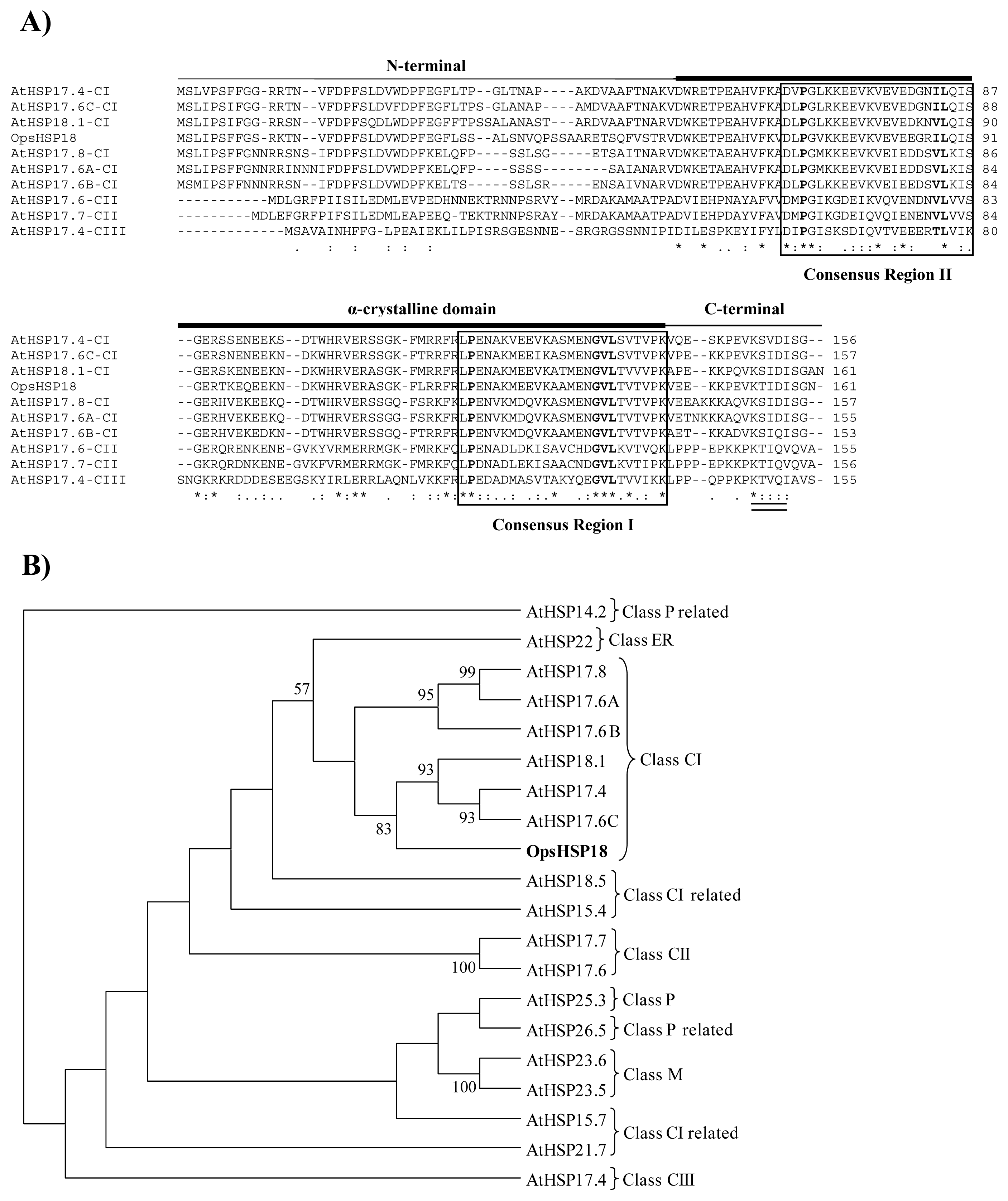
2.1. The OpsHSP18 Gene Encodes a Small Heat Shock Protein (sHSP)
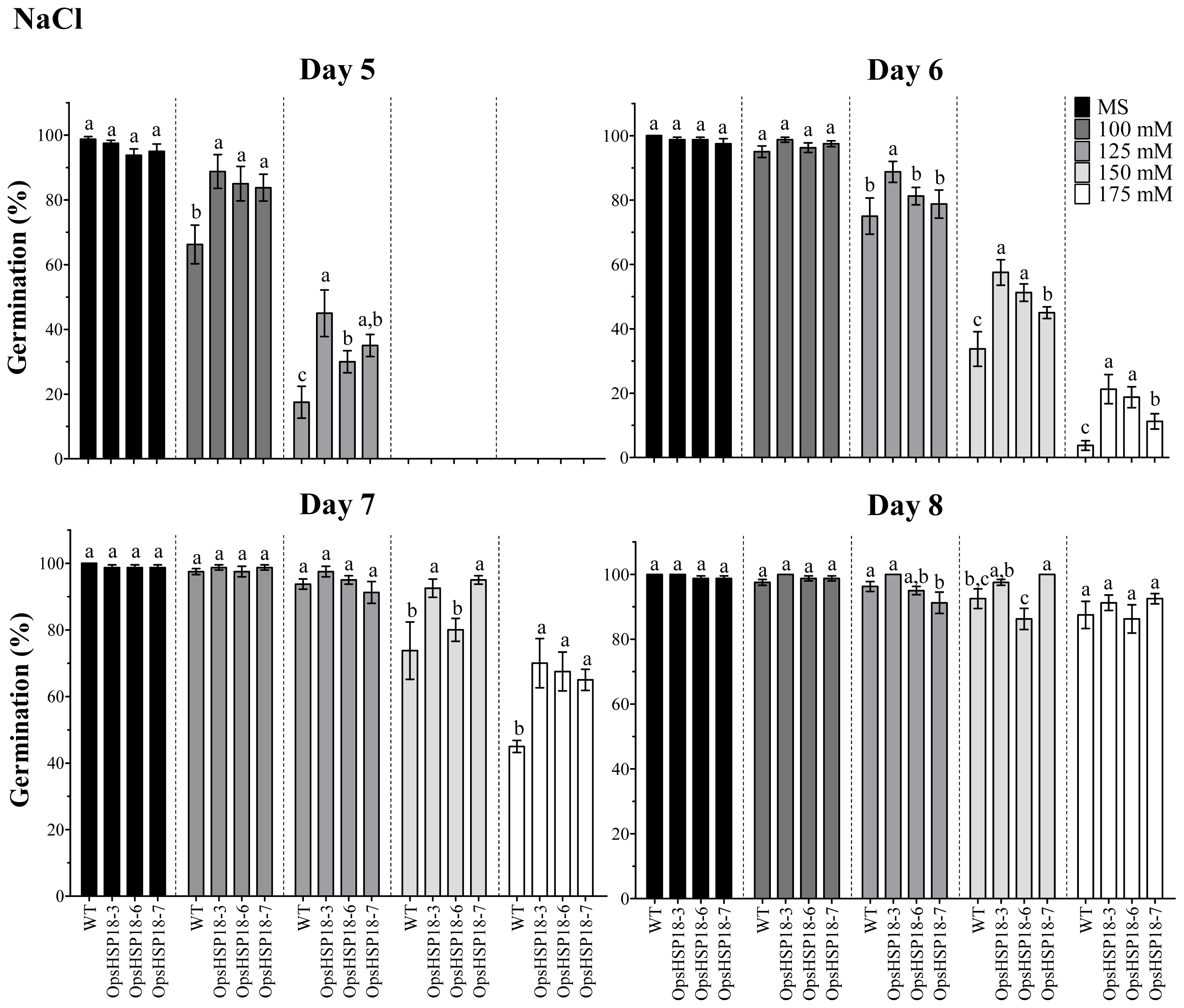
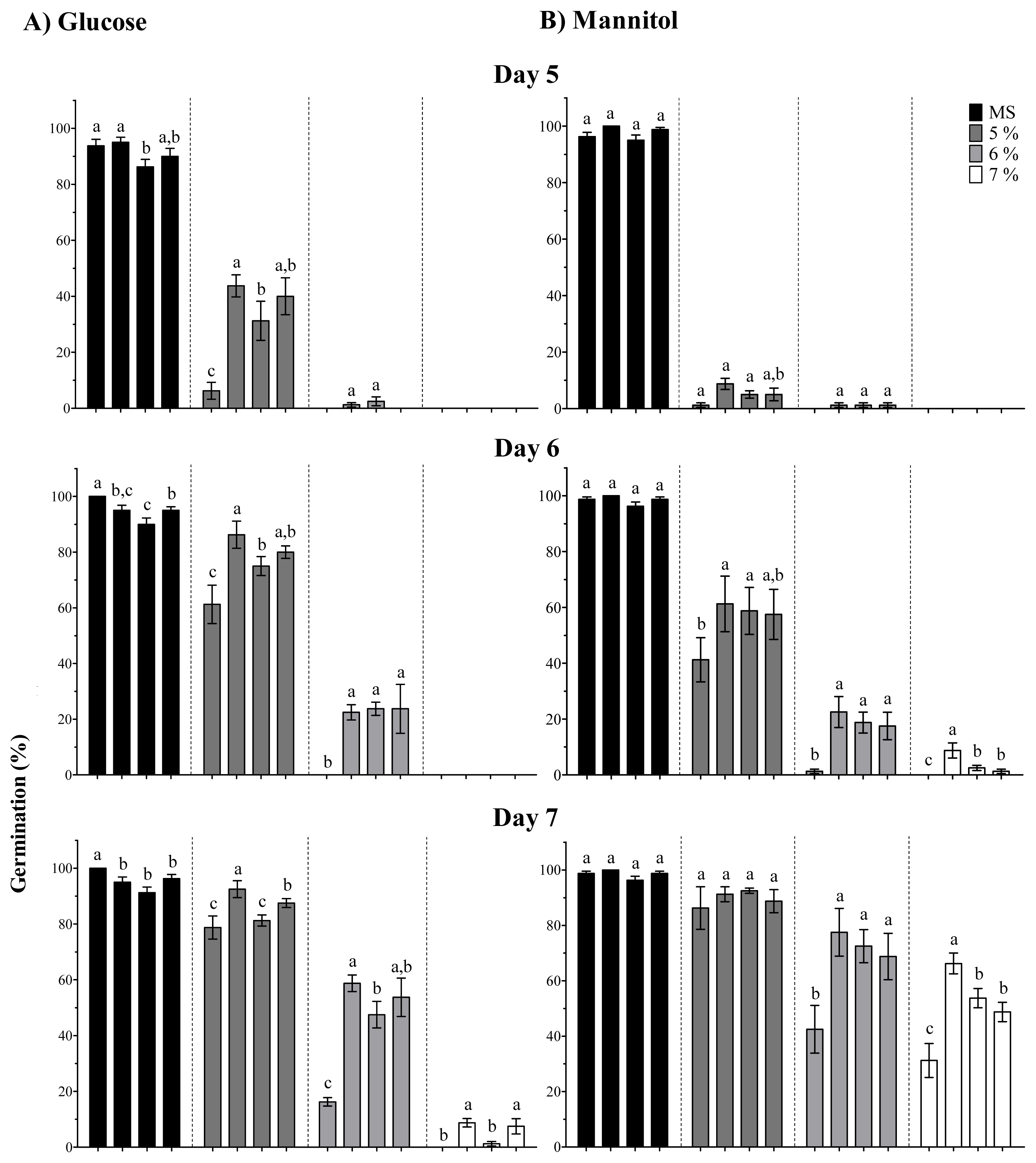
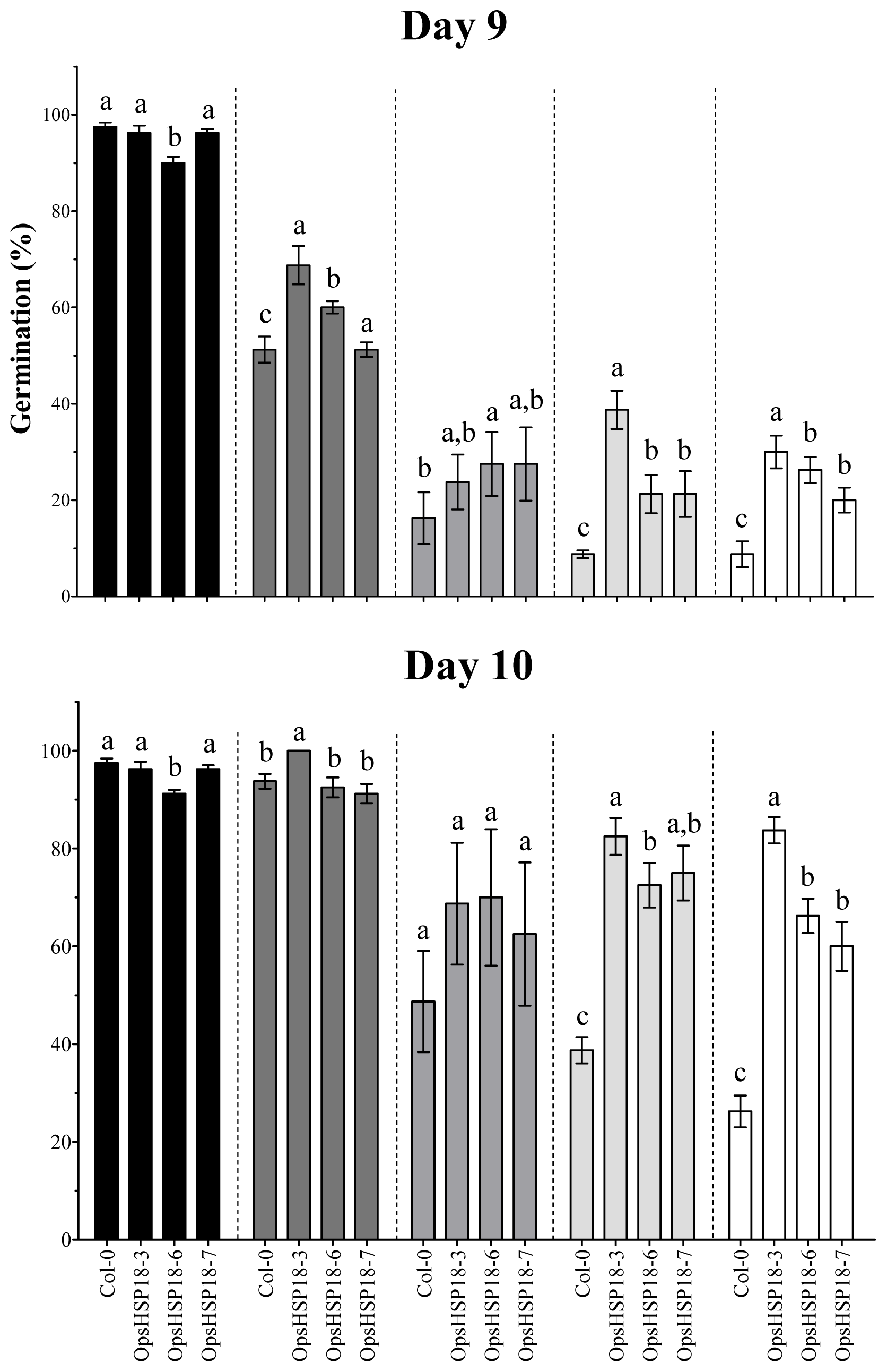
2.2. OpsHSP18 Gene Is Involved in the Response to Abiotic Stress during Seed Germination
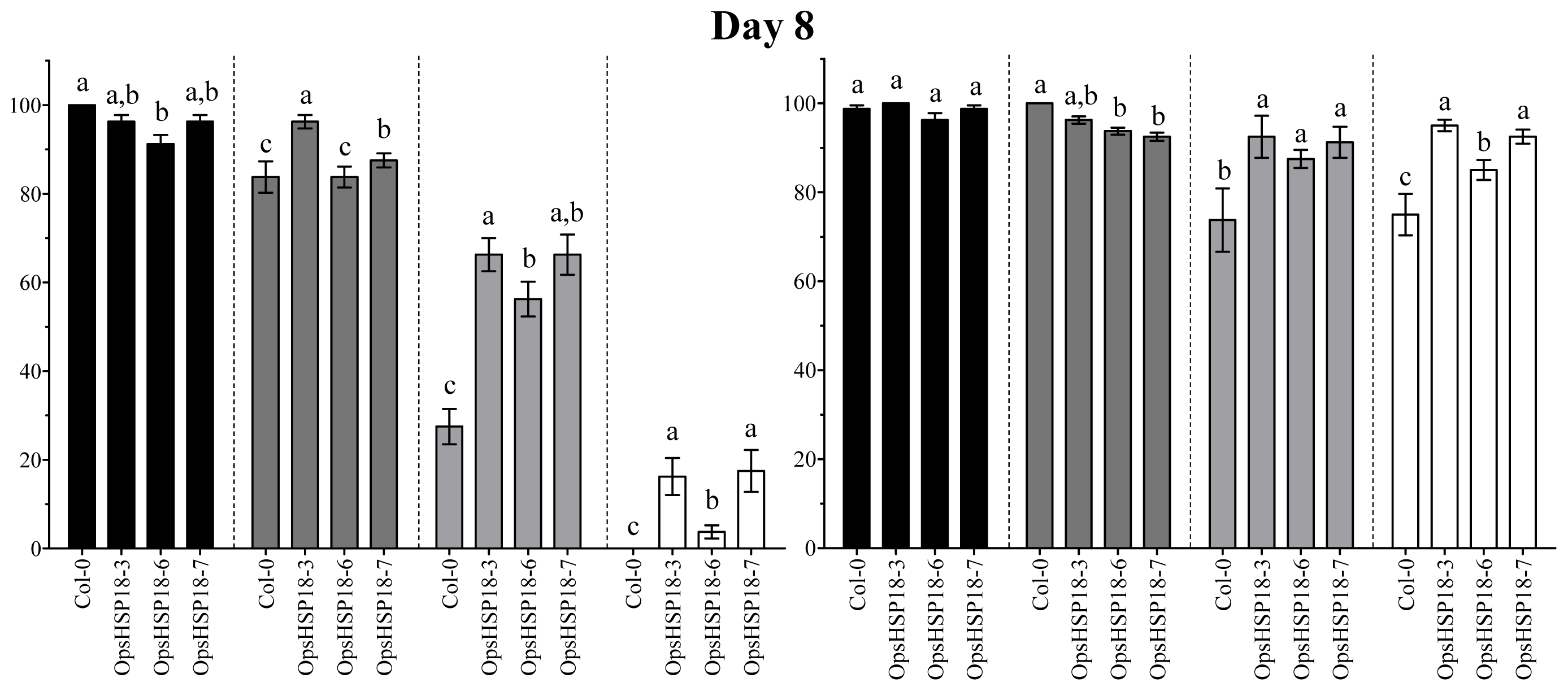
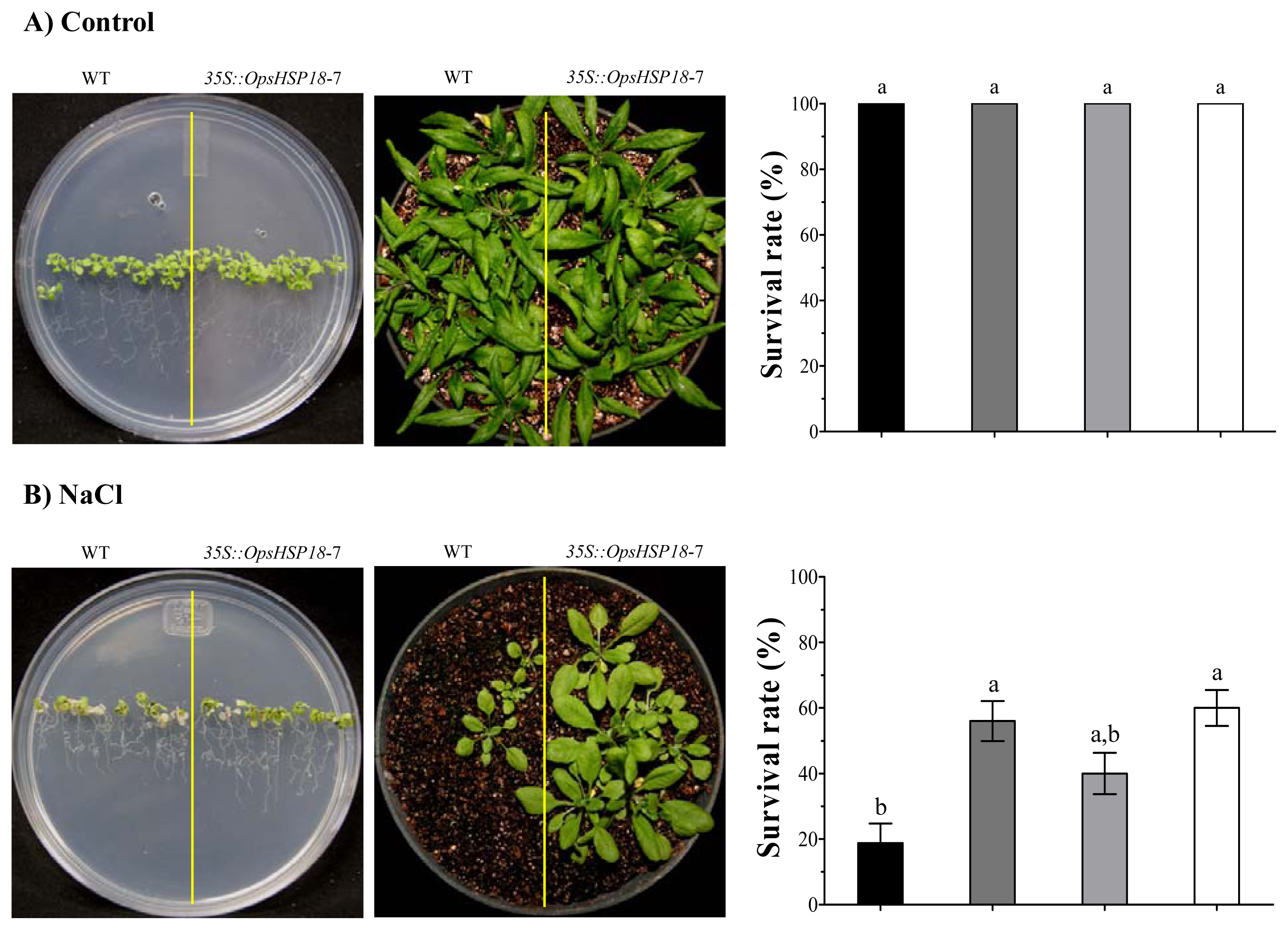
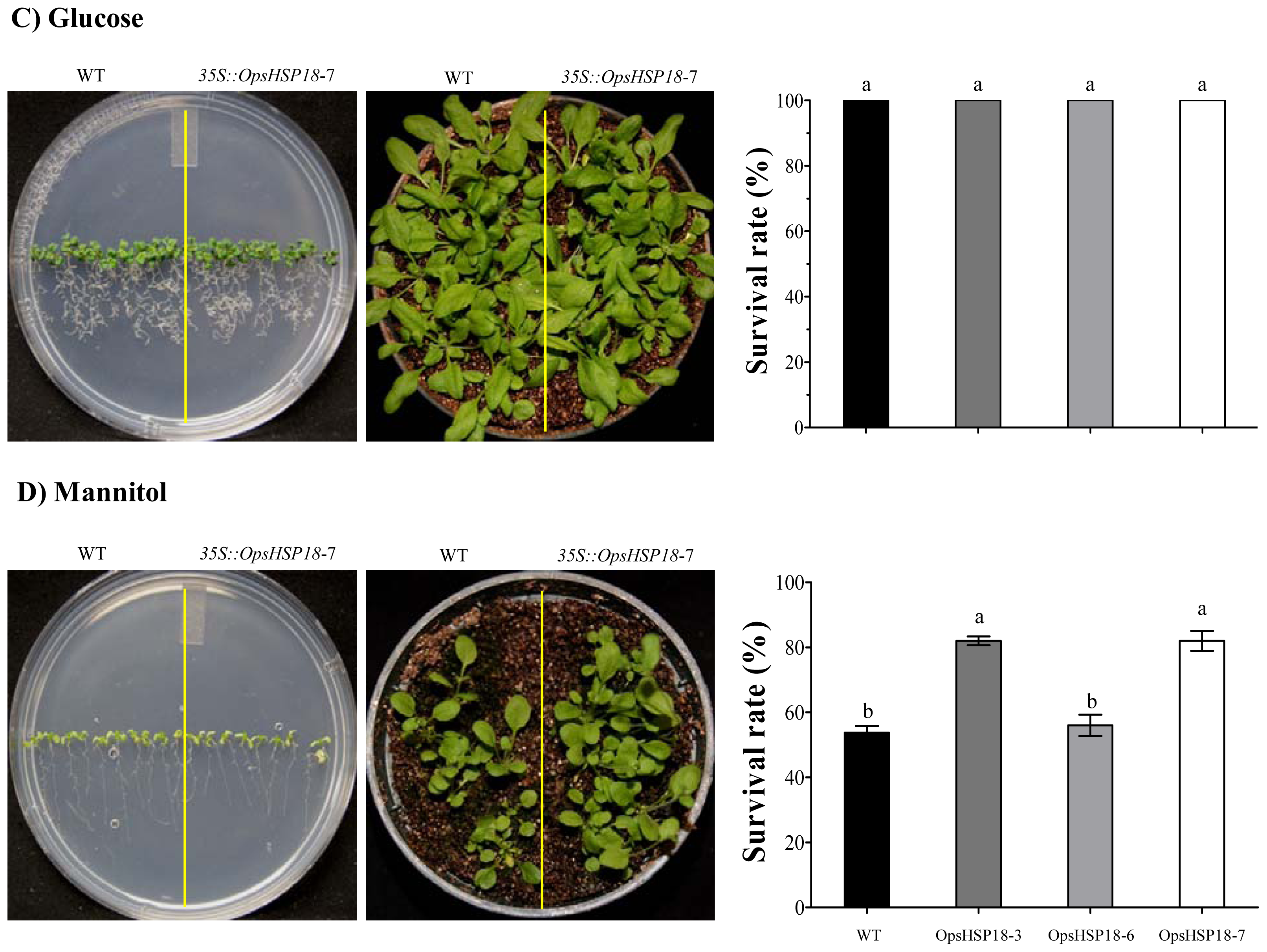
2.3. The Over-Expression of the OpsHSP18 Gene Allows A. thaliana Transgenic Plants Recover from Abiotic Stress
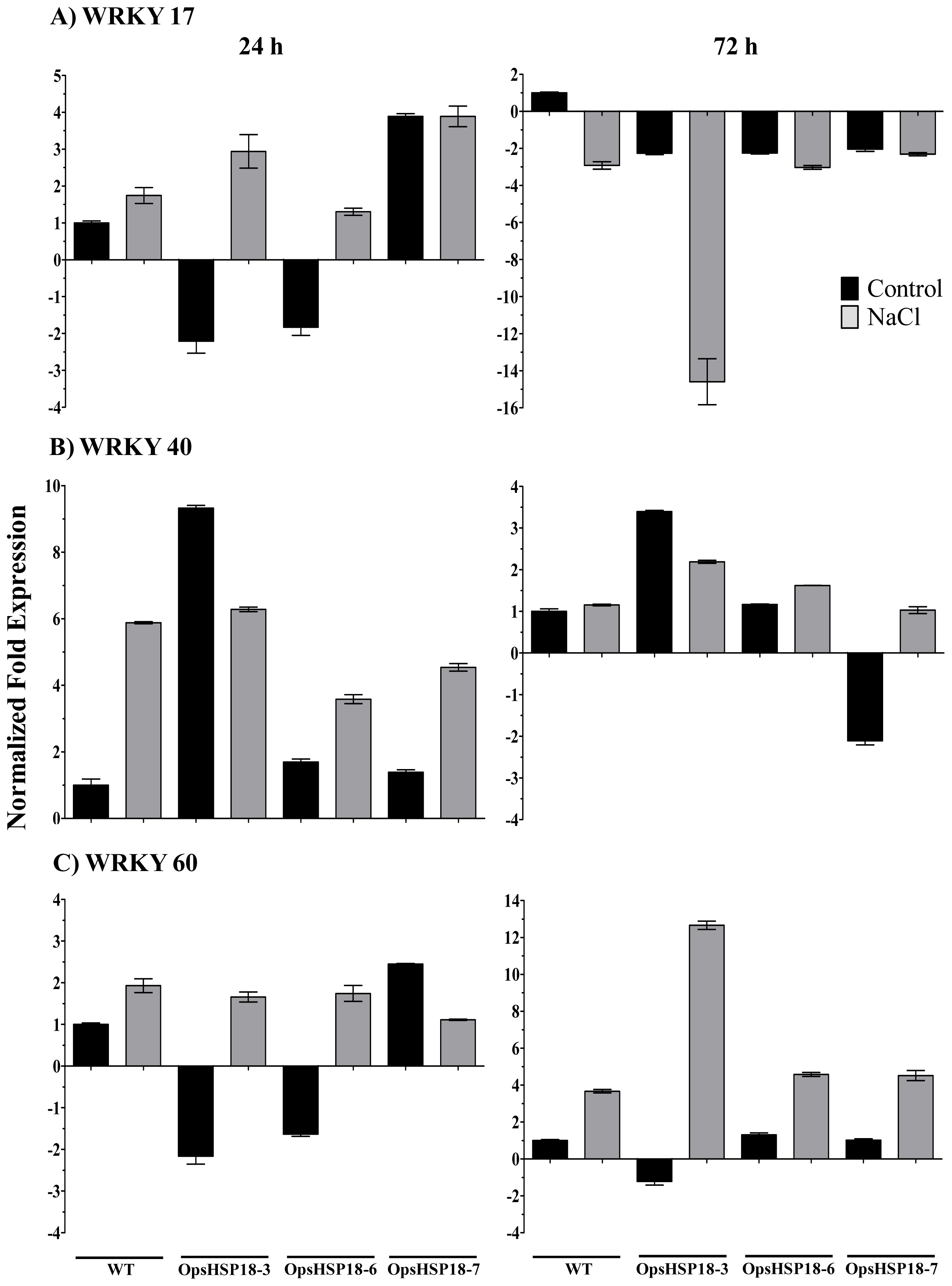
2.4. Expression of Stress Related Genes in the A. thaliana OpsHSP18 Transgenic Plants
3. Discussion
4. Experimental Section
4.1. Plant Material and Growth Conditions
4.2. Inverse Polymerase Chain Reaction (IPCR)
4.3. Generation of Arabidopsis thaliana Over-Expressing Lines
4.4. RNA Isolation, RT-PCR and Real-Time qRT-PCR Analyses
4.5. Germination Assays under Stress and Hormone Treatments
4.6. Estimation of Seedling Survival Rates under Stress Treatments
4.7. In Silico Analysis of the OpsHSP18 Protein
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
ijms-13-10154-s001.pdfAcknowledgments
References
- Mahajan, S.; Tuteja, N. Cold, salinity and drought stresses: An overview. Arch. Biochem. Biophys 2005, 444, 139–158. [Google Scholar]
- Cushman, J.C.; Bohnert, H.J. Genomic approaches to plant stress tolerance. Curr. Opin. Plant Biol 2000, 3, 117–124. [Google Scholar]
- Zhu, J.K. Salt and drought stress signal transduction in plants. Annu. Rev. Plant Biol 2002, 53, 247–273. [Google Scholar]
- Wang, W.; Vinocur, B.; Altman, A. Plant responses to drought, salinity and extreme temperatures: Towards genetic engineering for stress tolerance. Planta 2003, 218, 1–14. [Google Scholar]
- Bartels, D.; Sunkar, R. Drought and salt tolerance in plants. Crit. Rev. Plant Sci 2005, 24, 23–58. [Google Scholar]
- Becker, J.; Craig, E.A. Heat-shock proteins as molecular chaperones. Rev. Eur. J. Biochem 1994, 219, 11–23. [Google Scholar]
- Parsell, D.A.; Lindquist, S. The function of heat -shock proteins in stress tolerance: Degradation and reactivation of damaged proteins. Annu. Rev. Genet 1993, 27, 437–496. [Google Scholar]
- Vierling, E. The roles of heat shock proteins in plants. Annu. Rev. Plant Physiol. Plant Mol. Biol 1991, 42, 579–620. [Google Scholar]
- Wang, W.; Vinocur, B.; Shoseyov, O.; Altman, A. Role of plant heat-shock proteins and molecular chaperones in the abiotic stress response. Trends Plant Sci 2004, 9, 244–252. [Google Scholar]
- Xue, Y.; Peng, R.; Xiong, A.; Li, X.; Zha, D.; Yao, Q. Over-expression of heat shock protein gene hsp26 in Arabidopsis thaliana enhances heat tolerance. Biol. Plant 2010, 54, 105–111. [Google Scholar]
- Vierling, E. The small heat shock proteins in plants are members of an ancient family of heat induced proteins. Acta Physiol. Plant 1997, 19, 539–547. [Google Scholar]
- Waters, E.R.; Lee, G.J.; Vierling, E. Evolution, structure and function of the small heat shock proteins in plants. J. Exp. Bot 1996, 47, 325–338. [Google Scholar]
- Sun, W.; Montagu, M.V.; Verbruggen, N. Small heat shock proteins and stress tolerance in plants. BBA-Gene Struct. Expr 2002, 1577, 1–9. [Google Scholar]
- Vierling, E.; Jaya, N.; Garcia, V. Substrate binding site flexibility of the small heat shock protein molecular chaperones. Proc. Natl. Acad. Sci. USA 2009, 106, 15604–15609. [Google Scholar]
- Stamler, R.; Kappe, G.; Boelens, W.; Slingsby, C. Wrapping the α-crystallin domain fold in a chaperone assembly. J. Mol. Biol 2005, 353, 68–79. [Google Scholar]
- Sun, Y.; MacRae, T.H. Small heat shock proteins: Molecular structure and chaperone function. Cell Mol. Life Sci 2005, 62, 2460–2476. [Google Scholar]
- Haslbeck, M.; Franzmann, T.; Weinfurtner, D.; Buchner, J. Some like it hot: The structure and function of small heat-shock proteins. Nat. Struct. Mol. Biol 2005, 12, 842–846. [Google Scholar]
- Scharf, K.D.; Siddique, M.; Vierling, E. The expanding family of Arabidopsis thaliana small heat stress proteins and a new family of proteins containing α-crystallin domains (Acd proteins). Cell Stress Chaperon 2001, 6, 225–237. [Google Scholar]
- Winter, K.; Aranda, J.; Holtum, J.A.M. Carbon isotope composition and water-use efficiency in plants with crassulacean acid metabolism. Funct. Plant Biol 2005, 32, 381–388. [Google Scholar]
- Nobel, P.S.; Bobich, E.G. Initial net CO2 uptake responses and root growth for a CAM community placed in a closed environment. Ann. Bot 2002, 90, 593–598. [Google Scholar]
- Rabas, A.R.; Martin, C.E. Movement of water from old to young leaves in three species succulents. Ann. Bot 2003, 92, 529–536. [Google Scholar]
- Cushman, J.C. Crassulacean Acid Metabolism. A plastic photosynthetic adaptation to arid environments. Plant Physiol 2001, 127, 1439–1448. [Google Scholar]
- Luttge, U. Ecophysiology of Crassulacean Acid Metabolism (CAM). Ann. Bot 2004, 93, 629–652. [Google Scholar]
- Ochoa-Alfaro, A.E.; Rodríguez-Kessler, M.; Pérez-Morales, M.B.; Delgado-Sánchez, P.; Cuevas-Velazquez, C.L.; Gómez-Anduro, G.; Jiménez-Bremont, J.F. Functional characterization of an acidic SK3 dehydrin isolated from an Opuntia streptacantha cDNA library. Planta 2012, 235, 565–557. [Google Scholar]
- Assab, E.; Rampino, P.; Mita, G.; Perrotta, C. Heat shock response in olive (Olea europaea L.) twigs: Identification and analysis of a cDNA coding a class I small heat shock protein. Plant Biosyst 2011, 145, 419–425. [Google Scholar]
- Mahmood, T.; Safdar, W.; Abbasi, B.H.; Naqvi, S.M.S. An overview on the small heat shock proteins. Afr. J. Biotechnol 2010, 9, 927–949. [Google Scholar]
- De Jong, W.W.; Caspers, G.J.; Leunissen, J.A.M. Genealogy of the α-crystallin small heat-stress superfamily. Int. J. Biol. Macromol 1998, 22, 151–162. [Google Scholar]
- Felsenstein, J. PHYLIP-phylogeny inference package (version 3.2). Cladistics 1989, 5, 164–166. [Google Scholar]
- Sairam, R.K.; Tyagi, A. Physiology and molecular biology of salinity stress tolerance in plants. Curr. Sci 2004, 86, 407–421. [Google Scholar]
- Kim, K.K.; Kim, R.; Kim, S.H. Crystal structure of a small heat-shock protein. Nature 1998, 394, 595–599. [Google Scholar]
- Wehmeyer, N.; Hernandez, L.D.; Finkelstein, R.R.; Vierling, E. Synthesis of small heat-shock proteins is part of the developmental program of late seed maturation. Plant Physiol 1996, 112, 747–757. [Google Scholar]
- Wehmeyer, N.; Vierling, E. The expression of small heat shock proteins in seeds responds to discrete developmental signals and suggests a general protective role in desiccation tolerance. Plant Physiol 2000, 122, 1099–1108. [Google Scholar]
- Volkov, R.A.; Panchuk, I.I.; Schöffl, F. Small heat shock proteins are differentially regulated during pollen development and following heat stress in tobacco. Plant Mol. Biol 2005, 57, 487–502. [Google Scholar]
- Zhou, Y.; Chen, H.; Chu, P.; Li, Y.; Tan, B.; Ding, Y.; Tsang, E.W.T.; Jiang, L.; Wu, K.; Huang, S. NnHSP17.5, a cytosolic class II small heat shock protein gene from Nelumbo nucifera, contributes to seed germination vigor and seedling thermotolerance in transgenic Arabidopsis. Plant Cell Rep 2012, 31, 379–389. [Google Scholar]
- Neta-Sharir, I.; Isaacson, T.; Lurie, S.; Weissa, D. Dual role for tomato heat shock protein 21: Protecting photosystem II from oxidative stress and promoting color changes during fruit maturation. Plant Cell 2005, 17, 1829–1838. [Google Scholar]
- Takahashi, T.; Komeda, Y. Characterization of two genes encoding small heat-shock proteins in Arabidopsis thaliana. Mol. Gen. Genet 1989, 219, 365–372. [Google Scholar]
- Helm, K.W.; Vierling, E. An Arabidopsis thaliana cDNA clone encoding a low molecular weight heat shock protein. Nuc. Acids Res 1989, 17, 7995. [Google Scholar]
- Sun, W.; Bernard, C.; van de Cotte, B.; Montagu, M.; Montagu, M.V.; Verbruggen, N. At-HSP17.6A, encoding a small heat-shock protein in Arabidopsis, can enhance osmotolerance upon overexpression. Plant J 2001, 27, 407–415. [Google Scholar]
- Murakami, T.; Matsuba, S.; Funatsuki, H.; Kawaguchi, K.; Saruyama, H.; Tanida, M.; Sato, Y. Over-expression of a small heat shock protein, sHSP17.7, confers both heat tolerance and UV-B resistance to rice plants. Mol. Breed. 2004, 13, 165–175. [Google Scholar]
- Sato, Y.; Yokoya, S. Enhanced tolerance to drought stress in transgenic rice plants overexpressing a small heat-shock protein, sHSP17.7. Plant Cell 2008, 27, 329–334. [Google Scholar]
- Jiang, Y.; Deyholos, M.K. Comprehensive transcriptional profiling of NaCl-stressed Arabidopsis roots reveals novel classes of responsive genes. BMC Plant Biol 2006, 6, 1–20. [Google Scholar]
- Chen, H.; Lai, Z.; Shi, J.; Xiao, Y.; Chen, Z.; Xu, X. Roles of arabidopsis WRKY18, WRKY40 and WRKY60 transcription factors in plant responses to abscisic acid and abiotic stress. BMC Plant Biol 2010, 10, 1–15. [Google Scholar]
- Zahur, M.; Maqbool, A.; Irfan, M.; Barozai, M.Y.K.; Qaiser, U.; Rashid, B.; Husnain, T.; Riazuddin, S. Functional analysis of cotton small heat shock protein promoter region in response to abiotic stresses in tobacco using Agrobacterium-mediated transient assay. Mol. Biol. Rep 2009, 36, 1915–1921. [Google Scholar]
- Zhang, X.; Henriques, R.; Lin, S.S.; Niu, Q.W.; Chua, N.H. Agrobacterium-mediated transformation of Arabidopsis thaliana using the floral dip method. Nat. Prot 2006, 1, 641–646. [Google Scholar]
- Zhao, Y.; Wang, T.; Zhang, W.; Li, X. SOS3 mediates lateral root development under low salt stress through regulation of auxin redistribution and maxima in Arabidopsis. New Phytol 2011, 189, 1122–1134. [Google Scholar]
- Swiss Institute of Bioinformatics. Available online: www.isb-sib.ch accessed on 1 December 2011.
- Phytozome database. Available online: www.phytozome.net accessed on 10 December 2011.
- National Center for Biotechnology Information. Available online: www.ncbi.nlm.nih.gov accessed on 10 December 2011.
- Motif Scan tool at the Expasy proteomics server. Available online: http://hits.isb-sib.ch/cgi-bin/motif_scan accessed on 1 December 2011.
- T-Coffee programs at the European Bioinformatics Institute. Available online: www.ebi.ac.uk accessed on 10 December 2011.
- Veerassamy, S.; Smith, A.; Tillier, E.R.M. A transition probability model for amino acid substitutions from blocks. J. Comp. Biol 2003, 10, 997–1010. [Google Scholar]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol 2011, 28, 2731–2739. [Google Scholar]











© 2012 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Salas-Muñoz, S.; Gómez-Anduro, G.; Delgado-Sánchez, P.; Rodríguez-Kessler, M.; Jiménez-Bremont, J.F. The Opuntia streptacantha OpsHSP18 Gene Confers Salt and Osmotic Stress Tolerance in Arabidopsis thaliana. Int. J. Mol. Sci. 2012, 13, 10154-10175. https://doi.org/10.3390/ijms130810154
Salas-Muñoz S, Gómez-Anduro G, Delgado-Sánchez P, Rodríguez-Kessler M, Jiménez-Bremont JF. The Opuntia streptacantha OpsHSP18 Gene Confers Salt and Osmotic Stress Tolerance in Arabidopsis thaliana. International Journal of Molecular Sciences. 2012; 13(8):10154-10175. https://doi.org/10.3390/ijms130810154
Chicago/Turabian StyleSalas-Muñoz, Silvia, Gracia Gómez-Anduro, Pablo Delgado-Sánchez, Margarita Rodríguez-Kessler, and Juan Francisco Jiménez-Bremont. 2012. "The Opuntia streptacantha OpsHSP18 Gene Confers Salt and Osmotic Stress Tolerance in Arabidopsis thaliana" International Journal of Molecular Sciences 13, no. 8: 10154-10175. https://doi.org/10.3390/ijms130810154
APA StyleSalas-Muñoz, S., Gómez-Anduro, G., Delgado-Sánchez, P., Rodríguez-Kessler, M., & Jiménez-Bremont, J. F. (2012). The Opuntia streptacantha OpsHSP18 Gene Confers Salt and Osmotic Stress Tolerance in Arabidopsis thaliana. International Journal of Molecular Sciences, 13(8), 10154-10175. https://doi.org/10.3390/ijms130810154