Characterization of Erysiphe necator-Responsive Genes in Chinese Wild Vitis quinquangularis

Abstract
:1. Introduction
2. Results
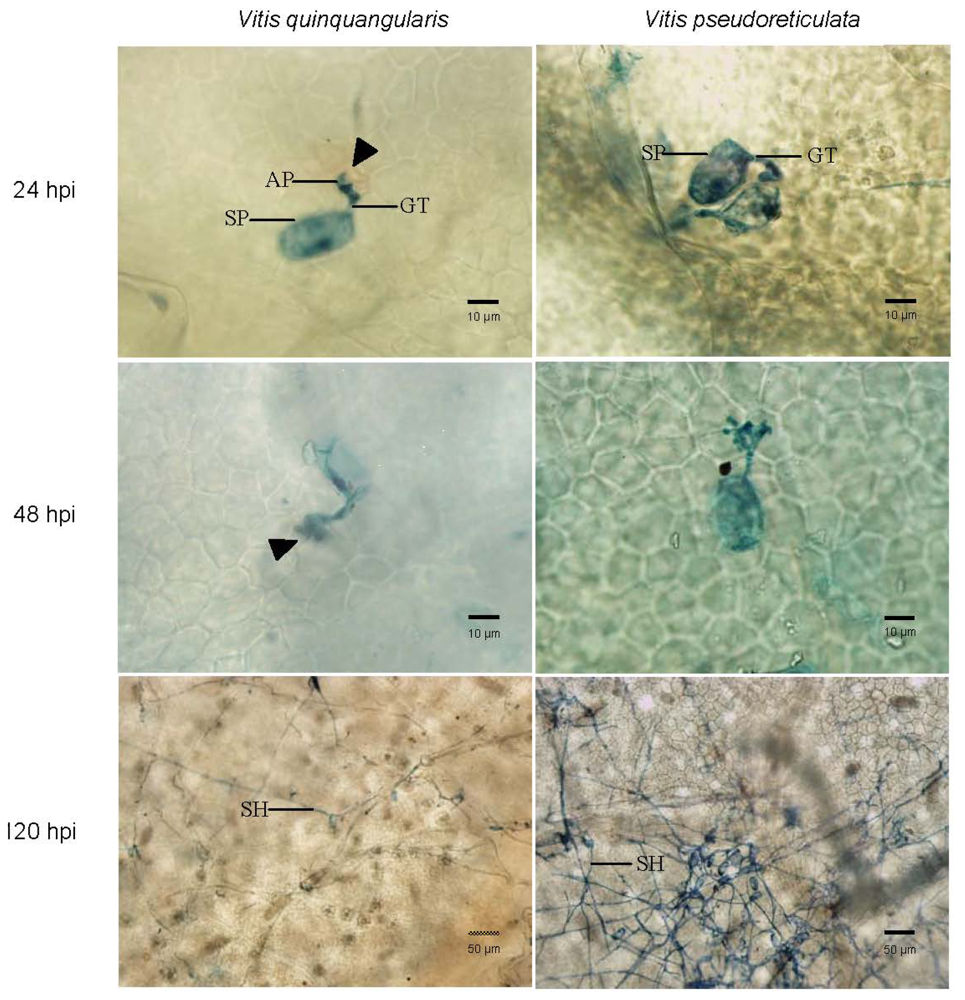
2.1. E. necator Developmental Stages
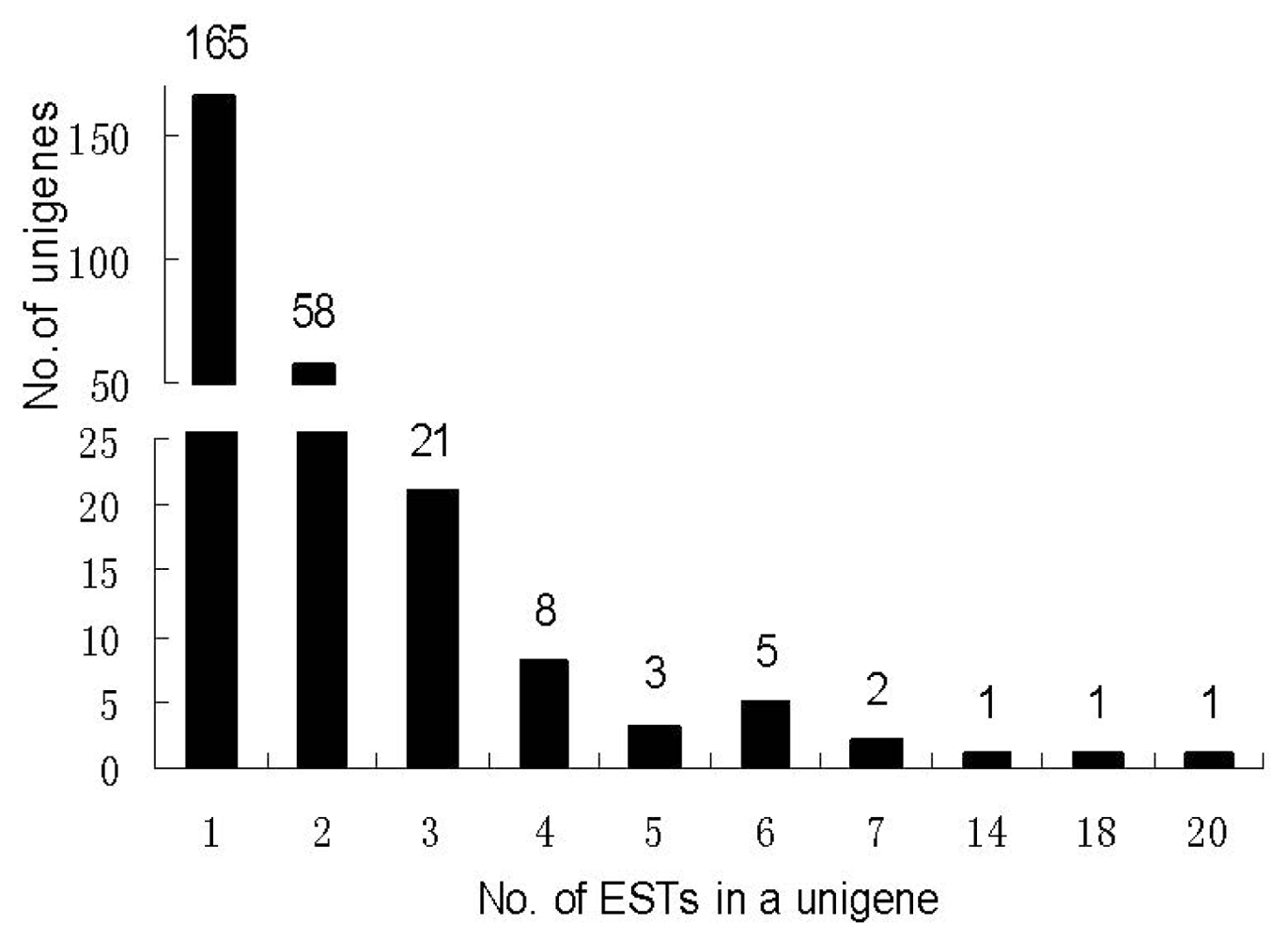
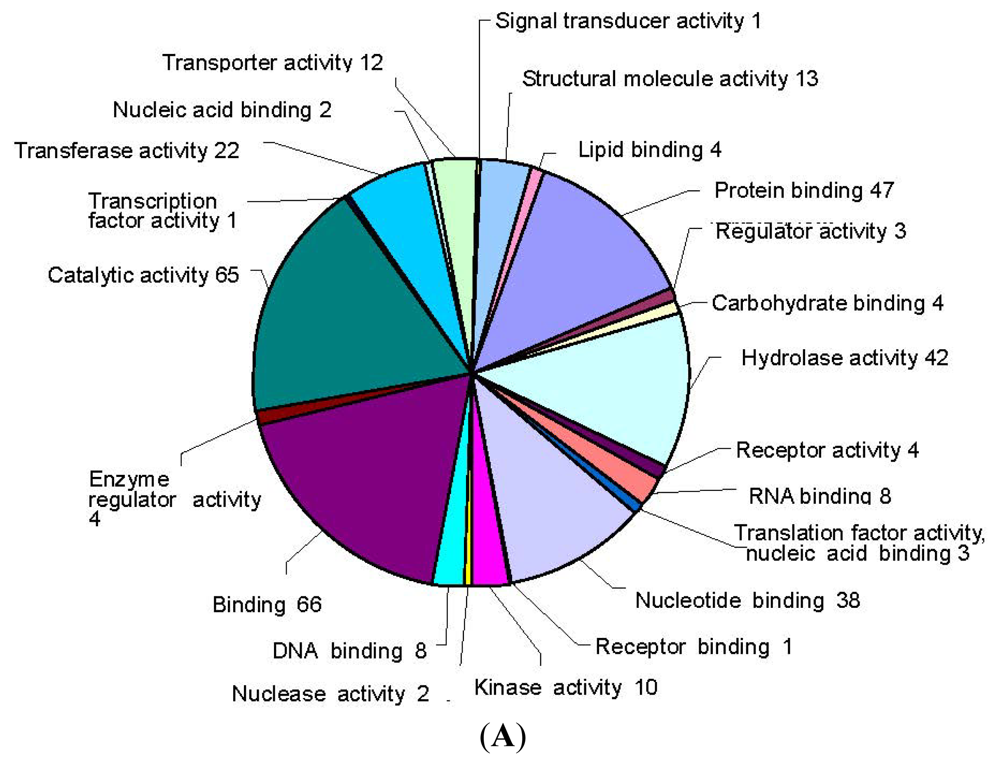
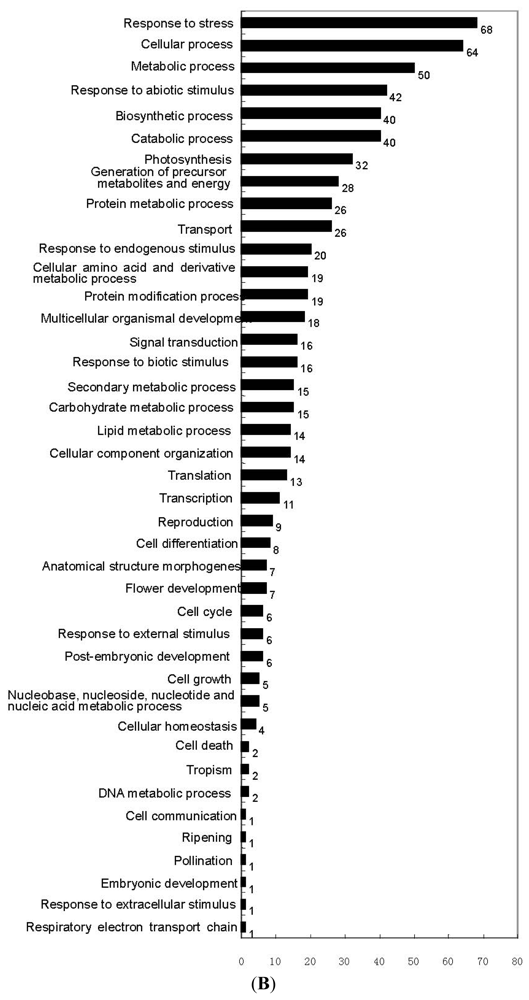
2.2. SSH Library Construction, EST Sequencing, Assembly, and Annotation
2.3. PM Resistance-Related Genes
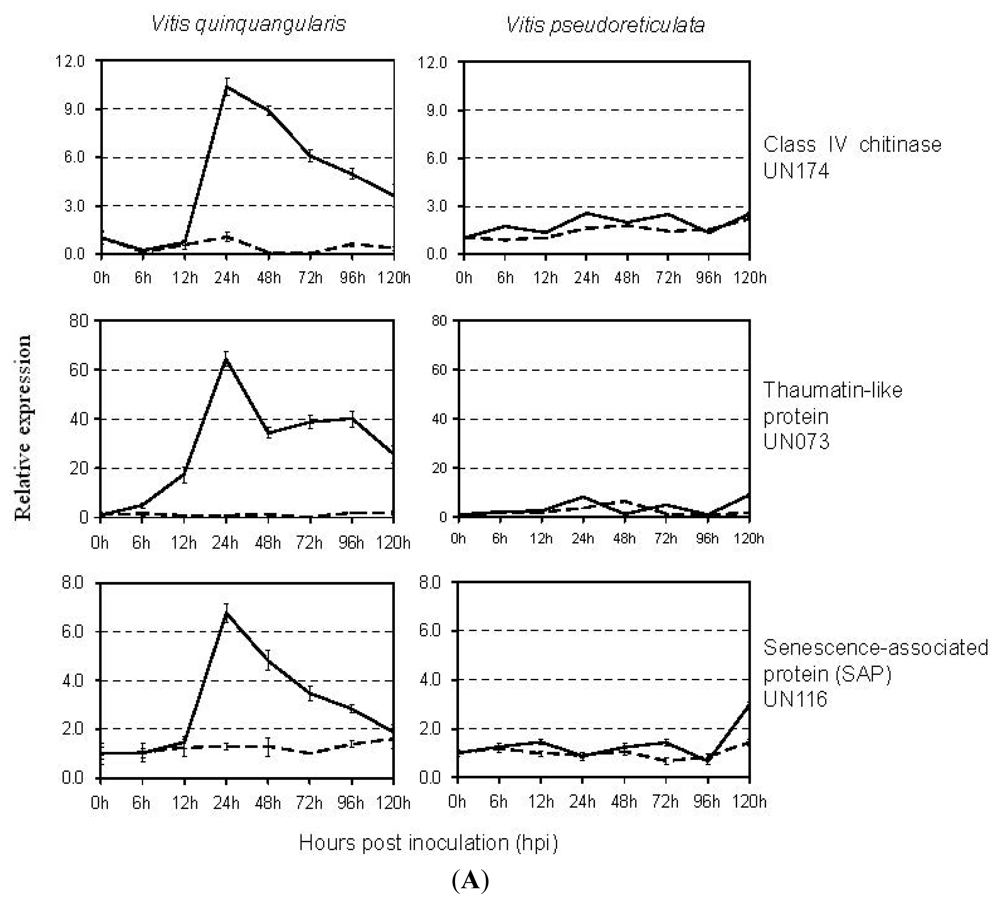
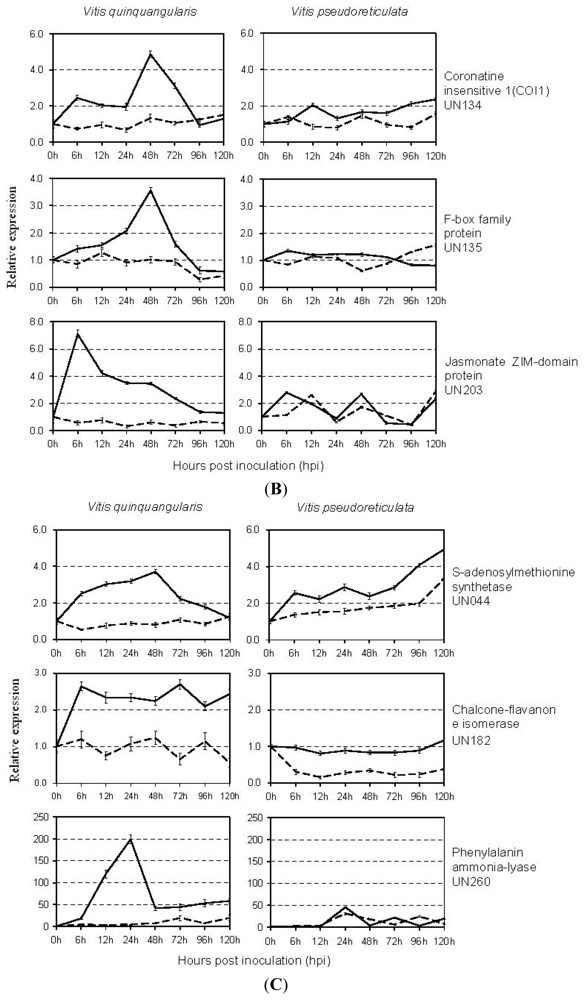
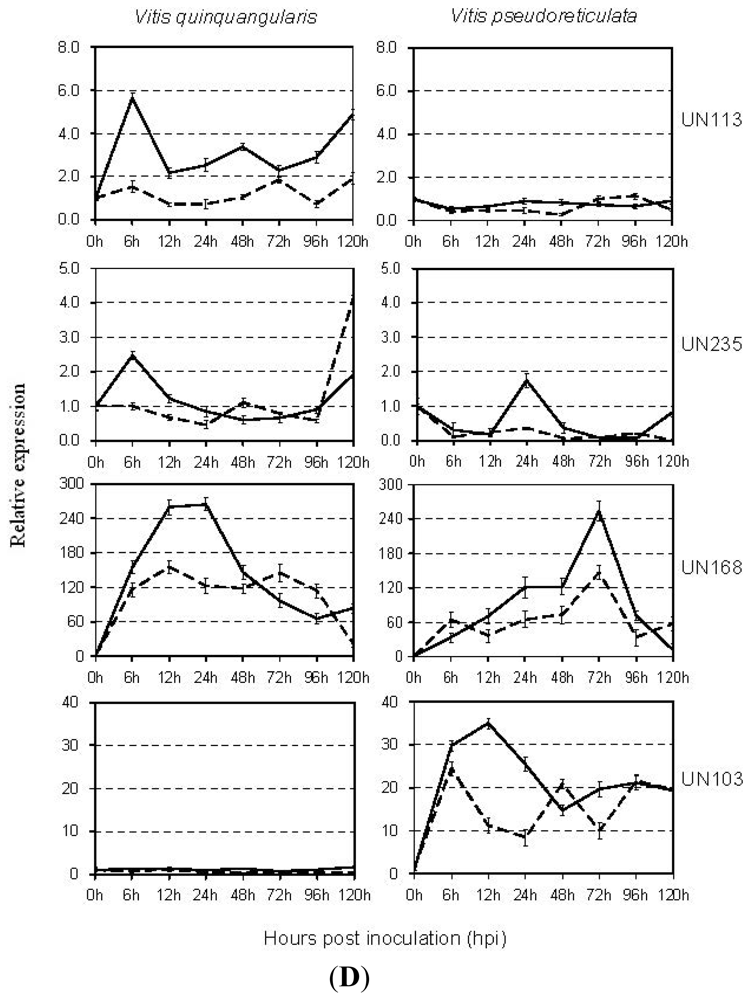
2.4. Expression Analysis of E. necator-Responsive Genes by qRT-PCR
3. Discussion
3.1. Analysis of E. Necator Developmental Stages
3.2. Construction of an SSH Library after E. Necator Inoculation
3.3. Defense-Related Genes
3.4. Transport-Related Genes
3.5. Ubiquitin/26S Proteasome Pathway
3.6. Photosynthesis and Energy-Related Genes
3.7. Metabolism-Associated Genes
3.8. Novel Genes
4. Experimental Section
4.1. Plant Material and Inoculation
4.2. Microscopic Observations
4.3. RNA Extraction and SSH Library Construction
4.4. Amplification of cDNA Inserts
4.5. Sequencing and Data Analysis
4.6. Real Time qRT-PCR Analysis
5. Conclusions
Supplementary Materials
ijms-13-11497-s002.pdfAcknowledgments
Abbreviations
| ABC | ATP-binding cassette |
| CHI | chalcone-flavanone isomerase family protein |
| COI1 | coronatine insensitive 1 |
| EST | expressed sequence tag |
| GO | gene ontology |
| hpi | hour post inoculation |
| JAZ | jasmonate ZIM-domain protein |
| LTP | lipid transfer protein |
| PAL | phenylalanine ammonia-lyase |
| PM | powdery mildew |
| qRT-PCR | quantitative reverse transcriptase polymerase chain reaction |
| SAMS | S-adenosylmethionine synthetase |
| SAR | systemic acquired resistance |
| SSH | suppression subtractive hybridization |
References
- Fekete, C.; Fung, R.W.M.; Szabó, Z.; Qiu, W.; Chang, L.; Schachtman, D.P.; Kovács, L.G. Up-regulated transcripts in a compatible powdery mildew-grapevine interaction. Plant Physiol. Bioch 2009, 47, 732–738. [Google Scholar]
- Donald, T.; Pellerone, F.; Adam-Blondon, A.F.; Bouquet, A.; Thomas, M.; Dry, I. Identification of resistance gene analogs linked to a powdery mildew resistance locus in grapevine. Theor. Appl. Genet 2002, 104, 610–618. [Google Scholar]
- Barker, C.; Donald, T.; Pauquet, J.; Ratnaparkhe, M.; Bouquet, A.; Adam-Blondon, A.F.; Thomas, M.; Dry, I. Genetic and physical mapping of the grapevine powdery mildew resistance gene, Run1, using a bacterial artificial chromosome library. Theor. Appl. Genet 2005, 111, 370–377. [Google Scholar]
- Fung, R.W.M.; Gonzalo, M.; Fekete, C.; Kovacs, L.G.; He, Y.; Marsh, E.; McIntyre, L.M.; Schachtman, D.P.; Qiu, W. Powdery mildew induces defense-oriented reprogramming of the transcriptome in a susceptible but not in a resistant grapevine. Plant Physiol 2008, 146, 236–249. [Google Scholar]
- Winterhagen, P.; Howard, S.F.; Qiu, W.; Kovács, L.G. Transcriptional up-regulation of grapevine MLO genes in response to powdery mildew infection. Am. J. Enol. Vitic 2008, 59, 159–168. [Google Scholar]
- He, P.C.; Wang, Y.J.; Wang, G.Y. Study of the resistance disease in Chinese native wild Vitis L. species. Sci. Agric. Sin 1991, 24, 50–56. [Google Scholar]
- Wang, Y.; Liu, Y.; He, P.; Chen, J.; Lamikanra, O.; Lu, J. Evaluation of foliar resistance to uncinula necator in Chinese wild vitis species. Vitis 1995, 34, 159–164. [Google Scholar]
- Legay, G.; Marouf, E.; Berger, D.; Neuhaus, J.M.; Mauch-Mani, B.; Slaughter, A. Identification of genes expressed during the compatible interaction of grapevine with Plasmopara viticola through suppression subtractive hybridization (SSH). Eur. J. Plant Pathol 2011, 129, 281–301. [Google Scholar]
- Norelli, J.L.; Farrell, R.E.; Bassett, C.L.; Baldo, A.M.; Lalli, D.A.; Aldwinckle, H.S.; Wisniewski, M.E. Rapid transcriptional response of apple to fire blight disease revealed by cDNA suppression subtractive hybridization analysis. Tree Genet. Genomes 2009, 5, 27–40. [Google Scholar]
- Qi, X.; Xu, Q.; Shen, L.; Alfandi, M.; Luo, J.; Chen, X. Identification of differentially expressed genes between powdery mildew resistant near-isogenic line and susceptible line of cucumber by suppression subtractive hybridization. Sci. Hortic 2010, 126, 27–32. [Google Scholar]
- Yu, X.; Wang, X.; Wang, C.; Chen, X.; Qu, Z.; Han, Q.; Zhao, J.; Guo, J.; Huang, L. Wheat defense genes in fungal (Puccinia striiformis) infection. Funct. Integr. Genomics 2010, 10, 227–239. [Google Scholar]
- Vanacker, H.; Carver, T.L.W.; Foyer, C.H. Early H2O2 accumulation in mesophyll cells leads to induction of glutathione during the hyper-sensitive response in the barley-powdery mildew interaction. Plant Physiol 2000, 123, 1289–1300. [Google Scholar]
- Polesani, M.; Bortesi, L.; Ferrarini, A.; Zamboni, A.; Fasoli, M.; Zadra, C.; Lovato, A.; Pezzotti, M.; Delledonne, M.; Polverari, A. General and species-specific transcriptional responses to downy mildew infection in a susceptible (Vitis vinifera) and a resistant (V. riparia) grapevine species. BMC Genomics 2010, 11, 117. [Google Scholar]
- Jiang, H.; Song, W.; Li, A.; Yang, X.; Sun, D. Identification of genes differentially expressed in cauliflower associated with resistance to Xanthomonas campestris pv. campestris. Mol. Biol. Rep 2011, 38, 621–629. [Google Scholar]
- Van Loon, L.; Rep, M.; Pieterse, C. Significance of inducible defense-related proteins in infected plants. Annu. Rev. Phytopathol 2006, 44, 135–162. [Google Scholar]
- Sels, J.; Mathys, J.; de Coninck, B.; Cammue, B.; de Bolle, M.F.C. Plant pathogenesis-related (PR) proteins: A focus on PR peptides. Plant Physiol. Biochem 2008, 46, 941–950. [Google Scholar]
- De A Gerhardt, L.B.; Sachetto-Martins, G.; Contarini, M.G.; Sandroni, M.; de P Ferreira, R.; de Lima, V.M.; Cordeiro, M.C.; de Oliveira, D.E.; Margis-Pinheiro, M. Arabidopsis thaliana class IV chitinase is early induced during the interaction with Xanthomonas campestris. FEBS Lett 1997, 419, 69–75. [Google Scholar]
- Monteiro, S.; Barakat, M.; Piçarra-Pereira, M.A.; Teixeira, A.R.; Ferreira, R.B. Osmotin and thaumatin from grape: A putative general defense mechanism against pathogenic fungi. Phytopathology 2003, 93, 1505–1512. [Google Scholar]
- Punja, Z.K. Transgenic carrots expressing a thaumatin-like protein display enhanced resistance to several fungal pathogens. Can. J. Plant Pathol 2005, 27, 291–296. [Google Scholar]
- Ho, V.S.M.; Wong, J.H.; Ng, T. A thaumatin-like antifungal protein from the emperor banana. Peptides 2007, 28, 760–766. [Google Scholar]
- Kim, B.G.; Fukumoto, T.; Tatano, S.; Gomi, K.; Ohtani, K.; Tada, Y.; Akimitsu, K. Molecular cloning and characterization of a thaumatin-like protein-encoding cDNA from rough lemon. Physiol. Mol. Plant Pathol 2009, 74, 3–10. [Google Scholar]
- Kortekamp, A. Expression analysis of defence-related genes in grapevine leaves after inoculation with a host and a non-host pathogen. Plant Physiol. Biochem 2006, 44, 58–67. [Google Scholar]
- McGee, J.D.; Hamer, J.E.; Hodges, T.K. Characterization of a PR-10 pathogenesis-related gene family induced in rice during infection with Magnaporthe grisea. Mol. Plant Microbe Interact 2001, 14, 877–886. [Google Scholar]
- Vieira, F.A.; Carvalho, A.O.; Vitória, Â.P.; Retamal, C.A.; Gomes, V.M. Differential expression of defence-related proteins in Vigna unguiculata (L. Walp.) seedlings after infection with Fusarium oxysporum. Crop Prot. 2010, 29, 440–447. [Google Scholar]
- Yazaki, K. ABC transporters involved in the transport of plant secondary metabolites. FEBS Lett 2006, 580, 1183–1191. [Google Scholar]
- Kobae, Y.; Sekino, T.; Yoshioka, H.; Nakagawa, T.; Martinoia, E.; Maeshima, M. Loss of AtPDR8, a plasma membrane ABC transporter of Arabidopsis thaliana, causes hypersensitive cell death upon pathogen infection. Plant Cell Physiol 2006, 47, 309–318. [Google Scholar]
- Devoto, A.; Muskett, P.R.; Shirasu, K. Role of ubiquitination in the regulation of plant defence against pathogens. Curr. Opin. Plant Biol 2003, 6, 307–311. [Google Scholar]
- Chung, H.S. Role of JAZ Proteins in the Regulation of Jasmonate Signaling in Arabidopsis; Michigan State University: East Lansing, MI, USA, 2010. [Google Scholar]
- Chung, H.S.; Koo, A.J.K.; Gao, X.; Jayanty, S.; Thines, B.; Jones, A.D.; Howe, G.A. Regulation and function of Arabidopsis JASMONATE ZIM-domain genes in response to wounding and herbivory. Plant Physiol 2008, 146, 952–964. [Google Scholar]
- Li, J.W.; Liu, J.; Zhang, H.; Xie, C.H. Identification and transcriptional profiling of differentially expressed genes associated with resistance to Pseudoperonospora cubensis in cucumber. Plant Cell Rep 2011, 30, 345–357. [Google Scholar]
- Scharte, J.; Schön, H.; Weis, E. Photosynthesis and carbohydrate metabolism in tobacco leaves during an incompatible interaction with Phytophthora nicotianae. Plant Cell Environ 2005, 28, 1421–1435. [Google Scholar]
- Akagi, A.; Dandekar, A.M.; Stotz, H.U. Resistance of Malus domestica fruit to Botrytis cinerea depends on endogenous ethylene biosynthesis. Phytopathology 2011, 101, 1311–1321. [Google Scholar]
- Moschou, P.N.; Sarris, P.F.; Skandalis, N.; Andriopoulou, A.H.; Paschalidis, K.A.; Panopoulos, N.J.; Roubelakis-Angelakis, K.A. Engineered polyamine catabolism preinduces tolerance of tobacco to bacteria and oomycetes. Plant Physiol 2009, 149, 1970–1981. [Google Scholar]
- Jeandet, P.; Douillet-Breuil, A.C.; Bessis, R.; Debord, S.; Sbaghi, M.; Adrian, M. Phytoalexins from the Vitaceae: Biosynthesis, phytoalexin gene expression in transgenic plants, antifungal activity, and metabolism. J. Agric. Food Chem 2002, 50, 2731–2741. [Google Scholar]
- Cools, H.; Ishii, H. Pre-treatment of cucumber plants with acibenzolar-S-methyl systemically primes a phenylalanine ammonia lyase gene (PAL1) for enhanced expression upon attack with a pathogenic fungus. Physiol. Mol. Plant Pathol 2002, 61, 273–280. [Google Scholar]
- Geetha, N.; Amruthesh, K.; Sharathchandra, R.; Shetty, H. Resistance to downy mildew in pearl millet is associated with increased phenylalanine ammonia lyase activity. Funct. Plant Biol 2005, 32, 267–275. [Google Scholar]
- Zhang, J.; Wang, Y.; Wang, X.; Yang, K.; Yang, J. An improved method for rapidly extracting total RNA from Vitis. J. Fruit Sci 2003, 20, 178–181. [Google Scholar]
- Ewing, B.; Hillier, L.D.; Wendl, M.C.; Green, P. Base-calling of automated sequencer traces using Phred. I. Accuracy assessment. Genome Res 1998, 8, 175–185. [Google Scholar]
- Chou, H.H.; Holmes, M.H. DNA sequence quality trimming and vector removal. Bioinformatics 2001, 17, 1093–1104. [Google Scholar]
- Zheng, Y.; Zhao, L.; Gao, J.; Fei, Z. iAssembler: A package for de novo assembly of Roche-454/Sanger transcriptome sequences. BMC Bioinform 2011, 12, 453. [Google Scholar]
- Apweiler, R.; Martin, M.; O’Donovan, C.; Magrane, M.; Alam-Faruque, Y.; Antunes, R.; Barrell, D.; Bely, B.; Bingley, M.; Binns, D. The universal protein resource (UniProt) in 2010. Nucleic Acids Res 2010, 38, 142–148. [Google Scholar]
- Finn, R.D.; Mistry, J.; Tate, J.; Coggill, P.; Heger, A.; Pollington, J.E.; Gavin, O.L.; Gunasekaran, P.; Ceric, G.; Forslund, K. The Pfam protein families database. Nucleic Acids Res 2010, 38, 211–222. [Google Scholar]
- Eddy, S.R. Profile hidden Markov models. Bioinformatics 1998, 14, 755–763. [Google Scholar]
- GO Slim and Subset Guide. Available online: http://www.geneontology.org/GO.slims.shtml accessed on 15 May 2012.








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Unigene No. | Length (bp) | Putative Function | Organism of Best Homology | Accession No. | E-Value |
|---|---|---|---|---|---|
| Metabolism (19) | |||||
| UN018 | 706 | Abscisic stress ripening protein | V. pseudoreticulata | ABC86744 | 5 × 10−21 |
| UN022 | 548 | GDP-d-glucose phosphorylase | A. thaliana | NP_200323 | 2 × 10−19 |
| UN044 a | 562 | S-adenosylmethionine synthetase | R. communis | XP_002512570 | 4 × 10−101 |
| UN102 | 369 | ATP-dependent peptidase/ATPase/metallopeptidase | A. thaliana | NP_564563 | 3 × 10−59 |
| UN127 | 829 | Malate dehydrogenase | N. tabacum | CAC12826 | 3 × 10−139 |
| UN145 | 540 | Putative cinnamoyl-CoA reductase | R. communis | XP_002533639 | 4 × 10−19 |
| UN173 | 250 | ATP-dependent peptidase/ATPase/metallopeptidase | A. thaliana | NP_568604 | 9 × 10−6 |
| UN178 | 592 | GDSL-motif lipase/hydrolase family protein | A. thaliana | NP_190416 | 7 × 10−59 |
| UN183 | 452 | Malate dehydrogenase | V. vinifera | AAF69802 | 6 × 10−75 |
| UN230 | 247 | Aldo/keto reductase, putative | R. communis | XP_002535506 | 8 × 10−18 |
| UN240 | 416 | S-adenosylmethionine-dependent methyltransferase | A. thaliana | NP_173458 | 2 × 10−63 |
| UN262 | 443 | HMGB3 (High mobility group B 3) | A. thaliana | NP_001031075 | 3 × 10−27 |
| UN266 | 558 | NADH dehydrogenase subunit 2 | A. thaliana | NP_085584 | 5 × 10−16 |
| UN035 | 395 | Naringenin-chalcone synthase | Humulus lupulus | CAK19317 | 2 × 10−45 |
| UN040 | 645 | Cinnamate-4-hydroxylase | Canarium album | ACR10242 | 2 × 10−99 |
| UN118 | 204 | Flavonol synthase | V. vinifera | BAE75808 | 1 × 10−26 |
| UN166 | 333 | Phenylalanine ammonia-lyase | Camellia oleifera | ACT21093 | 8 × 10−50 |
| UN182 a | 616 | Chalcone-flavanone isomerase | A. thaliana | NP_568154 | 8 × 10−41 |
| UN260 a | 307 | Phenylalanin ammonia-lyase | V. vinifera | ABM67591 | 3 × 10−25 |
| Photosynthesis and Energy (15) | |||||
| UN001 | 618 | Ribulose-1,5-bisphosphate carboxylase/oxygenase activase 1 | Acer rubrum | ABI94077 | 2 × 10−108 |
| UN017 | 562 | Ribulose bisphosphate carboxylase/oxygenase activase 1 | R. communis | XP_002532996 | 2 × 10−89 |
| UN020 | 455 | Photosystem I reaction center subunit II, chloroplast precursor | R. communis | XP_002516772 | 4 × 10−56 |
| UN023 | 624 | Photosystem II 22 kDa protein, chloroplast precursor | R. communis | XP_002513761 | 1 × 10−68 |
| UN025 | 438 | Ribulose bisphosphate carboxylase/oxygenase activase 1 | R. communis | XP_002532996 | 3 × 10−73 |
| UN052 | 365 | Chloroplast photosystem II 10 kDa polypeptide | Jatropha curcas | ADB93062 | 4 × 10−41 |
| UN086 | 255 | Photosystem I reaction center subunit II, chloroplast precursor | R. communis | XP_002516772 | 3 × 10−17 |
| UN093 | 298 | Putative fructose-1,6-bisphosphatase | R. communis | XP_002527886 | 3 × 10−28 |
| UN095 | 348 | Ribulose-1,5-bisphophate carboxylase/oxygenase activase 1 | V. pseudoreticulata | ABC86738 | 5 × 10−47 |
| UN098 | 571 | Putative fructose-bisphosphate aldolase | R. communis | XP_002512993 | 2 × 10−100 |
| UN106 | 268 | Ribulose-1,5-bisphosphate carboxylase/oxygenase activase 1 | Gossypium hirsutum | ABB20913 | 2 × 10−20 |
| UN121 | 371 | Chlorophyll A/B binding protein | R. communis | XP_002533251 | 1 × 10−45 |
| UN198 | 312 | Light harvesting chlorophyll a/b-binding protein | Nicotiana sylvestris | BAA25394 | 2 × 10−35 |
| UN241 | 189 | Light-harvesting complex II protein Lhcb1 | Populus trichocarpa | XP_002307725 | 4 × 10−11 |
| UN255 | 324 | Chlorophyll A/B binding protein | R. communis | XP_002519950 | 5 × 10−44 |
| Disease/Defence (22) | |||||
| UN016 | 477 | Stress-induced cysteine proteinase | Lavatera thuringiaca | AAB62937 | 4 × 10−66 |
| UN033 | 427 | Similar to pathogenesis-related protein 10.3 | V. vinifera | XP_002274617 | 1 × 10−51 |
| UN040 | 645 | Cytochrome P450 | Populus trichocarpa | XP_002325637 | 3 × 10−98 |
| UN073 a | 351 | Thaumatin-like protein | V. vinifera | AAQ10092 | 7 × 10−67 |
| UN075 | 346 | Glutathione S-transferase 3 | Papaver somniferum | AAF22519 | 1 × 10−8 |
| UN077 | 450 | Chaperone protein dnaJ | R. communis | XP_002514419 | 3 × 10−68 |
| UN079 | 294 | Responsive to dehydration 19 | A. thaliana | NP_568052 | 2 × 10−43 |
| UN096 | 755 | Lipid transfer protein isoform 4 | V. vinifera | AAO33394 | 3 × 10−47 |
| UN170 | 468 | Lipid transfer protein family protein | Tamarix hispida | ACM78616 | 6 × 10−8 |
| UN205 | 470 | Lipid transfer protein family protein | A. thaliana | NP_680546 | 5 × 10−31 |
| UN226 | 537 | Lipid transfer protein family protein | A. thaliana | NP_565348 | 2 × 10−33 |
| UN115 | 574 | Bacterial-induced peroxidase precursor | G. hirsutum | AAD43561 | 1 × 10−71 |
| UN116 a | 769 | Similar to senescence-associated protein | A. thaliana | BAD42919 | 3 × 10−70 |
| UN119 | 698 | Catalase 1 | N. tabacum | AAB71764 | 2 × 10−80 |
| UN174 a | 443 | Class IV chitinase | N. tabacum | BAF44533 | 3 × 10−41 |
| UN179 | 318 | Proline-rich protein | Solanum tuberosum | CAA04449 | 1 × 10−22 |
| UN209 | 212 | Harpin-induced family protein | A. thaliana | NP_190008 | 1 × 10−22 |
| UN219 | 341 | Responsive to dehydration 21 | A. thaliana | NP_564497 | 2 × 10−44 |
| UN233 | 465 | Proline-rich protein | G. hirsutum | ABM05952 | 2 × 10−16 |
| UN239 | 240 | Chitinase-like protein | G. hirsutum | AAQ56598 | 2 × 10−35 |
| UN058 | 474 | 2OG-Fe(II) oxygenase family protein | A. thaliana | NP_192787 | 1 × 10−50 |
| UN043 | 581 | Ascorbate peroxidase 3 | A. thaliana | NP_195226 | 4 × 10−67 |
| Protein synthesis and fate (7) | |||||
| UN004 | 372 | 40S ribosomal protein S17 | A. thaliana | NP_196100 | 1 × 10−41 |
| UN010 | 252 | 26S proteasome regulatory subunit | A. thaliana | AAP86657 | 2 × 10−39 |
| UN134 a | 468 | Coronatine insensitive 1 (COI1) | A. thaliana | ABR45948 | 2 × 10−47 |
| UN135 a | 356 | F-box family protein | A. thaliana | NP_199429 | 2 × 10−31 |
| UN203 a | 784 | Jasmonate ZIM-domain protein 2 | N. tabacum | BAG68656 | 5 × 10−43 |
| UN214 | 352 | Putative 40S Ribosomal protein | O. sativa | AAK92638 | 3 × 10−39 |
| UN254 | 229 | Aspartyl protease family protein | A. thaliana | NP_566966 | 3 × 10−15 |
| Signal transduction (8) | |||||
| UN005 | 653 | Putative zinc finger protein | O. sativa | BAD87736 | 1 × 10−24 |
| UN008 | 415 | CIPK2; protein serine/threonine kinase | A. thaliana | NP_196324 | 2 × 10−11 |
| UN031 | 410 | Serine/threonine phosphatase | A. thaliana | NP_188632 | 5 × 10−29 |
| UN151 | 332 | CIPK21; protein serine/threonine kinase | A. thaliana | NP_568860 | 3 × 10−29 |
| UN211 | 636 | Lipid phosphate phosphatase 3 | A. thaliana | NP_001078096 | 2 × 10−35 |
| UN216 | 447 | GTP binding/phospholipase activator | A. thaliana | NP_191788 | 1 × 10−79 |
| UN217 | 508 | Rab-type small GTP-binding protein | Cicer arietinum | BAA76423 | 2 × 10−49 |
| UN238 | 543 | Phosphoribulose kinase, putative | R. communis | XP_002519002 | 3 × 10−82 |
| Transport (12) | |||||
| UN011 | 324 | NRAMP3; transmembrane transporter | A. thaliana | NP_179896 | 6 × 10−45 |
| UN029 | 862 | Plasma membrane aquaporin | V. vinifera | AAF80557 | 2 × 10−148 |
| UN030 | 910 | Antiporter/triose-phosphate transmembrane transporter | A. thaliana | NP_851138 | 1 × 10−144 |
| UN042 | 584 | Putative sorbitol transporter | Prunus cerasus | AAM44082 | 7 × 10−91 |
| UN067 | 647 | Non-specific lipid transfer protein | V. vinifera | ABA29446 | 2 × 10−36 |
| UN088 | 249 | Plasma membrane intrinsic protein | A. thaliana | AAB65787 | 3 × 10−38 |
| UN129 | 401 | ABC transporter family protein | Populus trichocarpa | XP_002331473 | 6 × 10−60 |
| UN136 | 478 | ABC transporter-like | A. thaliana | BAF01914 | 3 × 10−83 |
| UN231 | 565 | ABC transporter family protein | Populus trichocarpa | XP_002308160 | 7 × 10−56 |
| UN164 | 509 | Polyol transporter | Populus trichocarpa | XP_002313809 | 4 × 10−68 |
| UN223 | 269 | Nitrate transporter | Malus hupehensis | ACN72639 | 2 × 10−34 |
| UN221 | 329 | Solute symporter family protein | A. thaliana | NP_199351 | 5 × 10−42 |
| Transcription (4) | |||||
| UN014 | 517 | Transcription activator/transcription factor | A. thaliana | NP_197904 | 2 × 10−18 |
| UN064 | 446 | Transcript elongation factor | A. thaliana | NP_181390 | 5 × 10−27 |
| UN065 | 443 | IAA16; transcription factor | A. thaliana | NP_187124 | 3 × 10−46 |
| UN167 | 430 | DNA binding/transcription factor | A. thaliana | NP_172094 | 1 × 10−21 |
© 2012 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Gao, M.; Niu, J.; Zhao, S.; Jiao, C.; Xu, W.; Fei, Z.; Wang, X. Characterization of Erysiphe necator-Responsive Genes in Chinese Wild Vitis quinquangularis. Int. J. Mol. Sci. 2012, 13, 11497-11519. https://doi.org/10.3390/ijms130911497
Gao M, Niu J, Zhao S, Jiao C, Xu W, Fei Z, Wang X. Characterization of Erysiphe necator-Responsive Genes in Chinese Wild Vitis quinquangularis. International Journal of Molecular Sciences. 2012; 13(9):11497-11519. https://doi.org/10.3390/ijms130911497
Chicago/Turabian StyleGao, Min, Jiao Niu, Suping Zhao, Chen Jiao, Weirong Xu, Zhangjun Fei, and Xiping Wang. 2012. "Characterization of Erysiphe necator-Responsive Genes in Chinese Wild Vitis quinquangularis" International Journal of Molecular Sciences 13, no. 9: 11497-11519. https://doi.org/10.3390/ijms130911497
APA StyleGao, M., Niu, J., Zhao, S., Jiao, C., Xu, W., Fei, Z., & Wang, X. (2012). Characterization of Erysiphe necator-Responsive Genes in Chinese Wild Vitis quinquangularis. International Journal of Molecular Sciences, 13(9), 11497-11519. https://doi.org/10.3390/ijms130911497