Resveratrol Down-Regulates Myosin Light Chain Kinase, Induces Apoptosis and Inhibits Diethylnitrosamine-Induced Liver Tumorigenesis in Rats


Abstract
:1. Introduction
2. Results and Discussion
2.1. Body and Liver Weights and Nodule Growth
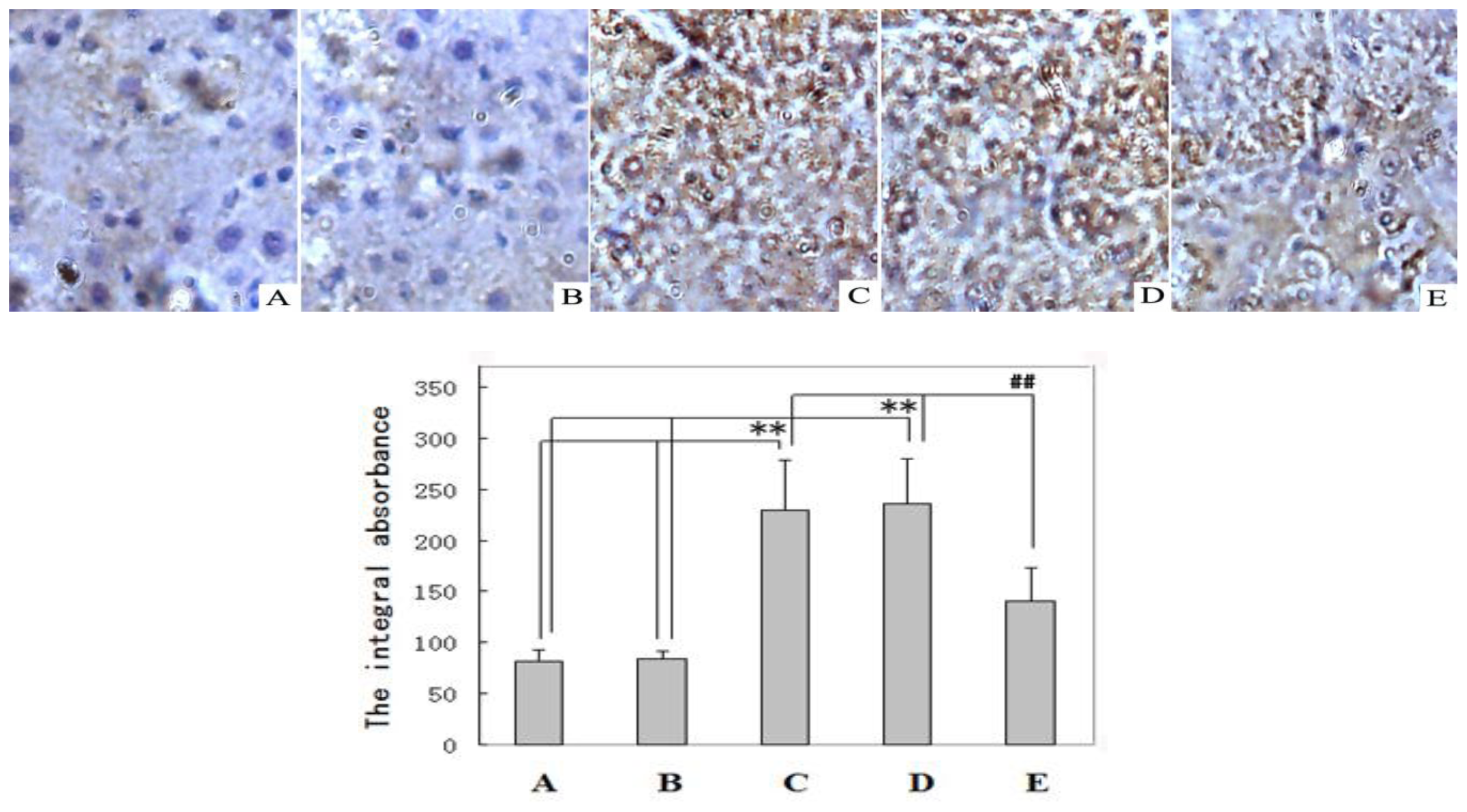
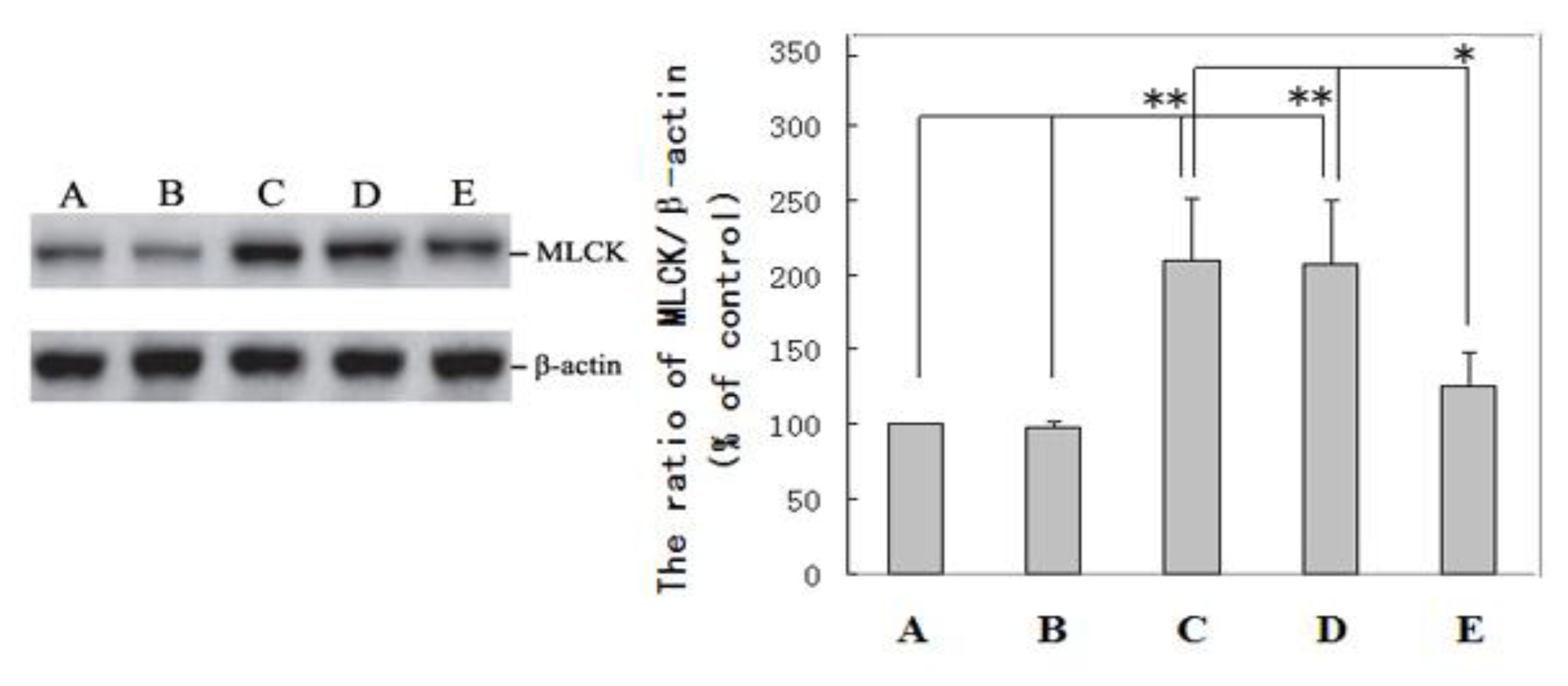
2.2. Immunohistochemical Examination and Western Blot Analysis
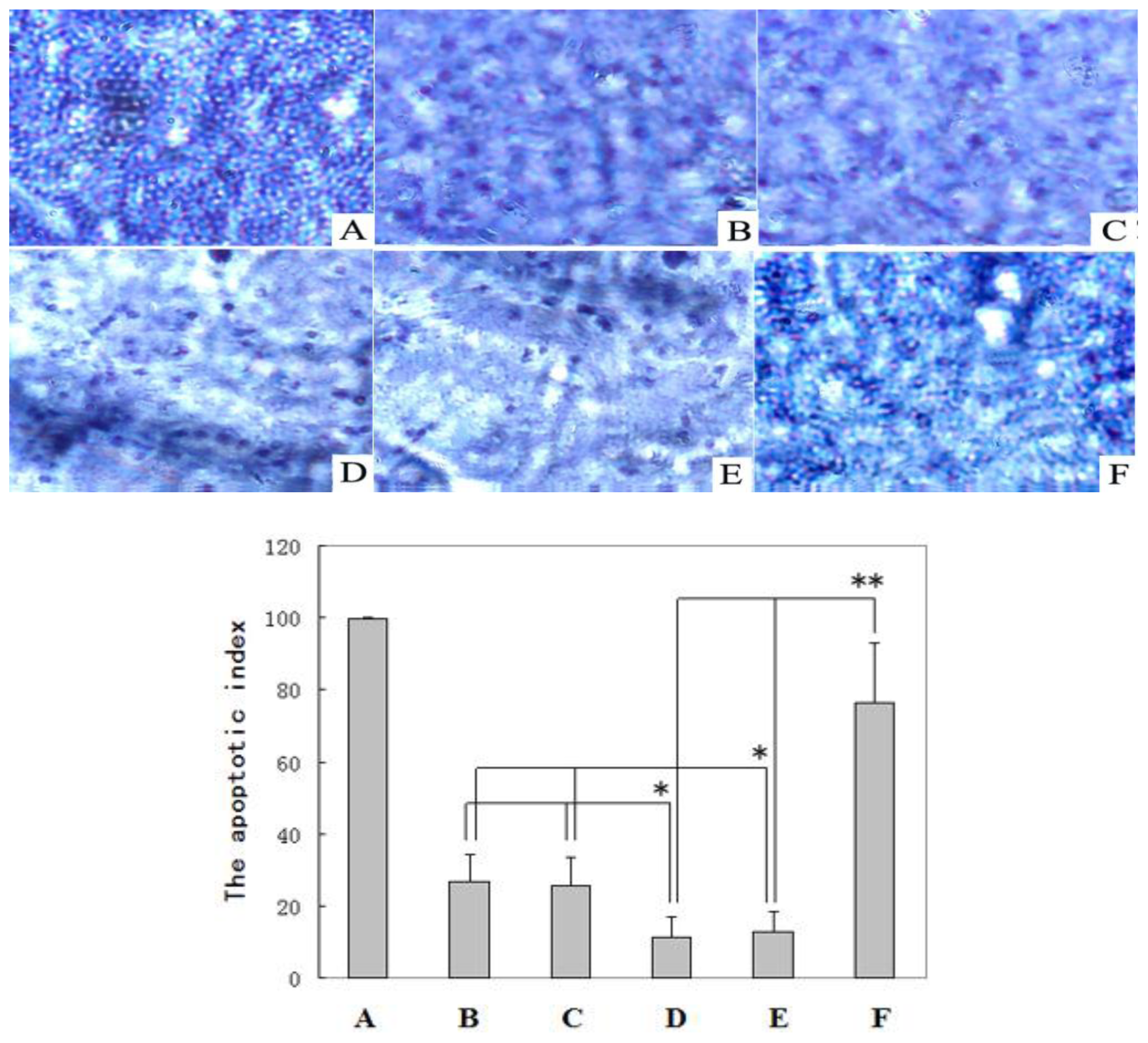
2.3. TUNEL Analysis
3. Experimental Section
3.1. Animals and Chemicals
3.2. Experimental Design
3.3. Tissue Collection and Isolation
3.4. Immunohistochemistry
3.5. Western Blot Analysis
3.6. Detection of in situ Cell Apoptosis by TUNEL Assay
3.7. Statistical Analysis
4. Conclusions
Acknowledgments
- Conflict of InterestThe authors declare no conflicts of interest.
References
- Jemal, A.; Bray, F.; Center, M.M.; Ferlay, J.; Ward, E.; Forman, D. Global cancer statistics. CA Cancer J. Clin 2011, 61, 69–90. [Google Scholar]
- Thorgeirsson, S.S.; Grisham, J.W. Molecular pathogenesis of human hepatocellular carcinoma. Nat. Genet 2002, 31, 339–346. [Google Scholar]
- Bosch, F.X.; Ribes, J.; Diaz, M.; Cleries, R. Primary liver cancer: Worldwide incidence and trends. Gastroenterology 2004, 127, S5–S16. [Google Scholar]
- Farazi, P.A.; DePinho, R.A. Hepatocellular carcinoma pathogenesis: From genes to environment. Nat. Rev. Cancer 2006, 6, 674–687. [Google Scholar]
- Bartsch, H.; Montesano, R. Relevance of nitrosamines to human cancer. Carcinogenesis 1984, 5, 1381–1393. [Google Scholar]
- Kensler, T.W.; Egner, P.A.; Wang, J.B. Chemoprevention of hepatocellular carcinoma in aflatoxin endemic areas. Gastroenterology 2004, 127, S310–S318. [Google Scholar]
- Brechot, C. Pathogenesis of hepatitis B virus-related hepatocellular carcinoma: Old and new paradigms. Gastroenterology 2004, 127, S56–S61. [Google Scholar]
- Yates, M.S.; Kensler, T.W. Keap1 eye on the target: Chemoprevention of liver cancer. Acta Pharmacol. Sin 2007, 28, 1331–1342. [Google Scholar]
- Aziz, M.H.; Kumar, R.; Ahmad, N. Cancer chemoprevention by resveratrol: In vitro and in vivo studies and the underlying mechanisms (review). Int. J. Oncol 2003, 23, 17–28. [Google Scholar]
- Bhat, K.P.; Pezzuto, J.M. Cancer chemopreventive activity of resveratrol. Ann. N. Y. Acad. Sci 2002, 957, 210–229. [Google Scholar]
- Bianchini, F.; Vainio, H. Wine and resveratrol: Mechanisms of cancer prevention? Eur. J. Cancer Prev 2003, 12, 417–425. [Google Scholar]
- Fulda, S. Resveratrol and derivatives for the prevention and treatment of cancer. Drug Discov. Today 2010, 15, 757–765. [Google Scholar]
- Aggarwal, B.B.; Bhardwaj, A.; Aggarwal, R.S. Role of resveratrol in prevention and therapy of cancer: Preclinical and clinical studies. Anticancer Res 2004, 24, 2783–2840. [Google Scholar]
- Bishayee, A.; Dhir, N. Resveratrol-Mediated chemoprevention of diethylnitrosamine-initiated hepatocarcinogenesis: Inhibition of cell proliferation and induction of apoptosis. Chem. Biol. Interact 2009, 179, 131–144. [Google Scholar]
- Bishayee, A.; Barnes, K.F.; Bhatia, D.; Darvesh, A.S.; Carroll, R.T. Resveratrol suppresses oxidative stress and inflammatory response in diethylnitrosamine-initiated rat hepatocarcinogene-sis. Cancer Prev. Res. (Phila) 2010, 3, 753–763. [Google Scholar]
- Bishayee, A.; Waghray, A.; Barnes, K.F.; Mbimba, T.; Bhatia, D.; Chatterjee, M.; Darvesh, A.S. Suppression of the inflammatory cascade is implicated in resveratrol chemoprevention of experimental hepatocarcinogenesis. Pharm. Res 2010, 27, 1080–1091. [Google Scholar]
- Mbimba, T.; Awale, P.; Bhatia, D.; Geldenhuys, W.J.; Darvesh, A.S.; Carroll, R.T.; Bishayee, A. Alteration of hepatic proinflammatory cytokines is involved in the resveratrol-mediated chemoprevention of chemically-induced hepatocarcinogenesis. Curr. Pharm. Biotechnol 2012, 13, 229–234. [Google Scholar]
- Bishayee, A.; Petit, D.M.; Samtani, K. Angioprevention is implicated in resveratrol chemoprevention of experimental hepatocarcinogenesis. J. Carcinog. Mutagen 2010, 1, 1–8. [Google Scholar]
- Staunton, M.J.; Gaffney, E.F. Apoptosis: Basic concepts and potential significance in human cancer. Arch. Pathol. Lab. Med 1998, 122, 310–319. [Google Scholar]
- Samali, A.; Gorman, A.M.; Cotter, T.G. Apoptosis: The story so far…. Expeientia 1996, 52, 933–941. [Google Scholar]
- Grossmann, J. Molecular mechanisms of “detachment-induced apoptosis-anoikis”. Apoptosis 2004, 7, 247–260. [Google Scholar]
- Wu, Y.; Qiao, X.; Qiao, S.; Yu, L. Targeting integrins in hepatocellular carcinoma. Expert. Opin. Ther. Targets 2011, 15, 421–437. [Google Scholar]
- Carpenter, C.L. Actin cytoskeleton and cell signaling. Crit. Care Med 2000, 28, 94–99. [Google Scholar]
- Minamiya, Y.; Nakagawa, T.; Saito, H. Increased expression of myosin light chain kinase mRNA is related to metastasis in non-small cell lung cancer. Tumour Biol 2005, 26, 153–157. [Google Scholar]
- Kaneko, K.; Satoh, K.; Masamune, A.; Satoh, A.; Shimosegawa, T. Myosin light chain kinase inhibitors can block invasion and adhesion of human pancreatic cancer cell lines. Pancreas 2002, 24, 34–41. [Google Scholar]
- Fazal, F.; Gu, L.; Ihnatovych, I. Inhibiting myosin light chain kinase induces apoptosis in vitro and in vivo. Mol. Cell. Biol 2005, 25, 6259–6266. [Google Scholar]
- Bishayee, A.; Chatterjee, M. Inhibitory effect of vanadium on rat liver carcinogenesis initiated with diethylnitrosamine and promoted by Phenobarbital. Br. J. Cancer 1995, 71, 1214–1220. [Google Scholar]
- Sporn, M.B.; Dunlop, N.M.; Newton, D.L.; Smith, J.M. Prevention of chemical carcinogenesis by vitamin A and its synthetic analogs. Fed. Proc 1976, 35, 1332–1338. [Google Scholar]
- Kelloff, G.J.; Boone, C.W.; Steele, V.E.; Fay, J.R.; Lubet, R.A.; Crowell, J.A.; Sigman, C.C. Mechanistic considerations in chemopreventive drug development. J. Cell. Biochem 1994, 20, S1–S24. [Google Scholar]
- Kelloff, G.J.; Boone, C.W.; Steele, V.E.; Crowell, J.A.; Lubet, R.A.; Greenwald, P.; Hawk, E.T.; Fay, J.R.; Sigman, C.C. Mechanistic considerations in the evaluation of chemopreventive data. IARC Sci 1996, 139, 203–219. [Google Scholar]
- Greenwald, P. From carcinogenesis to clinical interventions for cancer prevention. Toxicology 2001, 166, 37–45. [Google Scholar]
- Athar, M.; Back, J.H.; Kopelovich, L.; Bickers, D.R.; Kim, A.L. Multiple molecular targets of resveratrol: Anti-Carcinogenic mechanisms. Arch. Biochem. Biophys 2009, 486, 95–102. [Google Scholar]
- Kundu, J.K.; Surh, Y.J. Cancer chemopreventive and therapeutic potential of resveratrol: Mechanistic perspectives. Cancer Lett 2008, 269, 243–261. [Google Scholar]
- Bishayee, A. Cancer prevention and treatment with resveratrol: From rodent studies to clinical trials. Cancer Prev. Res 2009, 2, 409–418. [Google Scholar]
- Kerr, J.F.R.; Winterford, C.M.; Harmon, B.V. Apoptosis: Its significance in cancer and cancer therapy. Cancer 1994, 73, 2013–2026. [Google Scholar]
- Petrache, I.K.; Birukov, A.L.; Zaiman, M.T.; Crow, H.; Deng, R.; Wadgaonkar, L.H.; Romer, J.; Garcia, G.N. Caspase-Dependent cleavage of myosin light chain kinase (MLCK) is involved in TNF-alpha-mediated bovine pulmonary endothelial cell apoptosis. FASEB J 2003, 17, 407–416. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | n | Body weight (g) | Liver weight (g) | Liver weight (g)/Body weight (g) × 100% |
|---|---|---|---|---|
| A | 8 | 309.4 ± 23.2 | 10.12 ± 2.01 | 3.26 ± 0.23 |
| B | 8 | 307.9 ± 32.1 | 10.03 ± 2.89 | 3.25 ± 0.31 |
| C | 8 | 278.3 ± 31.2 | 13.08 ± 3.24 | 4.69 ± 0.47 ** |
| D | 8 | 280.1 ± 29.6 | 14.02 ± 3.53 | 4.71 ± 0.56 ** |
| E | 8 | 299.3 ± 27.1 | 11.82 ± 2.25 | 3.91 ± 0.36 * |
| Group | n | Number of rats with nodules | Nodule incidence % | Total number of nodules | Average number of nodules per nodule-bearing liver |
|---|---|---|---|---|---|
| C | 8 | 8 | 100% | 297 | 36.8 ± 4.3 |
| D | 8 | 8 | 100% | 291 | 36.2 ± 6.9 |
| E | 8 | 5 | 62.5% * | 53 | 10.1 ± 2.6 ** |
© 2013 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Zhang, X.-L.; Yu, H.; Xiong, Y.-Y.; Ma, S.-T.; Zhao, L.; She, S.-F. Resveratrol Down-Regulates Myosin Light Chain Kinase, Induces Apoptosis and Inhibits Diethylnitrosamine-Induced Liver Tumorigenesis in Rats. Int. J. Mol. Sci. 2013, 14, 1940-1951. https://doi.org/10.3390/ijms14011940
Zhang X-L, Yu H, Xiong Y-Y, Ma S-T, Zhao L, She S-F. Resveratrol Down-Regulates Myosin Light Chain Kinase, Induces Apoptosis and Inhibits Diethylnitrosamine-Induced Liver Tumorigenesis in Rats. International Journal of Molecular Sciences. 2013; 14(1):1940-1951. https://doi.org/10.3390/ijms14011940
Chicago/Turabian StyleZhang, Xiao-Lin, Hao Yu, You-Yi Xiong, Shi-Tang Ma, Lei Zhao, and Shi-Feng She. 2013. "Resveratrol Down-Regulates Myosin Light Chain Kinase, Induces Apoptosis and Inhibits Diethylnitrosamine-Induced Liver Tumorigenesis in Rats" International Journal of Molecular Sciences 14, no. 1: 1940-1951. https://doi.org/10.3390/ijms14011940
APA StyleZhang, X. -L., Yu, H., Xiong, Y. -Y., Ma, S. -T., Zhao, L., & She, S. -F. (2013). Resveratrol Down-Regulates Myosin Light Chain Kinase, Induces Apoptosis and Inhibits Diethylnitrosamine-Induced Liver Tumorigenesis in Rats. International Journal of Molecular Sciences, 14(1), 1940-1951. https://doi.org/10.3390/ijms14011940