Transforming Growth Factor-Beta-Induced Protein (TGFBI)/(βig-H3): A Matrix Protein with Dual Functions in Ovarian Cancer


Abstract
:1. Introduction
2. βig-H3 Regulation and Function
3. βig-H3 Roles in Disease
3.1. Role in Diabetes and Corneal Dystrophies
3.2. Roles in Cancer
3.2.1. Role as Tumor Suppressor
3.2.2. Roles as Tumor Promoter
3.2.3. Role of βig-H3 in Ovarian Cancer
3.2.3.1. Tumor Suppressive role of βig-H3 in Ovarian Cancer
3.2.3.2. Pro-Tumorigenic Role of βig-H3 in Ovarian Cancer
3.2.3.3. βig-H3 Processing by Ovarian Cancer Peritoneal Interactions
3.2.3.4. βig-H3 as a Predictor of Therapy Response
4. Conclusions
Acknowledgments
References
- Skonier, J.; Neubauer, M.; Madisen, L.; Bennett, K.; Plowman, G.D.; Purchio, A.F. cDNA cloning and sequence analysis of beta ig-h3, a novel gene induced in a human adenocarcinoma cell line after treatment with transforming growth factor-beta. DNA Cell Biol 1992, 11, 511–522. [Google Scholar]
- Becker, J.; Erdlenbruch, B.; Noskova, I.; Schramm, A.; Aumailley, M.; Schorderet, D.F.; Schweigerer, L. Keratoepithelin suppresses the progression of experimental human neuroblastomas. Cancer Res 2006, 66, 5314–5321. [Google Scholar]
- Hashimoto, K.; Noshiro, M.; Ohno, S.; Kawamoto, T.; Satakeda, H.; Akagawa, Y.; Nakashima, K.; Okimura, A.; Ishida, H.; Okamoto, T.; et al. Characterization of a cartilage-derived 66-kDa protein (RGD-CAP/beta ig-h3) that binds to collagen. Biochim. Biophys. Acta 1997, 1355, 303–314. [Google Scholar]
- Gibson, M.A.; Kumaratilake, J.S.; Cleary, E.G. Immunohistochemical and ultrastructural localization of MP78/70 (betaig-h3) in extracellular matrix of developing and mature bovine tissues. J. Histochem. Cytochem 1997, 45, 1683–1696. [Google Scholar]
- Liu, C.; Li, B.; Cheng, Y.; Lin, J.; Hao, J.; Zhang, S.; Mitchel, R.E.; Sun, D.; Ni, J.; Zhao, L.; Gao, F.; Cai, J. MiR-21 plays an important role in radiation induced carcinogenesis in BALB/c mice by directly targeting the tumor suppressor gene Big-h3. Int. J. Biol. Sci 2011, 7, 347–363. [Google Scholar]
- Sasaki, H.; Kobayashi, Y.; Nakashima, Y.; Moriyama, S.; Yukiue, H.; Kaji, M.; Kiriyama, M.; Fukai, I.; Yamakawa, Y.; Fujii, Y. Beta IGH3, a TGF-beta inducible gene, is overexpressed in lung cancer. Jpn. J. Clin. Oncol 2002, 32, 85–89. [Google Scholar]
- Rawe, I.M.; Zhan, Q.; Burrows, R.; Bennett, K.; Cintron, C. Beta-ig. Molecular cloning and in situ hybridization in corneal tissues. Invest. Ophthalmol. Vis. Sci 1997, 38, 893–900. [Google Scholar]
- Gibson, M.A.; Kumaratilake, J.S.; Cleary, E.G. The protein components of the 12-nanometer microfibrils of elastic and nonelastic tissues. J. Biol. Chem 1989, 264, 4590–4598. [Google Scholar]
- Schorderet, D.F.; Menasche, M.; Morand, S.; Bonnel, S.; Buchillier, V.; Marchant, D.; Auderset, K.; Bonny, C.; Abitbol, M.; Munier, F.L. Genomic characterization and embryonic expression of the mouse Bigh3 (Tgfbi) gene. Biochem. Biophys. Res. Commun 2000, 274, 267–274. [Google Scholar]
- LeBaron, R.G.; Bezverkov, K.I.; Zimber, M.P.; Pavelec, R.; Skonier, J.; Purchio, A.F. Beta IG-H3, a novel secretory protein inducible by transforming growth factor-beta, is present in normal skin and promotes the adhesion and spreading of dermal fibroblasts in vitro. J. Invest. Dermatol 1995, 104, 844–849. [Google Scholar]
- Kim, J.E.; Kim, S.J.; Lee, B.H.; Park, R.W.; Kim, K.S.; Kim, I.S. Identification of motifs for cell adhesion within the repeated domains of transforming growth factor-beta-induced gene, betaig-h3. J. Biol. Chem 2000, 275, 30907–30915. [Google Scholar]
- Thapa, N.; Lee, B.H.; Kim, I.S. TGFBIp/betaig-h3 protein: A versatile matrix molecule induced by TGF-beta. Int. J. Biochem. Cell Biol 2007, 39, 2183–2194. [Google Scholar]
- Uekita, T.; Kim, Y.J.; Yamanouchi, K.; Tojo, H.; Tachi, C. Dynamics of betaig-h3 mRNA expression during pregnancy in the uterus and the placenta of the mouse: A possible regulatory factor for trophoblastic invasion. J. Reprod. Dev 2003, 49, 243–252. [Google Scholar]
- Carson, D.D.; Lagow, E.; Thathiah, A.; Al-Shami, R.; Farach-Carson, M.C.; Vernon, M.; Yuan, L.; Fritz, M.A.; Lessey, B. Changes in gene expression during the early to mid-luteal (receptive phase) transition in human endometrium detected by high-density microarray screening. Mol. Hum. Reprod 2002, 8, 871–879. [Google Scholar]
- Gratchev, A.; Guillot, P.; Hakiy, N.; Politz, O.; Orfanos, C.E.; Schledzewski, K.; Goerdt, S. Alternatively activated macrophages differentially express fibronectin and its splice variants and the extracellular matrix protein betaIG-H3. Scand. J. Immunol 2001, 53, 386–392. [Google Scholar]
- Yun, S.J.; Kim, M.O.; Kim, S.O.; Park, J.; Kwon, Y.K.; Kim, I.S.; Lee, E.H. Induction of TGF-beta-inducible gene-h3 (betaig-h3) by TGF-beta1 in astrocytes: Implications for astrocyte response to brain injury. Brain Res. Mol. Brain Res 2002, 107, 57–64. [Google Scholar]
- Dokmanovic, M.; Chang, B.D.; Fang, J.; Roninson, I.B. Retinoid-induced growth arrest of breast carcinoma cells involves co-activation of multiple growth-inhibitory genes. Cancer Biol. Ther 2002, 1, 24–27. [Google Scholar]
- Nam, E.J.; Sa, K.H.; You, D.W.; Cho, J.H.; Seo, J.S.; Han, S.W.; Park, J.Y.; Kim, S.I.; Kyung, H.S.; Kim, I.S.; et al. Up-regulated transforming growth factor beta-inducible gene h3 in rheumatoid arthritis mediates adhesion and migration of synoviocytes through αvβ3 integrin: Regulation by cytokines. Arthritis Rheum 2006, 54, 2734–2744. [Google Scholar]
- Lee, S.H.; Kim, E.J.; Suk, K.; Kim, I.S.; Lee, W.H. TL1A induces the expression of TGF-beta-inducible gene h3 (βig-h3) through PKC, PI3K, and ERK in THP-1 cells. Cell. Immunol 2010, 266, 61–66. [Google Scholar]
- Shin, S.H.; Kim, J.; Heo, S.C.; Kwon, Y.W.; Kim, Y.M.; Kim, I.S.; Lee, T.G.; Kim, J.H. Proteomic identification of betaig-h3 as a lysophosphatidic acid-induced secreted protein of human mesenchymal stem cells: Paracrine activation of A549 lung adenocarcinoma cells by betaig-h3. Mol. Cell. Proteomics 2012, 11. [Google Scholar] [CrossRef]
- Lee, B.H.; Bae, J.S.; Park, R.W.; Kim, J.E.; Park, J.Y.; Kim, I.S. Betaig-h3 triggers signaling pathways mediating adhesion and migration of vascular smooth muscle cells through αvβ5 integrin. Exp. Mol. Med 2006, 38, 153–161. [Google Scholar]
- O’Brien, E.R.; Bennett, K.L.; Garvin, M.R.; Zderic, T.W.; Hinohara, T.; Simpson, J.B.; Kimura, T.; Nobuyoshi, M.; Mizgala, H.; Purchio, A.; et al. Beta ig-h3, a transforming growth factor-beta-inducible gene, is overexpressed in atherosclerotic and restenotic human vascular lesions. Arterioscler. Thromb. Vasc. Biol 1996, 16, 576–584. [Google Scholar]
- Dieudonne, S.C.; Kerr, J.M.; Xu, T.; Sommer, B.; DeRubeis, A.R.; Kuznetsov, S.A.; Kim, I.S.; Gehron Robey, P.; Young, M.F. Differential display of human marrow stromal cells reveals unique mRNA expression patterns in response to dexamethasone. J. Cell. Biochem 1999, 76, 231–243. [Google Scholar]
- Monticone, M.; Liu, Y.; Tonachini, L.; Mastrogiacomo, M.; Parodi, S.; Quarto, R.; Cancedda, R.; Castagnola, P. Gene expression profile of human bone marrow stromal cells determined by restriction fragment differential display analysis. J. Cell. Biochem 2004, 92, 733–744. [Google Scholar]
- Thapa, N.; Kang, K.B.; Kim, I.S. Beta ig-h3 mediates osteoblast adhesion and inhibits differentiation. Bone 2005, 36, 232–242. [Google Scholar]
- Billings, P.C.; Whitbeck, J.C.; Adams, C.S.; Abrams, W.R.; Cohen, A.J.; Engelsberg, B.N.; Howard, P.S.; Rosenbloom, J. The transforming growth factor-beta-inducible matrix protein (beta)ig-h3 interacts with fibronectin. J. Biol. Chem 2002, 277, 28003–28009. [Google Scholar]
- Hanssen, E.; Reinboth, B.; Gibson, M.A. Covalent and non-covalent interactions of betaig-h3 with collagen VI. Beta ig-h3 is covalently attached to the amino-terminal region of collagen VI in tissue microfibrils. J. Biol. Chem 2003, 278, 24334–24341. [Google Scholar]
- Reinboth, B.; Thomas, J.; Hanssen, E.; Gibson, M.A. Beta ig-h3 interacts directly with biglycan and decorin, promotes collagen VI aggregation, and participates in ternary complexing with these macromolecules. J. Biol. Chem 2006, 281, 7816–7824. [Google Scholar]
- Kim, J.E.; Jeong, H.W.; Nam, J.O.; Lee, B.H.; Choi, J.Y.; Park, R.W.; Park, J.Y.; Kim, I.S. Identification of motifs in the fasciclin domains of the transforming growth factor-beta-induced matrix protein betaig-h3 that interact with the αvβ5 integrin. J. Biol. Chem 2002, 277, 46159–46165. [Google Scholar]
- Ohno, S.; Noshiro, M.; Makihira, S.; Kawamoto, T.; Shen, M.; Yan, W.; Kawashima-Ohya, Y.; Fujimoto, K.; Tanne, K.; Kato, Y. RGD-CAP (βig-h3) enhances the spreading of chondrocytes and fibroblasts via integrin α1β1. Biochim. Biophys. Acta 1999, 1451, 196–205. [Google Scholar]
- Bae, J.S.; Lee, S.H.; Kim, J.E.; Choi, J.Y.; Park, R.W.; Yong Park, J.; Park, H.S.; Sohn, Y.S.; Lee, D.S.; Bae Lee, E.; et al. Betaig-h3 supports keratinocyte adhesion, migration, and proliferation through α3β1 integrin. Biochem. Biophys. Res. Commun 2002, 294, 940–948. [Google Scholar]
- Jeong, H.W.; Kim, I.S. TGF-beta1 enhances betaig-h3-mediated keratinocyte cell migration through the α3β1 integrin and PI3K. J. Cell. Biochem 2004, 92, 770–780. [Google Scholar]
- Kim, H.J.; Kim, I.S. Transforming growth factor-beta-induced gene product, as a novel ligand of integrin αMβ2, promotes monocytes adhesion, migration and chemotaxis. Int. J. Biochem. Cell Biol 2008, 40, 991–1004. [Google Scholar]
- Park, S.J.; Park, S.; Ahn, H.C.; Kim, I.S.; Lee, B.J. Conformational resemblance between the structures of integrin-activating pentapetides derived from betaig-h3 and RGD peptide analogues in a membrane environment. Peptides 2004, 25, 199–205. [Google Scholar]
- Nam, J.O.; Kim, J.E.; Jeong, H.W.; Lee, S.J.; Lee, B.H.; Choi, J.Y.; Park, R.W.; Park, J.Y.; Kim, I.S. Identification of the αvβ3 integrin-interacting motif of betaig-h3 and its anti-angiogenic effect. J. Biol. Chem 2003, 278, 25902–25909. [Google Scholar]
- Li, C.; Lim, S.W.; Choi, B.S.; Lee, S.H.; Cha, J.H.; Kim, I.S.; Kim, J.; Yang, C.W. Inhibitory effect of pravastatin on transforming growth factor beta1-inducible gene h3 expression in a rat model of chronic cyclosporine nephropathy. Am. J. Nephrol 2005, 25, 611–620. [Google Scholar]
- Lee, S.H.; Bae, J.S.; Park, S.H.; Lee, B.H.; Park, R.W.; Choi, J.Y.; Park, J.Y.; Ha, S.W.; Kim, Y.L.; Kwon, T.H.; et al. Expression of TGF-beta-induced matrix protein betaig-h3 is up-regulated in the diabetic rat kidney and human proximal tubular epithelial cells treated with high glucose. Kidney Int 2003, 64, 1012–1021. [Google Scholar]
- Cha, D.R.; Kim, I.S.; Kang, Y.S.; Han, S.Y.; Han, K.H.; Shin, C.; Ji, Y.H.; Kim, N.H. Urinary concentration of transforming growth factor-beta-inducible gene-h3(beta ig-h3) in patients with Type 2 diabetes mellitus. Diabet. Med 2005, 22, 14–20. [Google Scholar]
- Ha, S.W.; Kim, H.J.; Bae, J.S.; Jeong, G.H.; Chung, S.C.; Kim, J.G.; Park, S.H.; Kim, Y.L.; Kam, S.; Kim, I.S.; et al. Elevation of urinary betaig-h3, transforming growth factor-beta-induced protein in patients with type 2 diabetes and nephropathy. Diabetes Res. Clin. Pract 2004, 65, 167–173. [Google Scholar]
- Han, J.H.; Ha, S.W.; Lee, I.K.; Kim, B.W.; Kim, J.G. High glucose-induced apoptosis in bovine retinal pericytes is associated with transforming growth factor beta and betaIG-H3: BetaIG-H3 induces apoptosis in retinal pericytes by releasing Arg-Gly-Asp peptides. Clin. Exp. Ophthalmol 2010, 38, 620–628. [Google Scholar]
- Wang, Y.; Zhao, S.; Loyd, S.; Groome, L.J. Increased urinary excretion of nephrin, podocalyxin, and betaig-h3 in women with preeclampsia. Am. J. Physiol. Renal. Physiol 2012, 302, F1084–F1089. [Google Scholar]
- Hedegaard, C.J.; Thogersen, I.B.; Enghild, J.J.; Klintworth, G.K.; Moller-Pedersen, T. Transforming growth factor beta induced protein accumulation in granular corneal dystrophy type III (Reis-Bucklers dystrophy). Identification by mass spectrometry in 15 year old two-dimensional protein gels. Mol. Vis 2003, 9, 355–359. [Google Scholar]
- Konishi, M.; Yamada, M.; Nakamura, Y.; Mashima, Y. Immunohistology of kerato-epithelin in corneal stromal dystrophies associated with R124 mutations of the BIGH3 gene. Curr. Eye Res 2000, 21, 891–896. [Google Scholar]
- Korvatska, E.; Munier, F.L.; Chaubert, P.; Wang, M.X.; Mashima, Y.; Yamada, M.; Uffer, S.; Zografos, L.; Schorderet, D.F. On the role of kerato-epithelin in the pathogenesis of 5q31-linked corneal dystrophies. Invest. Ophthalmol. Vis. Sci 1999, 40, 2213–2219. [Google Scholar]
- Streeten, B.W.; Qi, Y.; Klintworth, G.K.; Eagle, R.C., Jr; Strauss, J.A.; Bennett, K. Immunolocalization of beta ig-h3 protein in 5q31-linked corneal dystrophies and normal corneas. Arch. Ophthalmol. 1999, 117, 67–75. [Google Scholar]
- Kannabiran, C.; Klintworth, G.K. TGFBI gene mutations in corneal dystrophies. Hum. Mutat 2006, 27, 615–625. [Google Scholar]
- Shao, G.; Berenguer, J.; Borczuk, A.C.; Powell, C.A.; Hei, T.K.; Zhao, Y. Epigenetic inactivation of Betaig-h3 gene in human cancer cells. Cancer Res 2006, 66, 4566–4573. [Google Scholar]
- Skonier, J.; Bennett, K.; Rothwell, V.; Kosowski, S.; Plowman, G.; Wallace, P.; Edelhoff, S.; Disteche, C.; Neubauer, M.; Marquardt, H.; et al. Beta ig-h3: A transforming growth factor-beta-responsive gene encoding a secreted protein that inhibits cell attachment in vitro and suppresses the growth of CHO cells in nude mice. DNA Cell Biol 1994, 13, 571–584. [Google Scholar]
- Kim, J.E.; Kim, S.J.; Jeong, H.W.; Lee, B.H.; Choi, J.Y.; Park, R.W.; Park, J.Y.; Kim, I.S. RGD peptides released from beta ig-h3, a TGF-beta-induced cell-adhesive molecule, mediate apoptosis. Oncogene 2003, 22, 2045–2053. [Google Scholar]
- Zhao, Y.L.; Piao, C.Q.; Hei, T.K. Overexpression of Betaig-h3 gene downregulates integrin α5β1 and suppresses tumorigenicity in radiation-induced tumorigenic human bronchial epithelial cells. Br. J. Cancer 2002, 86, 1923–1928. [Google Scholar]
- Zhao, Y.L.; Piao, C.Q.; Hei, T.K. Downregulation of Betaig-h3 gene is causally linked to tumorigenic phenotype in asbestos treated immortalized human bronchial epithelial cells. Oncogene 2002, 21, 7471–7477. [Google Scholar]
- Zhao, Y.; Shao, G.; Piao, C.Q.; Berenguer, J.; Hei, T.K. Down-regulation of Betaig-h3 gene is involved in the tumorigenesis in human bronchial epithelial cells induced by heavy-ion radiation. Radiat. Res 2004, 162, 655–659. [Google Scholar]
- Zhao, Y.; El-Gabry, M.; Hei, T.K. Loss of Betaig-h3 protein is frequent in primary lung carcinoma and related to tumorigenic phenotype in lung cancer cells. Mol. Carcinog 2006, 45, 84–92. [Google Scholar]
- Wen, G.; Partridge, M.A.; Li, B.; Hong, M.; Liao, W.; Cheng, S.K.; Zhao, Y.; Calaf, G.M.; Liu, T.; Zhou, J.; et al. TGFBI expression reduces in vitro and in vivo metastatic potential of lung and breast tumor cells. Cancer Lett 2011, 308, 23–32. [Google Scholar]
- Wen, G.; Hong, M.; Li, B.; Liao, W.; Cheng, S.K.; Hu, B.; Calaf, G.M.; Lu, P.; Partridge, M.A.; Tong, J.; et al. Transforming growth factor-beta-induced protein (TGFBI) suppresses mesothelioma progression through the Akt/mTOR pathway. Int. J. Oncol 2011, 39, 1001–1009. [Google Scholar]
- Becker, J.; Volland, S.; Noskova, I.; Schramm, A.; Schweigerer, L.L.; Wilting, J. Keratoepithelin reverts the suppression of tissue factor pathway inhibitor 2 by MYCN in human neuroblastoma: A mechanism to inhibit invasion. Int. J. Oncol 2008, 32, 235–240. [Google Scholar]
- Zamilpa, R.; Rupaimoole, R.; Phelix, C.F.; Somaraki-Cormier, M.; Haskins, W.; Asmis, R.; Lebaron, R.G. C-terminal fragment of transforming growth factor beta-induced protein (TGFBIp) is required for apoptosis in human osteosarcoma cells. Matrix Biol 2009, 28, 347–357. [Google Scholar]
- Zhang, Y.; Wen, G.; Shao, G.; Wang, C.; Lin, C.; Fang, H.; Balajee, A.S.; Bhagat, G.; Hei, T.K.; Zhao, Y. TGFBI deficiency predisposes mice to spontaneous tumor development. Cancer Res 2009, 69, 37–44. [Google Scholar]
- Kang, S.; Dong, S.M.; Park, N.H. Frequent promoter hypermethylation of TGFBI in epithelial ovarian cancer. Gynecol. Oncol 2010, 118, 58–63. [Google Scholar]
- Wang, N.; Zhang, H.; Yao, Q.; Wang, Y.; Dai, S.; Yang, X. TGFBI promoter hypermethylation correlating with paclitaxel chemoresistance in ovarian cancer. J. Exp. Clin. Cancer Res 2012, 31. [Google Scholar] [CrossRef]
- Ween, M.P.; Lokman, N.A.; Hoffmann, P.; Rodgers, R.J.; Ricciardelli, C.; Oehler, M.K. Transforming growth factor-beta-induced protein secreted by peritoneal cells increases the metastatic potential of ovarian cancer cells. Int. J. Cancer 2011, 128, 1570–1584. [Google Scholar]
- Calaf, G.M.; Echiburu-Chau, C.; Zhao, Y.L.; Hei, T.K. BigH3 protein expression as a marker for breast cancer. Int. J. Mol. Med 2008, 21, 561–568. [Google Scholar]
- Ivanov, S.V.; Ivanova, A.V.; Salnikow, K.; Timofeeva, O.; Subramaniam, M.; Lerman, M.I. Two novel VHL targets, TGFBI (BIGH3) and its transactivator KLF10, are up-regulated in renal clear cell carcinoma and other tumors. Biochem. Biophys. Res. Commun 2008, 370, 536–540. [Google Scholar]
- Yamanaka, M.; Kimura, F.; Kagata, Y.; Kondoh, N.; Asano, T.; Yamamoto, M.; Hayakawa, M. BIGH3 is overexpressed in clear cell renal cell carcinoma. Oncol. Rep 2008, 19, 865–874. [Google Scholar]
- Hourihan, R.N.; O’Sullivan, G.C.; Morgan, J.G. Transcriptional gene expression profiles of oesophageal adenocarcinoma and normal oesophageal tissues. Anticancer Res 2003, 23, 161–165. [Google Scholar]
- Kitahara, O.; Furukawa, Y.; Tanaka, T.; Kihara, C.; Ono, K.; Yanagawa, R.; Nita, M.E.; Takagi, T.; Nakamura, Y.; Tsunoda, T. Alterations of gene expression during colorectal carcinogenesis revealed by cDNA microarrays after laser-capture microdissection of tumor tissues and normal epithelia. Cancer Res 2001, 61, 3544–3549. [Google Scholar]
- Golembieski, W.A.; Rempel, S.A. cDNA array analysis of SPARC-modulated changes in glioma gene expression. J. Neurooncol 2002, 60, 213–226. [Google Scholar]
- Schneider, D.; Kleeff, J.; Berberat, P.O.; Zhu, Z.; Korc, M.; Friess, H.; Buchler, M.W. Induction and expression of betaig-h3 in pancreatic cancer cells. Biochim. Biophys. Acta 2002, 1588, 1–6. [Google Scholar]
- Aitkenhead, M.; Wang, S.J.; Nakatsu, M.N.; Mestas, J.; Heard, C.; Hughes, C.C. Identification of endothelial cell genes expressed in an in vitro model of angiogenesis: Induction of ESM-1, (beta)ig-h3, and NrCAM. Microvasc. Res 2002, 63, 159–171. [Google Scholar]
- Zajchowski, D.A.; Bartholdi, M.F.; Gong, Y.; Webster, L.; Liu, H.L.; Munishkin, A.; Beauheim, C.; Harvey, S.; Ethier, S.P.; Johnson, P.H. Identification of gene expression profiles that predict the aggressive behavior of breast cancer cells. Cancer Res 2001, 61, 5168–5178. [Google Scholar]
- Ma, C.; Rong, Y.; Radiloff, D.R.; Datto, M.B.; Centeno, B.; Bao, S.; Cheng, A.W.; Lin, F.; Jiang, S.; Yeatman, T.J.; et al. Extracellular matrix protein βig-h3/TGFBI promotes metastasis of colon cancer by enhancing cell extravasation. Genes Dev 2008, 22, 308–321. [Google Scholar]
- Kim, M.O.; Yun, S.J.; Kim, I.S.; Sohn, S.; Lee, E.H. Transforming growth factor-beta-inducible gene-h3 (βig-h3) promotes cell adhesion of human astrocytoma cells in vitro: Implication of α6β4 integrin. Neurosci. Lett 2003, 336, 93–96. [Google Scholar]
- Tang, J.; Zhou, H.W.; Jiang, J.L.; Yang, X.M.; Li, Y.; Zhang, H.X.; Chen, Z.N.; Guo, W.P. BetaIg-h3 is involved in the HAb18G/CD147-mediated metastasis process in human hepatoma cells. Exp. Biol. Med 2007, 232, 344–352. [Google Scholar]
- Tang, J.; Wu, Y.M.; Zhao, P.; Jiang, J.L.; Chen, Z.N. Betaig-h3 interacts with α3β1 integrin to promote adhesion and migration of human hepatoma cells. Exp. Biol. Med 2009, 234, 35–39. [Google Scholar]
- Irigoyen, M.; Anso, E.; Salvo, E.; de las Herrerias, J.D.; Martinez-Irujo, J.J.; Rouzaut, A. TGFbeta-induced protein mediates lymphatic endothelial cell adhesion to the extracellular matrix under low oxygen conditions. Cell. Mol. Life Sci 2008, 65, 2244–2255. [Google Scholar]
- Nummela, P.; Lammi, J.; Soikkeli, J.; Saksela, O.; Laakkonen, P.; Holtta, E. Transforming growth factor beta-induced (TGFBI) is an anti-adhesive protein regulating the invasive growth of melanoma cells. Am. J. Pathol 2012, 180, 1663–1674. [Google Scholar]
- Guo, Y.S.; Tang, J.; Chen, B.; Huang, W.; Li, Y.; Cui, H.Y.; Zhang, X.; Wang, S.J.; Chen, Z.N.; Jiang, J.L. Betaig-h3 regulates store-operated Ca2+ entry and promotes the invasion of human hepatocellular carcinoma cells. Cell Biol. Int 2011, 35, 811–817. [Google Scholar]
- Hu, Y.C.; Lam, K.Y.; Law, S.; Wong, J.; Srivastava, G. Profiling of differentially expressed cancer-related genes in esophageal squamous cell carcinoma (ESCC) using human cancer cDNA arrays: Overexpression of oncogene MET correlates with tumor differentiation in ESCC. Clin. Cancer Res 2001, 7, 3519–3525. [Google Scholar]
- Wong, F.H.; Huang, C.Y.; Su, L.J.; Wu, Y.C.; Lin, Y.S.; Hsia, J.Y.; Tsai, H.T.; Lee, S.A.; Lin, C.H.; Tzeng, C.H.; et al. Combination of microarray profiling and protein-protein interaction databases delineates the minimal discriminators as a metastasis network for esophageal squamous cell carcinoma. Int. J. Oncol 2009, 34, 117–128. [Google Scholar]
- Turtoi, A.; Musmeci, D.; Wang, Y.; Dumont, B.; Somja, J.; Bevilacqua, G.; de Pauw, E.; Delvenne, P.; Castronovo, V. Identification of novel accessible proteins bearing diagnostic and therapeutic potential in human pancreatic ductal adenocarcinoma. J. Proteome Res 2011, 10, 4302–4313. [Google Scholar]
- Tomioka, H.; Morita, K.; Hasegawa, S.; Omura, K. Gene expression analysis by cDNA microarray in oral squamous cell carcinoma. J. Oral Pathol. Med 2006, 35, 206–211. [Google Scholar]
- Lin, B.; Madan, A.; Yoon, J.G.; Fang, X.; Yan, X.; Kim, T.K.; Hwang, D.; Hood, L.; Foltz, G. Massively parallel signature sequencing and bioinformatics analysis identifies up-regulation of TGFBI and SOX4 in human glioblastoma. PLoS One 2010, 5, e10210. [Google Scholar]
- Ma, J.; Cui, W.; He, S.M.; Duan, Y.H.; Heng, L.J.; Wang, L.; Gao, G.D. Human U87 astrocytoma cell invasion induced by interaction of βig-h3 with integrin α5β1 involves calpain-2. PLoS One 2012, 7, e37297. [Google Scholar]
- Buckhaults, P.; Rago, C.; St Croix, B.; Romans, K.E.; Saha, S.; Zhang, L.; Vogelstein, B.; Kinzler, K.W. Secreted and cell surface genes expressed in benign and malignant colorectal tumors. Cancer Res 2001, 61, 6996–7001. [Google Scholar]
- Ohno, Y.; Izumi, M.; Tachibana, M.; Kawamura, T.; Yoshioka, K.; Aoyagi, T.; Ohori, M.; Namiki, K.; Sakamoto, N.; Nakagami, Y.; et al. Characterization and gene expression analysis of novel matched primary and metastatic renal cell carcinoma cell lines. Oncol. Rep 2008, 20, 501–509. [Google Scholar]
- Ahmed, A.A.; Mills, A.D.; Ibrahim, A.E.; Temple, J.; Blenkiron, C.; Vias, M.; Massie, C.E.; Iyer, N.G.; McGeoch, A.; Crawford, R.; et al. The extracellular matrix protein TGFBI induces microtubule stabilization and sensitizes ovarian cancers to paclitaxel. Cancer Cell 2007, 12, 514–527. [Google Scholar]
- Tumbarello, D.A.; Temple, J.; Brenton, J.D. Beta3 integrin modulates transforming growth factor beta induced (TGFBI) function and paclitaxel response in ovarian cancer cells. Mol. Cancer 2012, 11. [Google Scholar] [CrossRef]
- Clout, N.J.; Hohenester, E. A model of FAS1 domain 4 of the corneal protein beta(ig)-h3 gives a clearer view on corneal dystrophies. Mol. Vis 2003, 9, 440–448. [Google Scholar]
- Gardner, M.J.; Jones, L.M.; Catterall, J.B.; Turner, G.A. Expression of cell adhesion molecules on ovarian tumour cell lines and mesothelial cells, in relation to ovarian cancer metastasis. Cancer Lett 1995, 91, 229–234. [Google Scholar]
- Witz, C.A.; Takahashi, A.; Montoya-Rodriguez, I.A.; Cho, S.; Schenken, R.S. Expression of the α2β1 and α3β1 integrins at the surface of mesothelial cells: A potential attachment site of endometrial cells. Fertil. Steril 2000, 74, 579–584. [Google Scholar]
- Heyman, L.; Kellouche, S.; Fernandes, J.; Dutoit, S.; Poulain, L.; Carreiras, F. Vitronectin and its receptors partly mediate adhesion of ovarian cancer cells to peritoneal mesothelium in vitro. Tumour Biol 2008, 29, 231–244. [Google Scholar]
- Cannistra, S.A.; Abu-Jawdeh, G.; Niloff, J.; Strobel, T.; Swanson, L.; Andersen, J.; Ottensmeier, C. CD44 variant expression is a common feature of epithelial ovarian cancer: Lack of association with standard prognostic factors. J. Clin. Oncol 1995, 13, 1912–1921. [Google Scholar]
- Strobel, T.; Cannistra, S.A. Beta1-integrins partly mediate binding of ovarian cancer cells to peritoneal mesothelium in vitro. Gynecol. Oncol 1999, 73, 362–367. [Google Scholar]
- Park, Y.H.; Ryu, H.S.; Choi, D.S.; Chang, K.H.; Park, D.W.; Min, C.K. Effects of hepatocyte growth factor on the expression of matrix metalloproteinases and their tissue inhibitors during the endometrial cancer invasion in a three-dimensional coculture. Int. J. Gynecol. Cancer 2003, 13, 53–60. [Google Scholar]
- Walker, G.; MacLeod, K.; Williams, A.R.; Cameron, D.A.; Smyth, J.F.; Langdon, S.P. Estrogen-regulated gene expression predicts response to endocrine therapy in patients with ovarian cancer. Gynecol. Oncol 2007, 106, 461–468. [Google Scholar]
- Irigoyen, M.; Pajares, M.J.; Agorreta, J.; Ponz-Sarvise, M.; Salvo, E.; Lozano, M.D.; Pio, R.; Gil-Bazo, I.; Rouzaut, A. TGFBI expression is associated with a better response to chemotherapy in NSCLC. Mol. Cancer 2010, 9. [Google Scholar] [CrossRef]



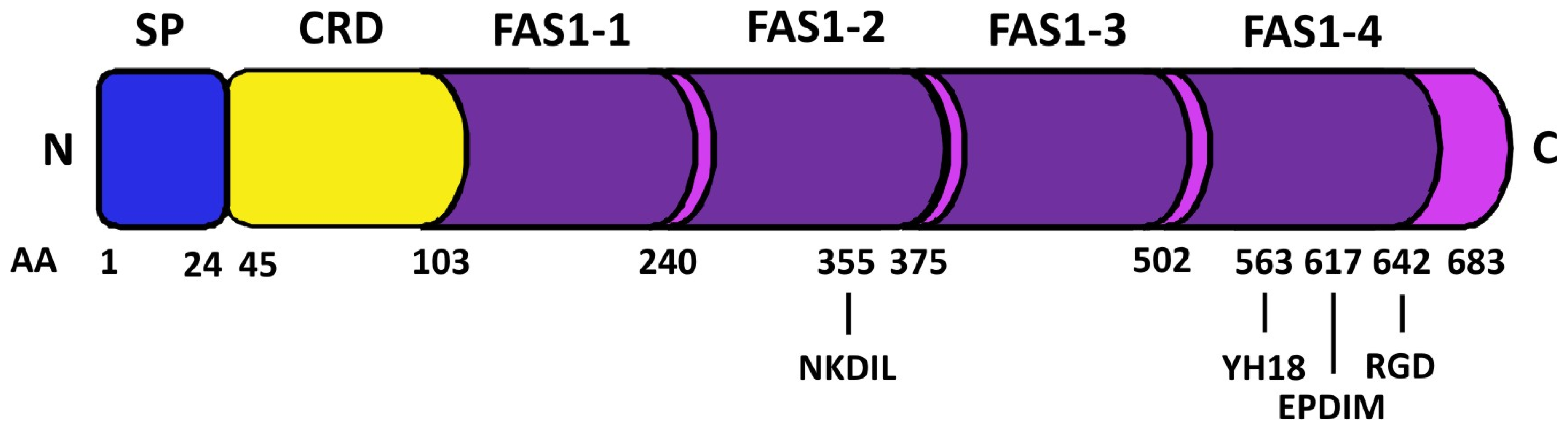{kind=link}
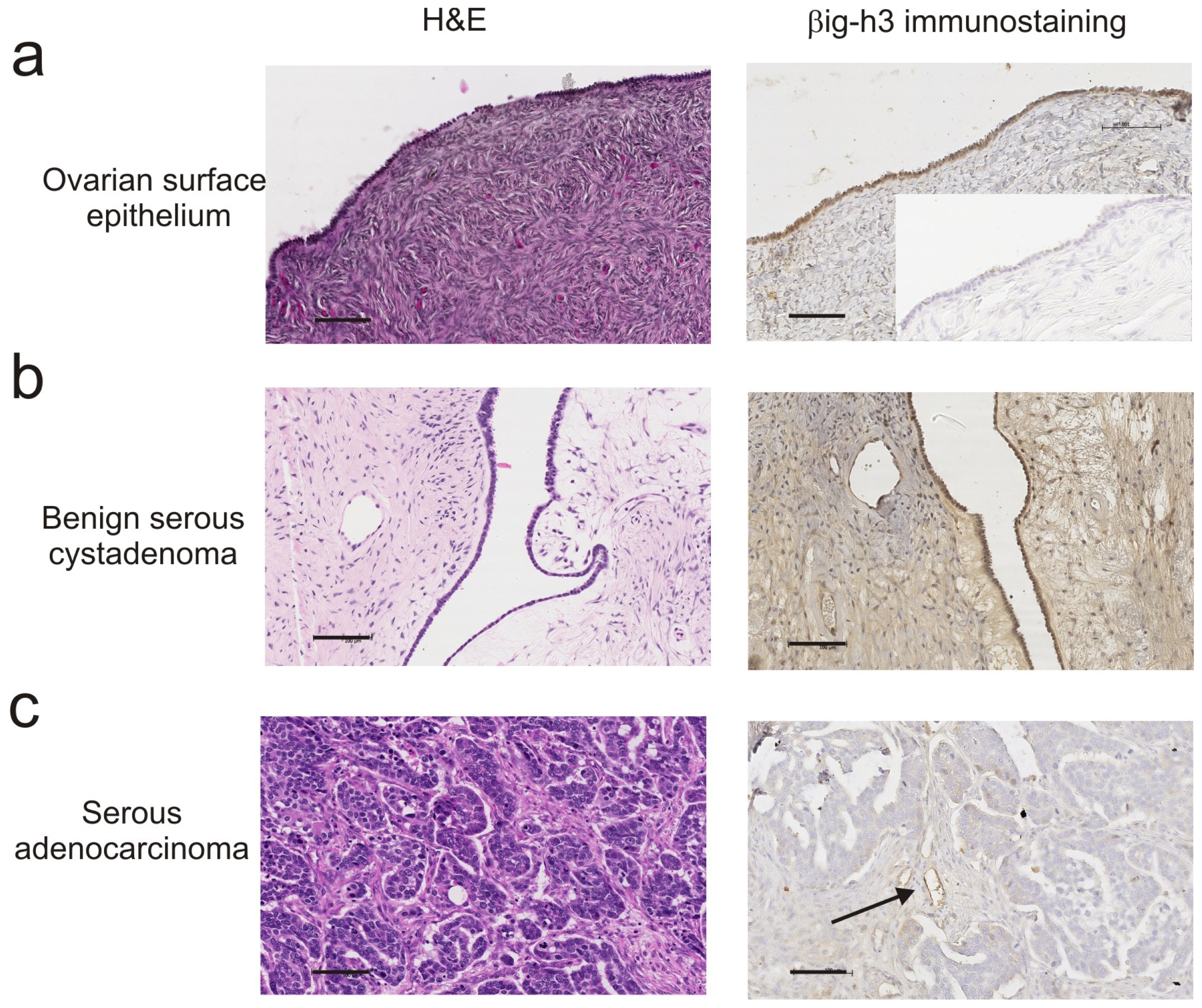{kind=link}
| Cell Type | Observation | References |
|---|---|---|
| CHO cells | βig-H3 inhibits cell attachment in vitro and suppresses the growth of CHO tumor cells in nude mice | [48] |
| RGD peptides released from βig-H3 mediate apoptosis of CHO tumor cells | [49] | |
| HeLa cells | RGD peptides released from βig-H3 mediate apoptosis of HeLa tumor cells | [49] |
| Bronchial epithelial cells | βig-H3 overexpression suppresses tumorigenicity in radiation-induced tumorigenic human bronchial epithelial cells | [50] |
| Loss of βig-H3 expression is associated with the tumorigenic phenotype in asbestos-treated bronchial epithelial cells | [51] | |
| βig-H3 gene down-regulation is involved in heavy-ion radiation-induced tumorigenesis of human bronchial epithelial cells | [52] | |
| Lung adenocarcinoma | Loss of βig-H3 protein is frequent in primary lung carcinoma and related to tumorigenic phenotype in lung cancer cells | [53] |
| Promoter methylation contributes to promoter silencing of the βig-H3 gene in human lung cancer cells | [47] | |
| βig-H3 is down-regulated in radiation-induced thymic lymphoma model in BALB/c mice | [5] | |
| βig-H3 overexpression in H522 lung carcinoma cells reduces motility in vitro and metastasis in vivo | [54] | |
| RGD βig-H3 peptides mediate apoptosis of H1299 lung carcinoma cells | [49] | |
| Mesothelioma cell lines | βig-H3 knockdown increases proliferation and anchorage independent growth of mesothelioma cell lines | [55] |
| Breast carcinoma | βig-H3 protein expression is reduced in in situ ductal carcinoma and breast carcinoma tissues, compared to benign tissues | [54] |
| βig-H3 overexpression in MCF-7 cells reduces motility in vitro and metastasis in vivo | [54] | |
| Neuroblastoma | βig-H3 significantly reduces proliferation and invasion of neuroblastoma cell in vitro and in vivo | [2,56] |
| Osteosarcoma | C-terminal fragment of βig-H3 is required for apoptosis in human osteosarcoma cells | [57] |
| Hepatoma | RGD βig-H3 peptides mediate apoptosis of Hep3B hepatoma cells | [47] |
| Knockout mice | βig-H3 knockout mice are prone to spontaneous tumors | [58] |
| Ovarian carcinoma | βig-H3 silencing and promoter hypermethylation is a frequent occurrence in ovarian cancer cell lines and ovarian cancer tissues | [59,60] |
| βig-H3 is down-regulated in serous ovarian carcinoma and borderline serous ovarian tumors | [61] | |
| βig-H3 induces apotosis in serous ovarian carcinoma cell lines | [61] |
| Cell type | Observation | References |
|---|---|---|
| Lung adenocarcinoma | βig-H3 is overexpressed in lung cancer | [6] |
| Recombinant βig-H3 stimulates proliferation and cell adhesion of A549 cells | [20] | |
| Oesophageal adenocarcinoma | βig-H3 is up-regulated in oesophageal adenocarcinoma and esophageal squamous cell carcinoma tissues and cell lines tissue | [65,78,79] |
| Pancreatic cancer | βig-H3 expression is increased in pancreatic cancer cell lines and tissues | [68,80] |
| Oral squamous cell carcinoma | βig-H3 expression is increased in oral squamous cell carcinoma tissues | [81] |
| Brain tumors | βig-H3 promotes cell adhesion of human astrocytoma cells in vitro via interactions with α6β4 integrin | [72] |
| βig-H3 expression is elevated in glioblastoma multiforme tissues | [82] | |
| Knockdown of βig-H3 inhibits glioma cell invasion and MMP secretion | [83] | |
| Hepatocellular carcinoma | βig-H3 knockdown reduced invasion of 7721 cells | [73] |
| βig-H3 interacts with α3β1 integrin to promote adhesion and invasion of 7721 cells | [74] | |
| Colon carcinoma | βig-H3 expression is elevated in human colon carcinoma tissues | [64,84] |
| Overexpression of βig-H3 promotes extravasation and enhances metastasis of colon cancer cells | [71] | |
| Renal cell carcinoma | βig-H3 is up-regulated in clear cell renal carcinoma | [63,64] |
| βig-H3 expression is increased in metastastic renal cell carcinoma | [85] | |
| Ovarian carcinoma | βig-H3 suppression leads to a chemoresistant phenotype | [86,87] |
| Recombinant βig-H3 promotes motility and invasion of OVCAR-5 and SKOV3 cells | [61] | |
| Recombinant βig-H3 promotes adhesion of OVCAR-3, OVCAR-5 and SKOV3 cells | [61] |
© 2012 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Ween, M.P.; Oehler, M.K.; Ricciardelli, C. Transforming Growth Factor-Beta-Induced Protein (TGFBI)/(βig-H3): A Matrix Protein with Dual Functions in Ovarian Cancer. Int. J. Mol. Sci. 2012, 13, 10461-10477. https://doi.org/10.3390/ijms130810461
Ween MP, Oehler MK, Ricciardelli C. Transforming Growth Factor-Beta-Induced Protein (TGFBI)/(βig-H3): A Matrix Protein with Dual Functions in Ovarian Cancer. International Journal of Molecular Sciences. 2012; 13(8):10461-10477. https://doi.org/10.3390/ijms130810461
Chicago/Turabian StyleWeen, Miranda P., Martin K. Oehler, and Carmela Ricciardelli. 2012. "Transforming Growth Factor-Beta-Induced Protein (TGFBI)/(βig-H3): A Matrix Protein with Dual Functions in Ovarian Cancer" International Journal of Molecular Sciences 13, no. 8: 10461-10477. https://doi.org/10.3390/ijms130810461
APA StyleWeen, M. P., Oehler, M. K., & Ricciardelli, C. (2012). Transforming Growth Factor-Beta-Induced Protein (TGFBI)/(βig-H3): A Matrix Protein with Dual Functions in Ovarian Cancer. International Journal of Molecular Sciences, 13(8), 10461-10477. https://doi.org/10.3390/ijms130810461