Apoptosis is Induced in Cancer Cells via the Mitochondrial Pathway by the Novel Xylocydine-Derived Compound JRS-15

Abstract
:1. Introduction
2. Results
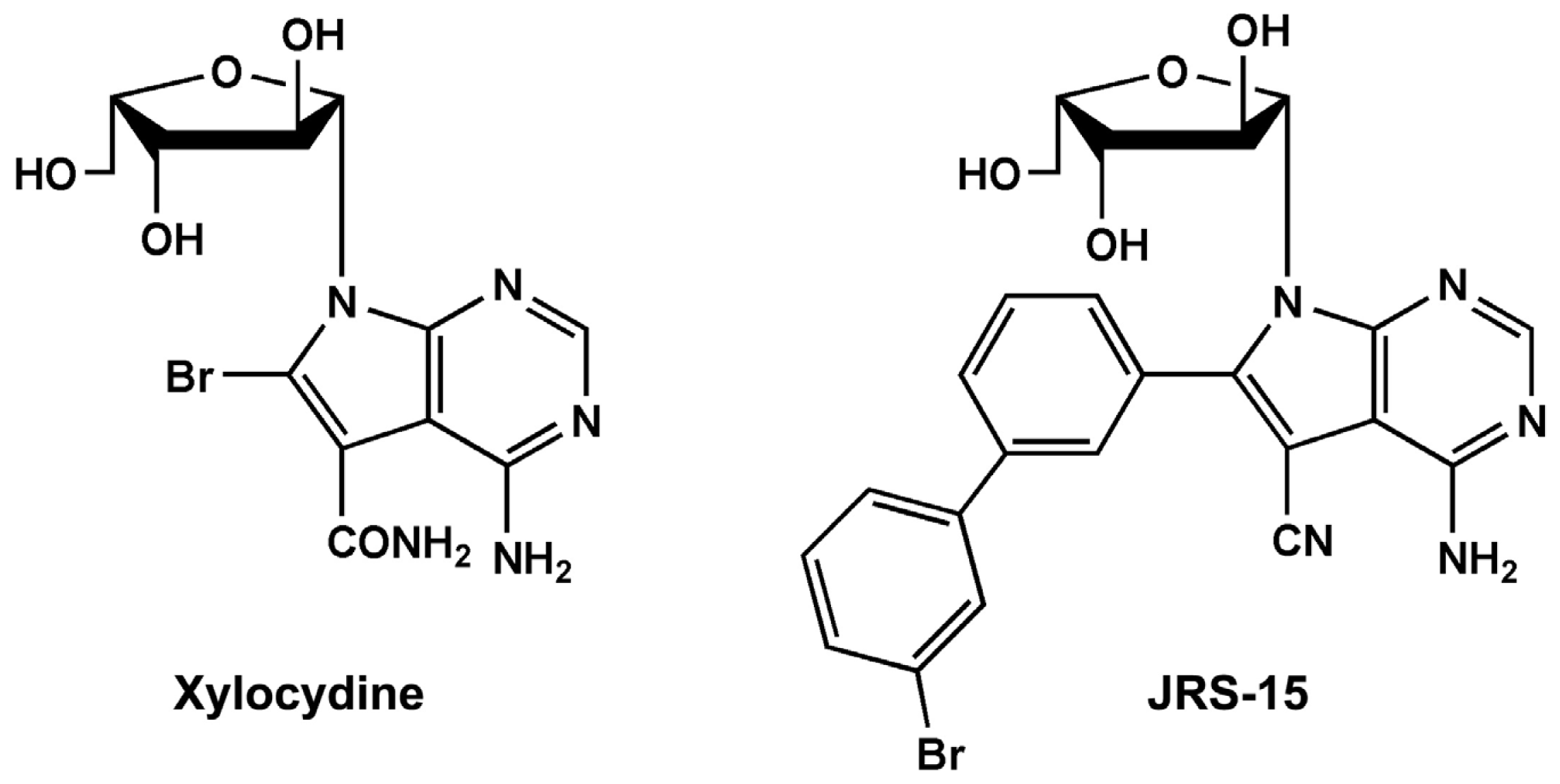
2.1. Synthesizing a Novel Compound JRS-15
2.2. JRS-15 Inhibits the Growth of Various Cancer Cells
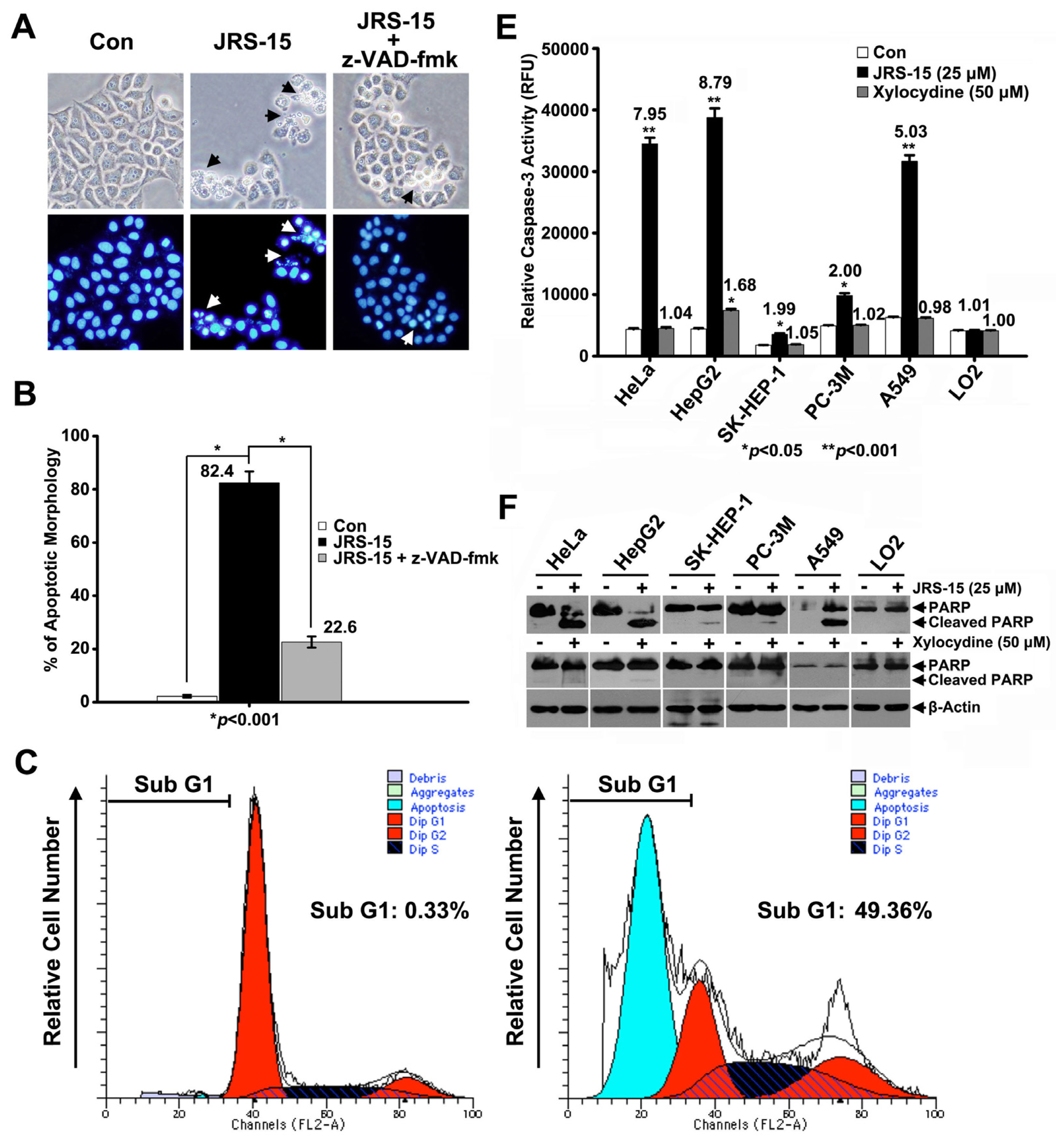
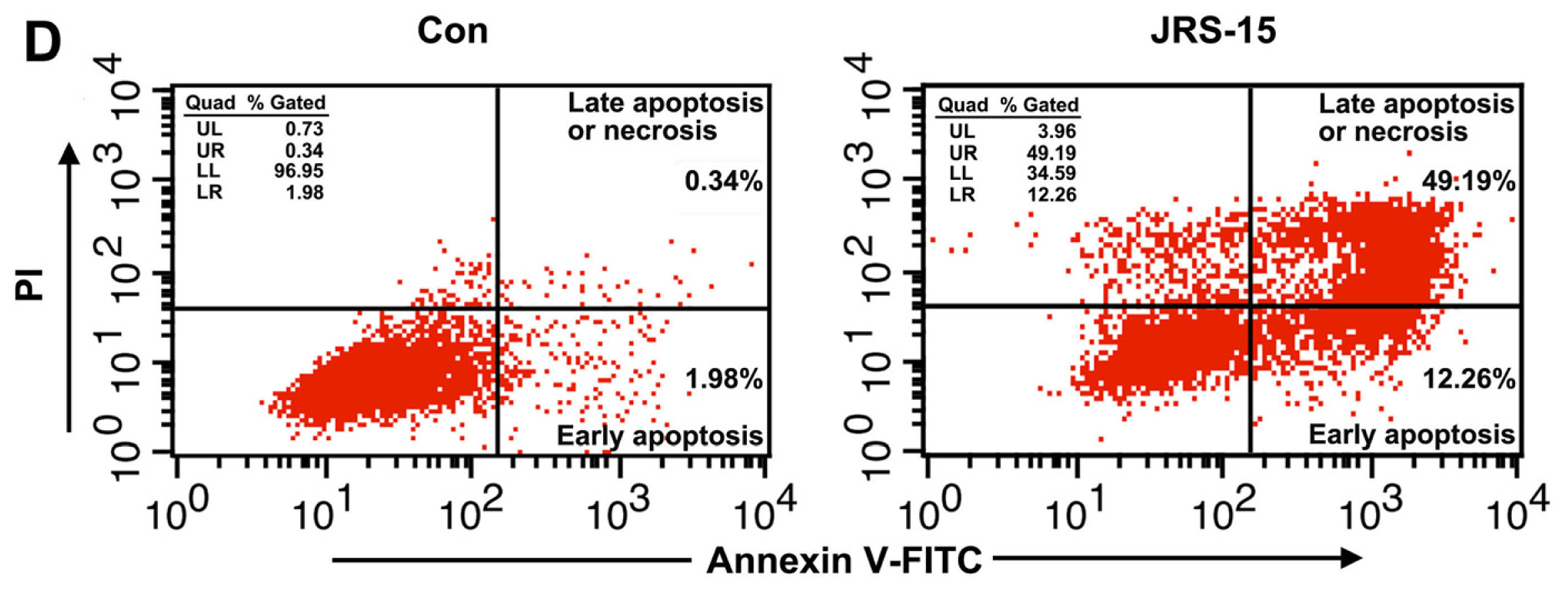
2.3. JRS-15 Induces Apoptotic Cell Death in Various Cancer Cells
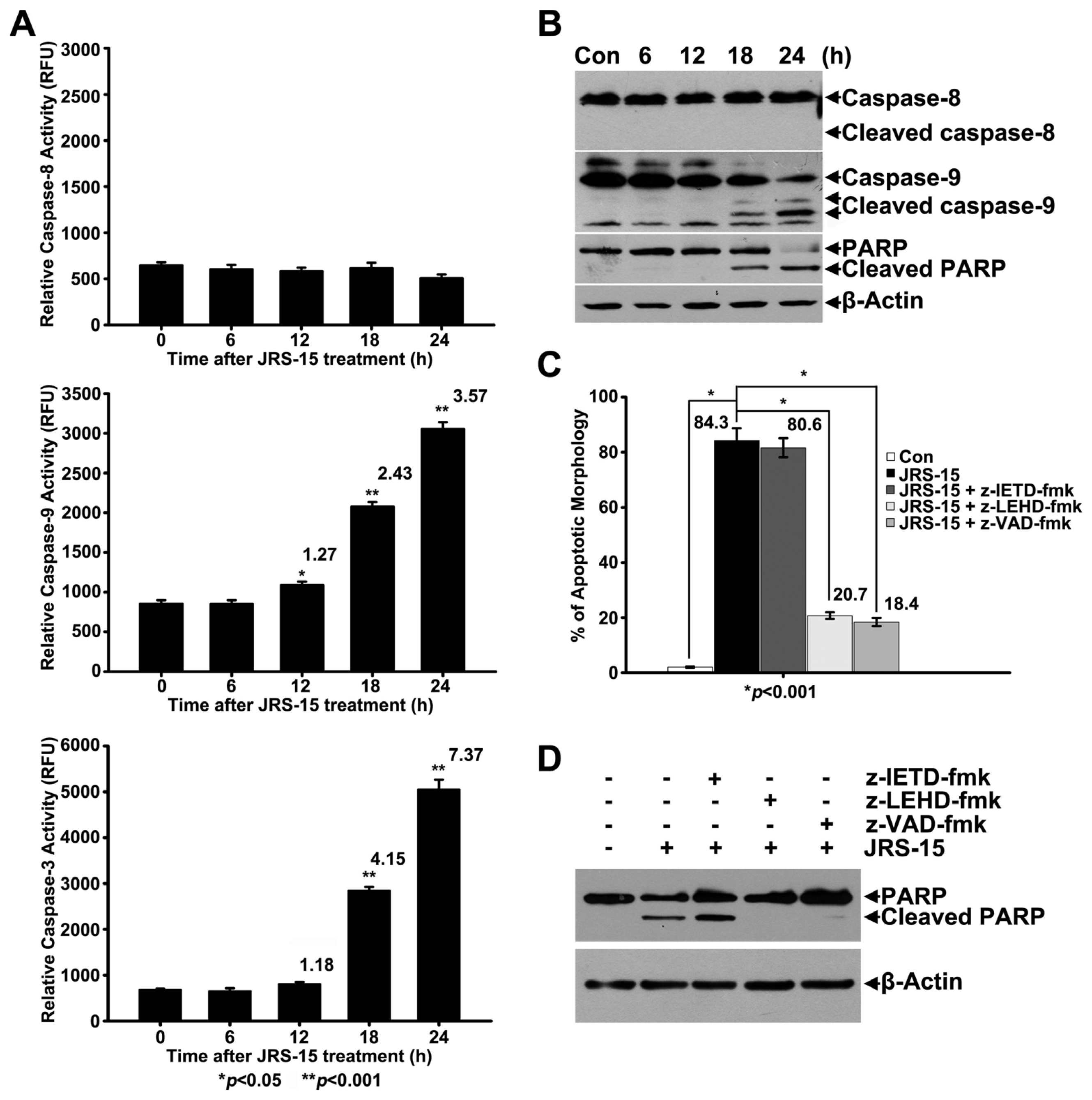
2.4. JRS-15-Induced Apoptosis in HeLa Cells is Caspase-9-Dependent
2.5. JRS-15 Triggers the Mitochondrial Apoptotic Pathway in HeLa Cells
2.6. JRS-15 Reduces Bcl-xL and XIAP Protein Levels in HeLa Cells
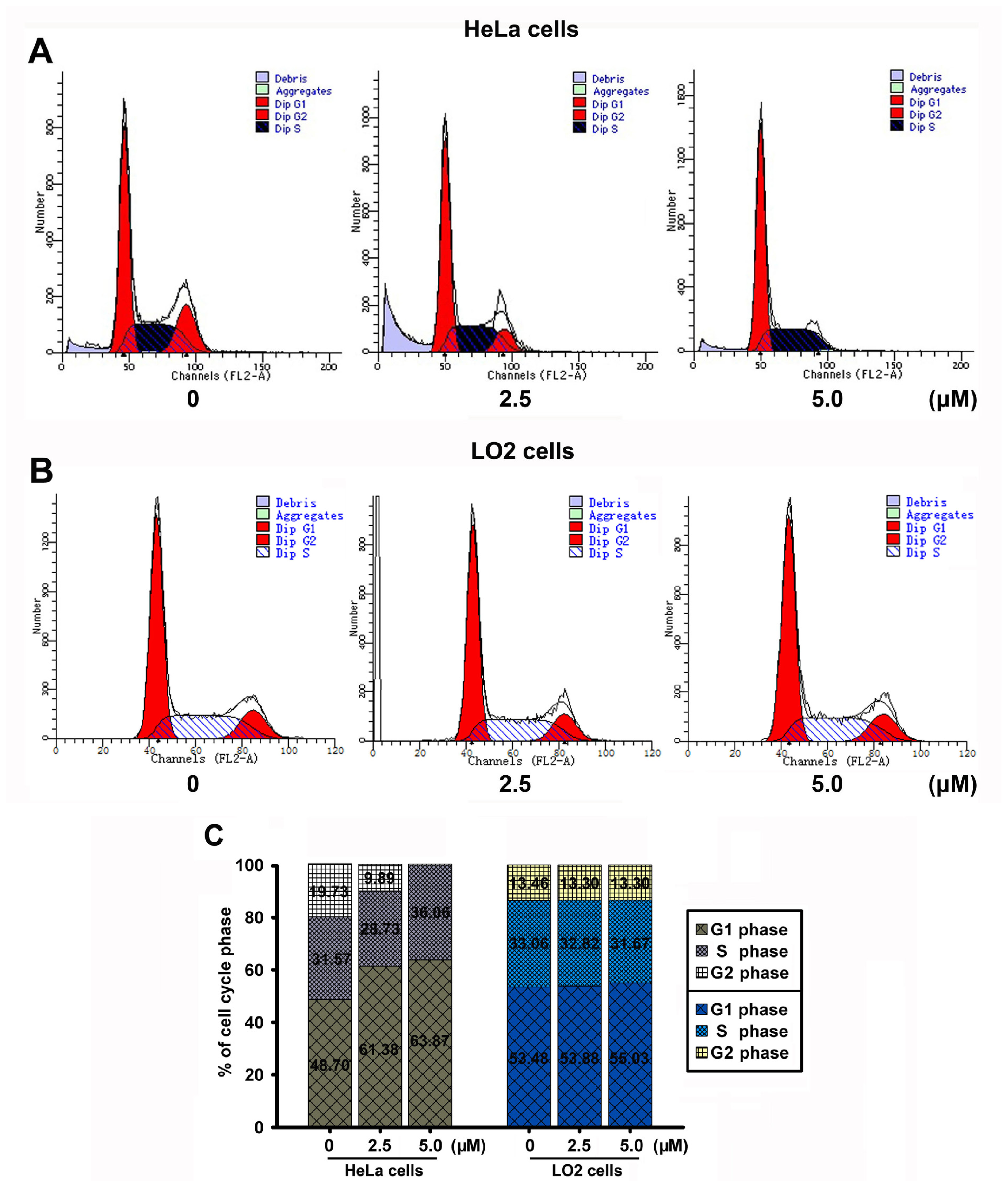
2.7. JRS-15 Induces Cell Cycle Arrest in HeLa Cells
2.8. Low Caspase-9 Expression in SK-HEP-1 Cells May Contribute to Their Resistance to JRS-15-Induced Apoptosis
3. Discussion
4. Experimental Section
4.1. Chemistry: General
4.2. 4-Amino-6-(3-(3-Bromophenyl)Phenyl)-7-(β-l-Xylofuranosyl)Pyrolo[2,3-d]Pyrimidine-5-Cyano (JRS-15)
4.3. Reagents and Antibodies
4.4. Cell Culture and MTT Assay
4.5. DAPI Staining Assay
4.6. Cell Cycle Distribution Assay
4.7. Annexin V-FITC/PI Double Staining Assay
4.8. Cell-Free Caspase Activity Assay
4.9. Immunoblot Analysis
4.10. Preparation of Mitochondrial and Cytosolic Protein Extracts
4.11. Depolarization Assay of Mitochondrial Membrane Potential
4.12. Transient Transfection Assay
4.13. Analysis the Value of AlogP
5. Conclusions
Supplementary Information
ijms-14-00850-s001.pdfAcknowledgments
- Conflict of InterestThe authors declare no conflict of interest.
References
- Jemal, A.; Bray, F.; Center, M.M.; Ferlay, J.; Ward, E.; Forman, D. Global cancer statistics. CA Cancer J. Clin 2011, 61, 69–90. [Google Scholar]
- Brown, J.M.; Attardi, L.D. The role of apoptosis in cancer development and treatment response. Nat. Rev. Cancer 2005, 5, 231–237. [Google Scholar]
- Fulda, S.; Debatin, K.M. Extrinsic versus intrinsic apoptosis pathways in anticancer chemotherapy. Oncogene 2006, 25, 4798–4811. [Google Scholar]
- Green, D.R. Apoptotic pathways: Paper wraps stone blunts scissors. Cell 2000, 102, 1–4. [Google Scholar]
- Ashkenazi, A.; Dixit, V.M. Death receptors: Signaling and modulation. Science 1998, 281, 1305–1308. [Google Scholar]
- Micheau, O.; Tschopp, J. Induction of TNF receptor I-mediated apoptosis via two sequential signaling complexes. Cell 2003, 114, 181–190. [Google Scholar]
- Mahmood, Z.; Shukla, Y. Death receptors: Targets for cancer therapy. Exp. Cell Res 2010, 316, 887–899. [Google Scholar]
- Patron, J.P.; Fendler, A.; Bild, M.; Jung, U.; Muller, H.; Arntzen, M.O.; Piso, C.; Stephan, C.; Thiede, B.; Mollenkopf, H.J.; et al. MiR-133b targets antiapoptotic genes and enhances death receptor-induced apoptosis. PLoS One 2012, 7, e35345. [Google Scholar]
- Yang, J.; Liu, X.; Bhalla, K.; Kim, C.N.; Ibrado, A.M.; Cai, J.; Peng, T.I.; Jones, D.P.; Wang, X. Prevention of apoptosis by Bcl-2: Release of cytochrome c from mitochondria blocked. Science 1997, 275, 1129–1132. [Google Scholar]
- Adrain, C.; Creagh, E.M.; Martin, S.J. Apoptosis-associated release of Smac/DIABLO from mitochondria requires active caspases and is blocked by Bcl-2. EMBO J 2001, 20, 6627–6636. [Google Scholar]
- Cande, C.; Cohen, I.; Daugas, E.; Ravagnan, L.; Larochette, N.; Zamzami, N.; Kroemer, G. Apoptosis-inducing factor (AIF): A novel caspase-independent death effector released from mitochondria. Biochimie 2002, 84, 215–222. [Google Scholar]
- Vaux, D.L. Apoptogenic factors released from mitochondria. Biochim. Biophys. Acta 2011, 1813, 546–550. [Google Scholar]
- Du, C.; Fang, M.; Li, Y.; Li, L.; Wang, X. Smac, a mitochondrial protein that promotes cytochrome c-dependent caspase activation by eliminating IAP inhibition. Cell 2000, 102, 33–42. [Google Scholar]
- Verhagen, A.M.; Ekert, P.G.; Pakusch, M.; Silke, J.; Connolly, L.M.; Reid, G.E.; Moritz, R.L.; Simpson, R.J.; Vaux, D.L. Identification of DIABLO, a mammalian protein that promotes apoptosis by binding to and antagonizing IAP proteins. Cell 2000, 102, 43–53. [Google Scholar]
- Vucic, D.; Deshayes, K.; Ackerly, H.; Pisabarro, M.T.; Kadkhodayan, S.; Fairbrother, W.J.; Dixit, V.M. SMAC negatively regulates the anti-apoptotic activity of melanoma inhibitor of apoptosis (ML-IAP). J. Biol. Chem 2002, 277, 12275–12279. [Google Scholar]
- Martinou, J.C.; Youle, R.J. Mitochondria in apoptosis: Bcl-2 family members and mitochondrial dynamics. Dev. Cell 2011, 21, 92–101. [Google Scholar]
- Chipuk, J.E.; Green, D.R. How do BCL-2 proteins induce mitochondrial outer membrane permeabilization? Trends Cell Biol 2008, 18, 157–164. [Google Scholar]
- Wei, M.C.; Zong, W.X.; Cheng, E.H.; Lindsten, T.; Panoutsakopoulou, V.; Ross, A.J.; Roth, K.A.; MacGregor, G.R.; Thompson, C.B.; Korsmeyer, S.J. Proapoptotic BAX and BAK: A requisite gateway to mitochondrial dysfunction and death. Science 2001, 292, 727–730. [Google Scholar]
- Eskes, R.; Desagher, S.; Antonsson, B.; Martinou, J.C. Bid induces the oligomerization and insertion of Bax into the outer mitochondrial membrane. Mol. Cell Biol 2000, 20, 929–935. [Google Scholar]
- Kuwana, T.; Bouchier-Hayes, L.; Chipuk, J.E.; Bonzon, C.; Sullivan, B.A.; Green, D.R.; Newmeyer, D.D. BH3 domains of BH3-only proteins differentially regulate Bax-mediated mitochondrial membrane permeabilization both directly and indirectly. Mol. Cell 2005, 17, 525–535. [Google Scholar]
- Ren, D.; Tu, H.C.; Kim, H.; Wang, G.X.; Bean, G.R.; Takeuchi, O.; Jeffers, J.R.; Zambetti, G.P.; Hsieh, J.J.; Cheng, E.H. BID, BIM, and PUMA are essential for activation of the BAX- and BAK-dependent cell death program. Science 2010, 330, 1390–1393. [Google Scholar]
- Cory, S.; Adams, J.M. The Bcl2 family: Regulators of the cellular life-or-death switch. Nat. Rev. Cancer 2002, 2, 647–656. [Google Scholar]
- Edlich, F.; Banerjee, S.; Suzuki, M.; Cleland, M.M.; Arnoult, D.; Wang, C.; Neutzner, A.; Tjandra, N.; Youle, R.J. Bcl-x(L) retrotranslocates Bax from the mitochondria into the cytosol. Cell 2011, 145, 104–116. [Google Scholar]
- Fletcher, J.I.; Meusburger, S.; Hawkins, C.J.; Riglar, D.T.; Lee, E.F.; Fairlie, W.D.; Huang, D.C.; Adams, J.M. Apoptosis is triggered when prosurvival Bcl-2 proteins cannot restrain Bax. Proc. Natl. Acad. Sci. USA 2008, 105, 18081–18087. [Google Scholar]
- Ham, Y.M. Xylocydine, a novel inhibitor of cyclin-dependent kinases, prevents the tumor necrosis factor-related apoptosis-inducing ligand-induced apoptotic cell death of SK-HEP-1 cells. J. Pharmacol. Exp. Ther 2003, 308, 814–819. [Google Scholar]
- Cho, S.-J.; Lee, S.-S.; Kim, Y.-J.; Park, B.-D.; Choi, J.-S.; Liu, L.; Ham, Y.-M.; Moon Kim, B.; Lee, S.-K. Xylocydine, a novel Cdk inhibitor, is an effective inducer of apoptosis in hepatocellular carcinoma cells in vitro and in vivo. Cancer Lett 2010, 287, 196–206. [Google Scholar]
- Lipinski, C.A.; Lombardo, F.; Dominy, B.W.; Feeney, P.J. Experimental and computational approaches to estimate solubility and permeability in drug discovery and development settings. Adv. Drug Deliv. Rev 2001, 46, 3–26. [Google Scholar]
- Ghose, A.K.; Viswanadhan, V.N.; Wendoloski, J.J. A knowledge-based approach in designing combinatorial or medicinal chemistry libraries for drug discovery. 1. A qualitative and quantitative characterization of known drug databases. J. Comb. Chem 1999, 1, 55–68. [Google Scholar]
- Sinhababu, A.K.; Thakker, D.R. Prodrugs of anticancer agents. Adv. Drug Deliv. Rev 1996, 19, 241–273. [Google Scholar]
- Xiao, C.; Sun, C.; Han, W.; Pan, F.; Dan, Z.; Li, Y.; Song, Z.-G.; Jin, Y.-H. Synthesis of 6-(het) ary Xylocydine analogues and evaluating their inhibitory activities of CDK1 and CDK2 in vitro. Bioorg. Med. Chem 2011, 19, 7100–7110. [Google Scholar]
- Plati, J.; Bucur, O.; Khosravi-Far, R. Dysregulation of apoptotic signaling in cancer: Molecular mechanisms and therapeutic opportunities. J. Cell Biochem 2008, 104, 1124–1149. [Google Scholar]
- Wilson, T.R.; Johnston, P.G.; Longley, D.B. Anti-apoptotic mechanisms of drug resistance in cancer. Curr. Cancer Drug Targets 2009, 9, 307–319. [Google Scholar]
- Li, Y.; He, K.; Huang, Y.; Zheng, D.; Gao, C.; Cui, L.; Jin, Y.H. Betulin induces mitochondrial cytochrome c release associated apoptosis in human cancer cells. Mol. Carcinog 2010, 49, 630–640. [Google Scholar]
- Porcari, A.R.; Townsend, L.B. Total synthesis of the naturally occurring antibiotic toyocamycin using new and improved synthetic procedures. Nucleosides Nucleotides 1999, 18, 153–159. [Google Scholar]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell types | Cell line | JRS-15 | Xylocydine |
|---|---|---|---|
| IC50 (μM) | IC50 (μM) | ||
| Cervical carcinoma | HeLa | 12.50 | Exceeding 50 |
| Hepatic carcinoma | HepG2 | 12.42 | Exceeding 50 |
| SK-HEP-1 | 28.25 | Exceeding 50 | |
| Prostate carcinoma | PC-3M | 27.20 | Exceeding 50 |
| Lung adenocarcinoma | A549 | 14.25 | Exceeding 50 |
| Normal liver cell line | LO2 | Exceeding 50 | Exceeding 50 |
© 2013 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Sun, C.; Guo, X.-X.; Zhu, D.; Xiao, C.; Bai, X.; Li, Y.; Zhan, Z.; Li, X.-L.; Song, Z.-G.; Jin, Y.-H. Apoptosis is Induced in Cancer Cells via the Mitochondrial Pathway by the Novel Xylocydine-Derived Compound JRS-15. Int. J. Mol. Sci. 2013, 14, 850-870. https://doi.org/10.3390/ijms14010850
Sun C, Guo X-X, Zhu D, Xiao C, Bai X, Li Y, Zhan Z, Li X-L, Song Z-G, Jin Y-H. Apoptosis is Induced in Cancer Cells via the Mitochondrial Pathway by the Novel Xylocydine-Derived Compound JRS-15. International Journal of Molecular Sciences. 2013; 14(1):850-870. https://doi.org/10.3390/ijms14010850
Chicago/Turabian StyleSun, Chao, Xiao-Xi Guo, Dan Zhu, Chuan Xiao, Xiao Bai, Yang Li, Zhuo Zhan, Xiang-Long Li, Zhi-Guang Song, and Ying-Hua Jin. 2013. "Apoptosis is Induced in Cancer Cells via the Mitochondrial Pathway by the Novel Xylocydine-Derived Compound JRS-15" International Journal of Molecular Sciences 14, no. 1: 850-870. https://doi.org/10.3390/ijms14010850
APA StyleSun, C., Guo, X.-X., Zhu, D., Xiao, C., Bai, X., Li, Y., Zhan, Z., Li, X.-L., Song, Z.-G., & Jin, Y.-H. (2013). Apoptosis is Induced in Cancer Cells via the Mitochondrial Pathway by the Novel Xylocydine-Derived Compound JRS-15. International Journal of Molecular Sciences, 14(1), 850-870. https://doi.org/10.3390/ijms14010850