Overexpression of Ferredoxin, PETF, Enhances Tolerance to Heat Stress in Chlamydomonas reinhardtii

 ,
, 

Abstract
:1. Introduction
2. Results
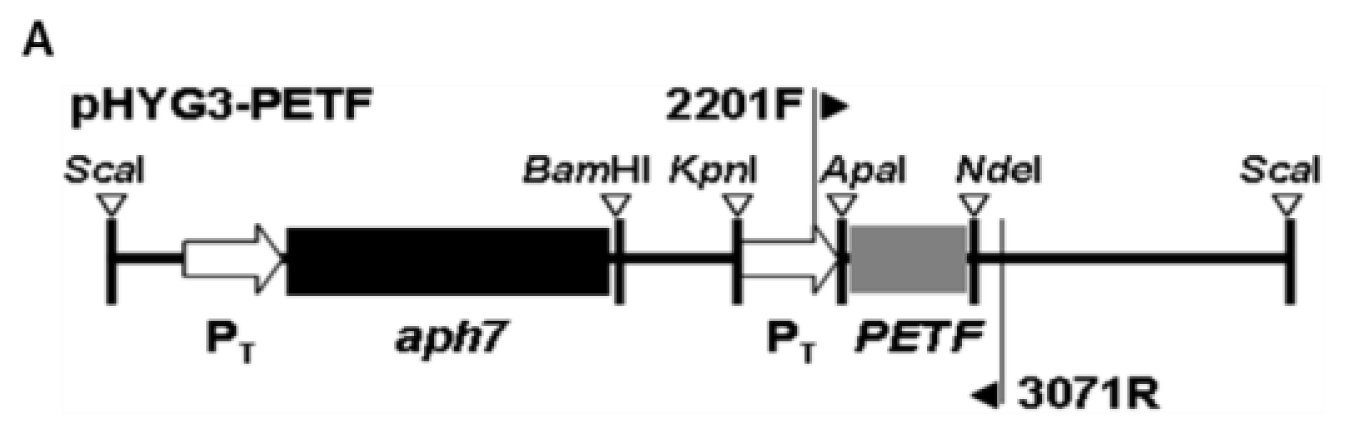
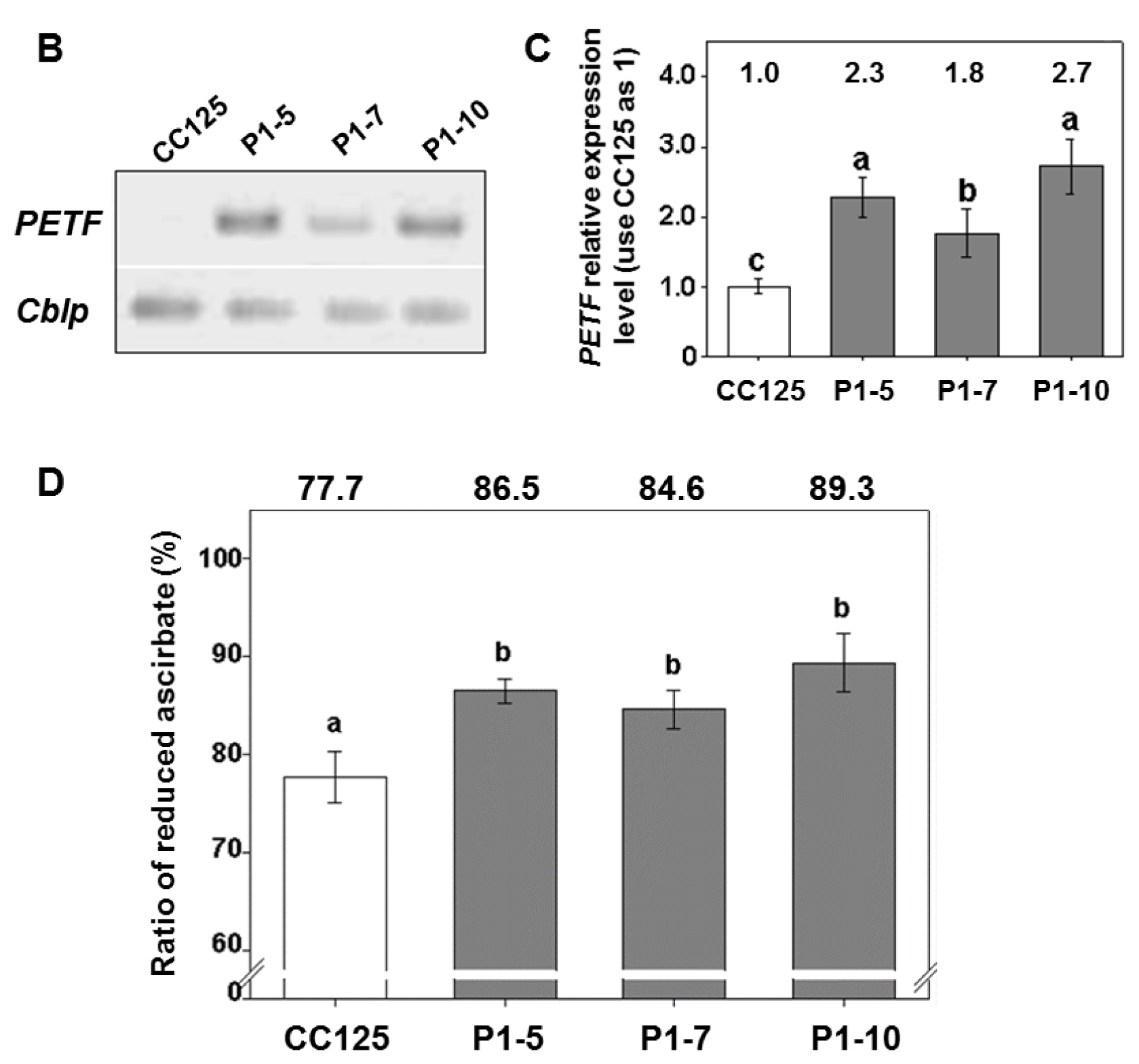
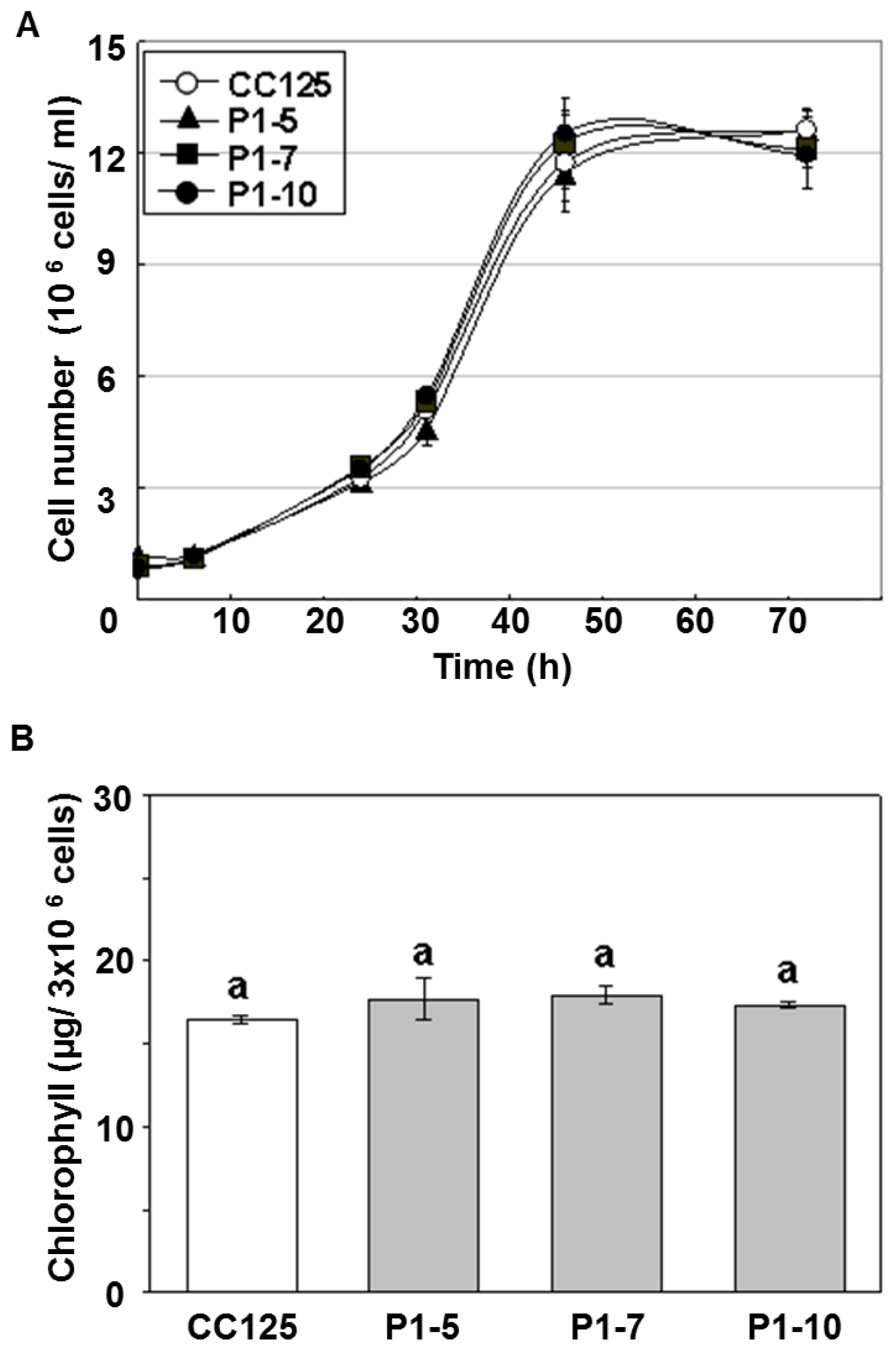
2.1. Generation and Characterizations of Transgenic Lines Overexpressing
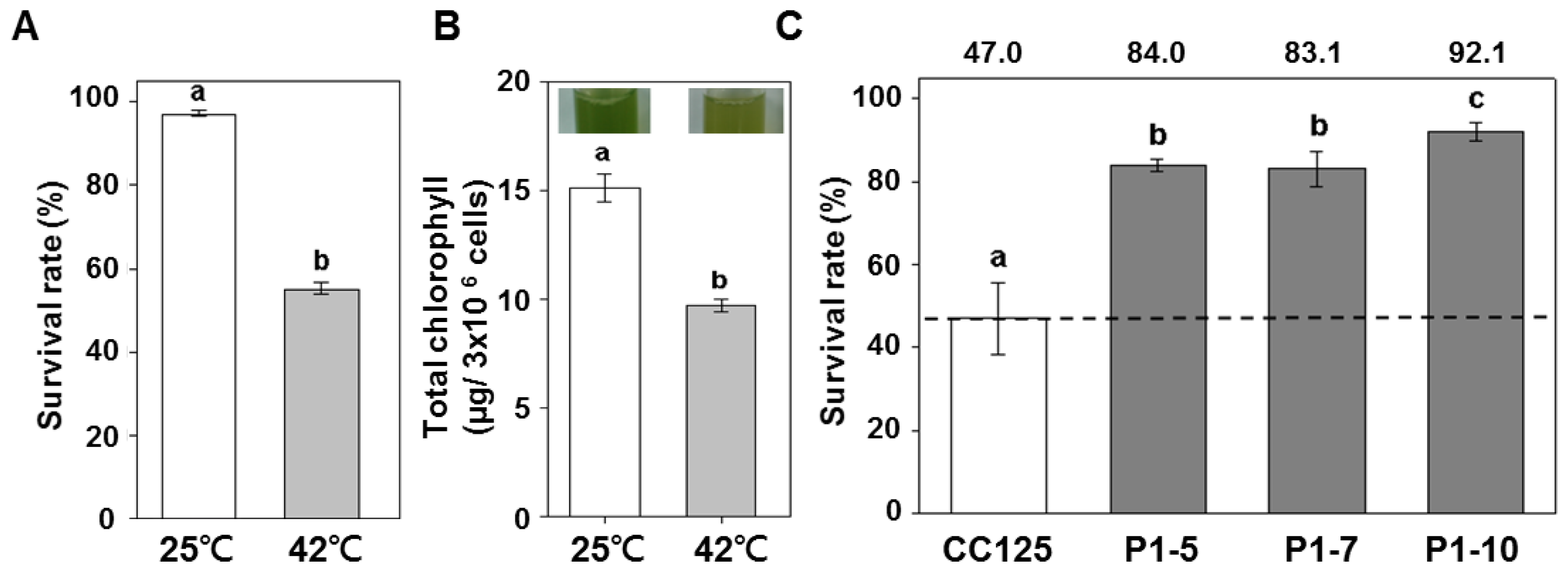
2.2. Overexpression of PETF in Chlamydomonas Enhances Stress Tolerance to Heat
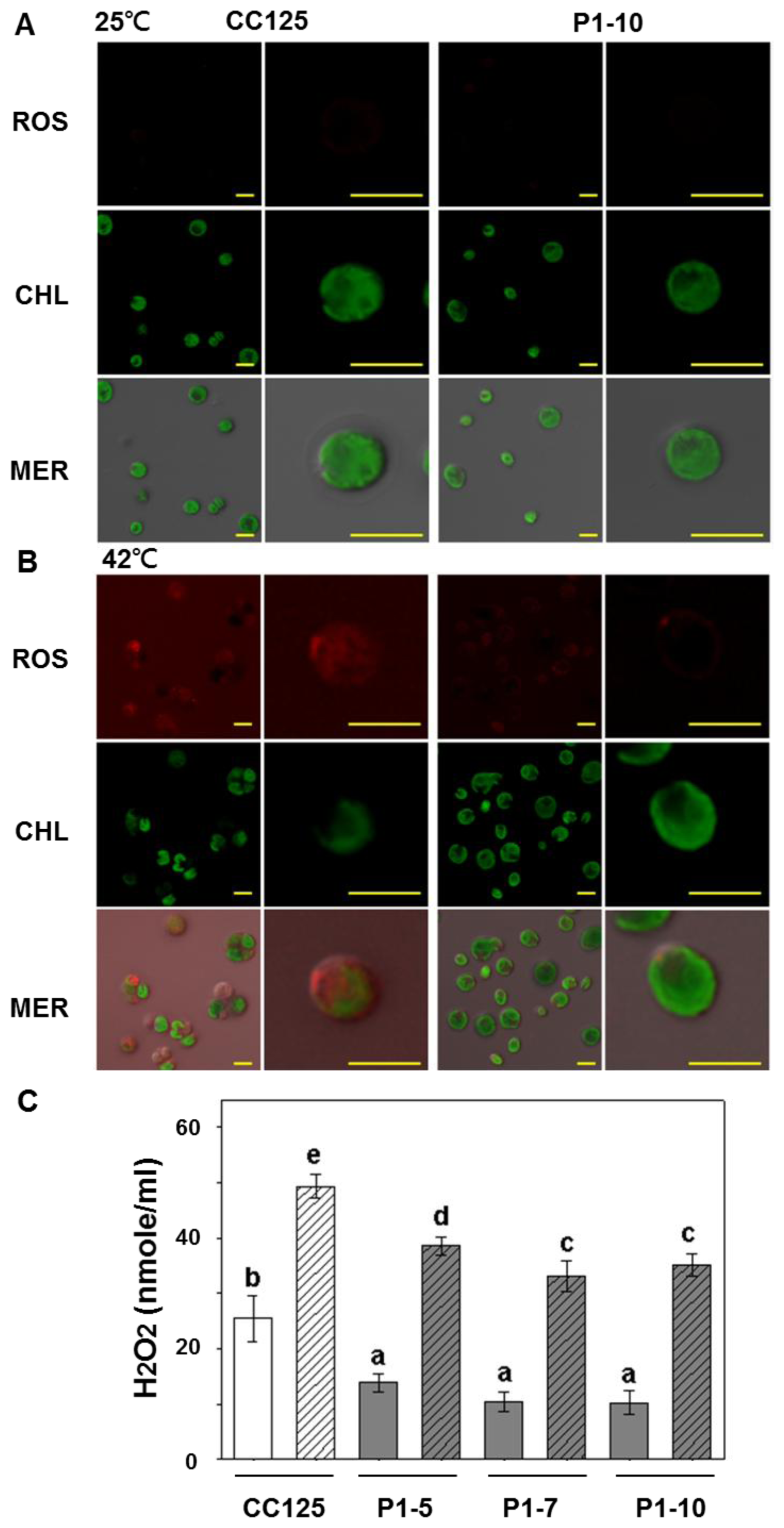
2.3. Accumulation of Reactive Oxygen Species (ROS) Is Reduced in Transgenic Lines Overexpressing PETF
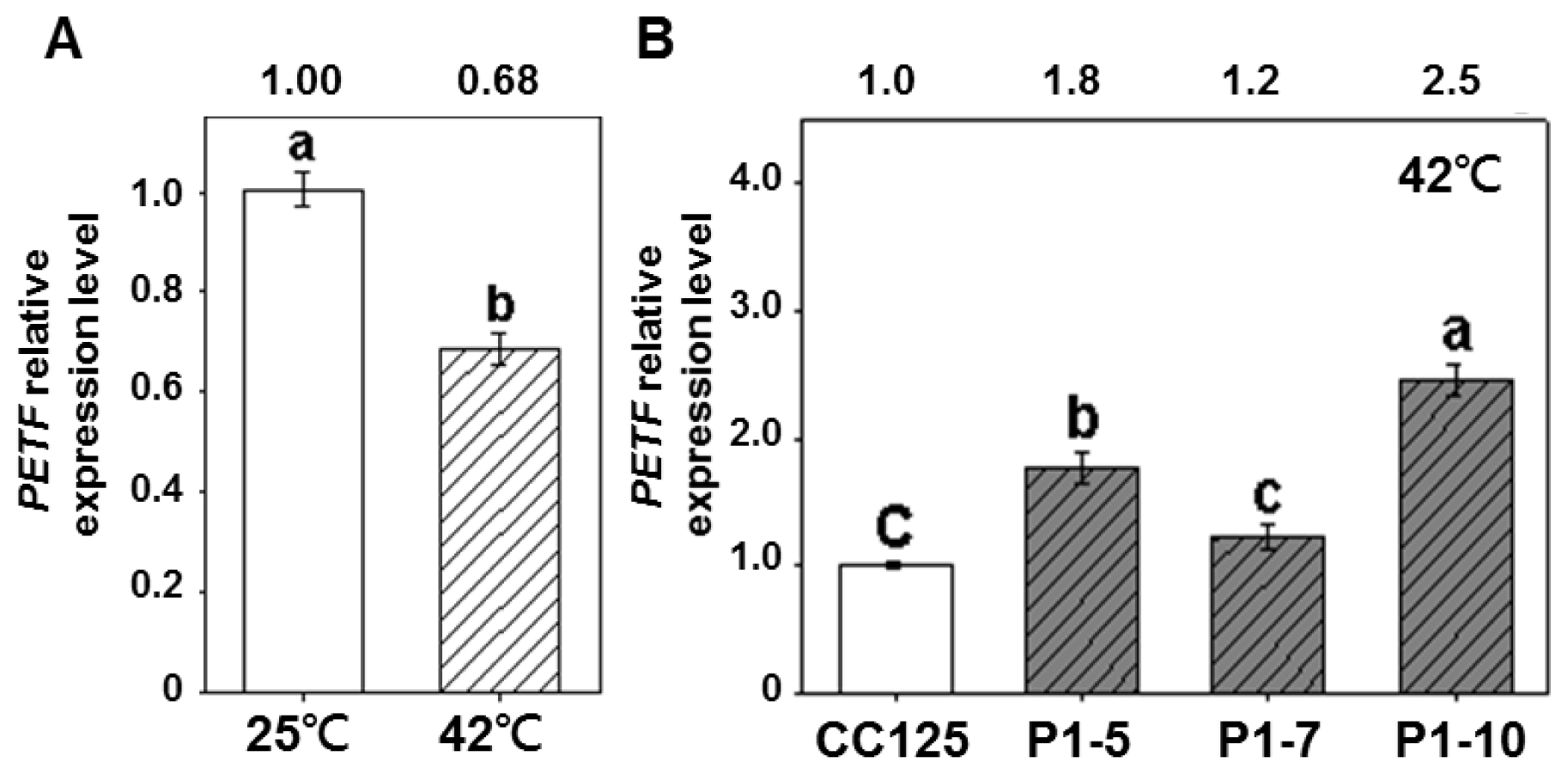
2.4. Expression of PETF Correlates Positively to Thermotolerance Ability in Transgenic Lines
3. Discussion
4. Experimental Section
4.1. Cultivation of Chlamydomonas reinhardtii
4.2. Construction of Plasmids
4.3. Gene Transformation of Chlamydomonas reinhardtii
4.4. Heat Treatment and Cell Survival Determination
4.5. Quantitative RT-PCR
4.6. Determination of Ascorbate Content
4.7. Reactive Oxygen Species (ROS) Detection and H2O2 Measurement
4.8. Determination of Chlorophyll Content
5. Conclusions







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Chlorophyll contents (μg/3 × 106 cells) | ||||||
|---|---|---|---|---|---|---|
| 25 °C | 42 °C | |||||
| Chl a | Chl b | Total | Chl a | Chl b | Total | |
| CC125 | 10.6 ± 0.6 a | 4.5 ± 0.1 a | 15.1 ± 0.6 a | 7.1 ± 0.8 a | 2.7 ± 0.2 a | 9.8 ± 1.0 a |
| P1-5 | 11.3 ± 0.9 a | 4.1 ± 0.3 a | 15.4 ± 1.1 a | 10.5 ± 0.8 c | 4.4 ± 0.3 c | 14.9 ± 1.1 c |
| P1-7 | 11.3 ± 0.7 a | 4.6 ± 0.3 a | 15.9 ± 1.0 a | 9.4 ± 0.1 b | 3.9 ± 0.1 b | 13.3 ± 0.2 b |
| P1-10 | 11.3 ± 0.6 a | 4.5 ± 0.1 a | 15.8 ± 0.8 a | 11.1 ± 0.6 c | 4.9 ± 0.1 d | 15.9 ± 0.6 c |
Acknowledgments
Conflicts of Interest
References
- Larkindale, J.; Knight, M.R. Protection against heat stress-induced oxidative damage in Arabidopsis involves calcium, abscisic acid, ethylene, and salicylic acid. Plant Physiol 2002, 128, 682–695. [Google Scholar]
- Allakhverdiev, S.I.; Kreslavski, V.D.; Klimov, V.V.; Los, D.A.; Carpentier, R.; Mohanty, P. Heat stress: An overview of molecular responses in photosynthesis. Photosynth. Res 2008, 98, 541–550. [Google Scholar]
- Foyer, C.H.; Lopez-Delgado, H.; Dat, J.F.; Scott, I.M. Hydrogen peroxide- and glutathione-associated mechanisms of acclimatory stress tolerance and signaling. Physiol. Plant 1997, 100, 241–254. [Google Scholar]
- Jakob, B.; Heber, U. Photoproduction and detoxification of hydroxyl radicals in chloroplasts and leaves and relation to photoinactivation of photosystems I and II. Plant Cell Physiol 1996, 37, 629–635. [Google Scholar]
- De Gara, L.; Locato, V.; Dipierro, S.; de Pinto, M.C. Redox homeostasis in plants. The challenge of living with endogenous oxygen production. Respir. Physiol. Neurobiol 2010, 173, S13–S19. [Google Scholar]
- Lintala, M.; Lehtimaki, N.; Benz, J.P.; Jungfer, A.; Soll, J.; Aro, E.M.; Bolter, B.; Mulo, P. Depletion of leaf-type ferredoxin-NADP(+) oxidoreductase results in the permanent induction of photoprotective mechanisms in Arabidopsis chloroplasts. Plant J 2012, 70, 809–817. [Google Scholar]
- Locato, V.; Gadaleta, C.; de Gara, L.; de Pinto, M.C. Production of reactive species and modulation of antioxidant network in response to heat shock: A critical balance for cell fate. Plant Cell Environ 2008, 31, 1606–1619. [Google Scholar]
- Davletova, S.; Rizhsky, L.; Liang, H.; Shengqiang, Z.; Oliver, D.J.; Coutu, J.; Shulaev, V.; Schlauch, K.; Mittler, R. Cytosolic ascorbate peroxidase 1 is a central component of the reactive oxygen gene network of Arabidopsis. Plant Cell 2005, 17, 268–281. [Google Scholar]
- Hasanuzzaman, M.; Nahar, K.; Alam, M.M.; Roychowdhury, R.; Fujita, M. Physiological, biochemical, and molecular mechanisms of heat stress tolerance in plants. Int. J. Mol. Sci 2013, 14, 9643–9684. [Google Scholar]
- Lv, W.T.; Lin, B.; Zhang, M.; Hua, X.J. Proline accumulation is inhibitory to Arabidopsis seedlings during heat stress. Plant Physiol 2011, 156, 1921–1933. [Google Scholar]
- Sathiyaraj, G.; Lee, O.R.; Parvin, S.; Khorolragchaa, A.; Kim, Y.J.; Yang, D.C. Transcript profiling of antioxidant genes during biotic and abiotic stresses in Panax ginseng C. A. Meyer. Mol. Biol. Rep 2011, 38, 2761–2769. [Google Scholar]
- Vacca, R.A.; Valenti, D.; Bobba, A.; Merafina, R.S.; Passarella, S.; Marra, E. Cytochrome c is released in a reactive oxygen species-dependent manner and is degraded via caspase-like proteases in tobacco Bright-Yellow 2 cells en route to heat shock-induced cell death. Plant Physiol 2006, 141, 208–219. [Google Scholar]
- Arnon, D.I. The discovery of ferredoxin: The photosynthetic path. Trends Biochem. Sci 1988, 13, 30–33. [Google Scholar]
- Schurmann, P.; Buchanan, B.B. The ferredoxin/thioredoxin system of oxygenic photosynthesis. Antioxid. Redox Signal 2008, 10, 1235–1274. [Google Scholar]
- Geigenberger, P.; Kolbe, A.; Tiessen, A. Redox regulation of carbon storage and partitioning in response to light and sugars. J. Exp. Bot 2005, 56, 1469–1479. [Google Scholar]
- Mulo, P. Chloroplast-targeted ferredoxin-NADP(+) oxidoreductase (FNR): Structure, function and location. Biochim. Biophys. Acta 2011, 1807, 927–934. [Google Scholar]
- Winkler, M.; Hemschemeier, A.; Jacobs, J.; Stripp, S.; Happe, T. Multiple ferredoxin isoforms in Chlamydomonas reinhardtii—Their role under stress conditions and biotechnological implications. Eur. J. Cell Biol 2010, 89, 998–1004. [Google Scholar]
- Asada, K. The Water-Water cycle in chloroplasts: scavenging of active oxygens and dissipation of excess photons. Annu. Rev. Plant Physiol. Plant Mol. Biol 1999, 50, 601–639. [Google Scholar]
- Rizhsky, L.; Liang, H.; Mittler, R. The water-water cycle is essential for chloroplast protection in the absence of stress. J. Biol. Chem 2003, 278, 38921–38925. [Google Scholar]
- Noctor, G.; Foyer, C.H. Ascorbate and glutathione: Keeping active oxygen under control. Annu. Rev. Plant Physiol. Plant Mol. Biol 1998, 49, 249–279. [Google Scholar]
- Zimmermann, P.; Hirsch-Hoffmann, M.; Hennig, L.; Gruissem, W. Genevestigator. Arabidopsis microarray database and analysis toolbox. Plant Physiol 2004, 136, 2621–2632. [Google Scholar]
- Tognetti, V.B.; Palatnik, J.F.; Fillat, M.F.; Melzer, M.; Hajirezaei, M.R.; Valle, E.M.; Carrillo, N. Functional replacement of ferredoxin by a cyanobacterial flavodoxin in tobacco confers broad-range stress tolerance. Plant Cell 2006, 18, 2035–2050. [Google Scholar]
- Holtgrefe, S.; Bader, K.P.; Horton, P.; Scheible, R.; von Schaewen, A.; Backhausen, J.E. Decreased content of leaf ferredoxin changes electron distribution and limits photosynthesis in transgenic potato plants. Plant Physiol 2003, 133, 1768–1778. [Google Scholar]
- Hanke, G.T.; Hase, T. Variable photosynthetic roles of two leaf-type ferredoxins in Arabidopsis, as revealed by RNA interference. Photochem. Photobiol 2008, 84, 1302–1309. [Google Scholar]
- Blanco, N.E.; Ceccoli, R.D.; Segretin, M.E.; Poli, H.O.; Voss, I.; Melzer, M.; Bravo-Almonacid, F.F.; Scheibe, R.; Hajirezaei, M.R.; Carrillo, N. Cyanobacterial flavodoxin complements ferredoxin deficiency in knocked-down transgenic tobacco plants. Plant J 2011, 65, 922–935. [Google Scholar]
- Zurbriggen, M.D.; Tognetti, V.B.; Carrillo, N. Stress-inducible flavodoxin from photosynthetic microorganisms. The mystery of flavodoxin loss from the plant genome. IUBMB Life 2007, 59, 355–360. [Google Scholar]
- Ceccoli, R.D.; Blanco, N.E.; Medina, M.; Carrillo, N. Stress response of transgenic tobacco plants expressing a cyanobacterial ferredoxin in chloroplasts. Plant Mol. Biol 2011, 76, 535–544. [Google Scholar]
- Hema, R.; Senthil-Kumar, M.; Shivakumar, S.; Reddy, P.C.; Udayakumar, M. Chlamydomonas reinhardtii, a model system for functional validation of abiotic stress responsive genes. Planta 2007, 226, 655–670. [Google Scholar]
- Noth, J.; Krawietz, D.; Hemschemeier, A.; Happe, T. Pyruvate:ferredoxin oxidoreductase is coupled to light-independent hydrogen production in Chlamydomonas reinhardtii. J. Biol. Chem 2013, 288, 4368–4377. [Google Scholar]
- Philipps, G.; Happe, T.; Hemschemeier, A. Nitrogen deprivation results in photosynthetic hydrogen production in Chlamydomonas reinhardtii. Planta 2012, 235, 729–745. [Google Scholar]
- Grossman, A.R.; Catalanotti, C.; Yang, W.; Dubini, A.; Magneschi, L.; Subramanian, V.; Posewitz, M.C.; Seibert, M. Multiple facets of anoxic metabolism and hydrogen production in the unicellular green alga Chlamydomonas reinhardtii. New Phytol 2011, 190, 279–288. [Google Scholar]
- Merchant, S.S.; Allen, M.D.; Kropat, J.; Moseley, J.L.; Long, J.C.; Tottey, S.; Terauchi, A.M. Between a rock and a hard place: Trace element nutrition in Chlamydomonas. Biochim. Biophys. Acta 2006, 1763, 578–594. [Google Scholar]
- Terashima, M.; Specht, M.; Hippler, M. The chloroplast proteome: A survey from the Chlamydomonas reinhardtii perspective with a focus on distinctive features. Curr. Genet 2011, 57, 151–168. [Google Scholar]
- Terauchi, A.M.; Lu, S.-F.; Zaffagnini, M.; Tappa, S.; Hirasawa, M.; Tripathy, J.N.; Knaff, D.B.; Farmer, P.J.; Lemaire, S.D.; Hase, T.; et al. Pattern of expression and substrate specificity of chloroplast ferredoxins from Chlamydomonas reinhardtii. J. Biol. Chem 2009, 284, 25867–25878. [Google Scholar]
- Schmitter, J.-M.; Jacquot, J.-P.; de Lamotte-Guéry, F.; Beauvallet, C.; Dutka, S.; Gadal, P.; Decottignies, P. Purification, properties and complete amino acid sequence of the ferredoxin from a green alga Clamydomonas reinhardtii. Eur. J. Biochem 1988, 172, 405–412. [Google Scholar]
- Lin, Y.H.; Huang, H.E.; Wu, F.S.; Ger, M.J.; Liao, P.L.; Chen, Y.R.; Tzeng, K.C.; Feng, T.Y. Plant ferredoxin-like protein (PFLP) outside chloroplast in Arabidopsis enhances disease resistance against bacterial pathogens. Plant Sci 2010, 179, 450–458. [Google Scholar]
- Khanna-Chopra, R. Leaf senescence and abiotic stresses share reactive oxygen species-mediated chloroplast degradation. Protoplasma 2012, 249, 469–481. [Google Scholar]
- Kumar, D.; Yusuf, M.A.; Singh, P.; Sardar, M.; Sarin, N.B. Modulation of antioxidant machinery in alpha-tocopherol-enriched transgenic Brassica juncea plants tolerant to abiotic stress conditions. Protoplasma 2013, 250, 1079–7089. [Google Scholar]
- Choe, Y.H.; Kim, Y.S.; Kim, I.S.; Bae, M.J.; Lee, E.J.; Kim, Y.H.; Park, H.M.; Yoon, H.S. Homologous expression of gamma-glutamylcysteine synthetase increases grain yield and tolerance of transgenic rice plants to environmental stresses. J. Plant Physiol 2013, 170, 610–618. [Google Scholar]
- Kavitha, K.; George, S.; Venkataraman, G.; Parida, A. A salt-inducible chloroplastic monodehydroascorbate reductase from halophyte Avicennia marina confers salt stress tolerance on transgenic plants. Biochimie 2010, 92, 1321–1329. [Google Scholar]
- Singh, A.; Grover, A. Genetic engineering for heat tolerance in plants. Physiol. Mol. Biol. Plants 2008, 14, 155–166. [Google Scholar]
- Hanke, G.; Mulo, P. Plant type ferredoxins and ferredoxin-dependent metabolism. Plant Cell Environ 2013, 36, 1071–1084. [Google Scholar]
- Scheumann, V.; Schoch, S.; Rudiger, W. Chlorophyll a formation in the chlorophyll b reductase reaction requires reduced ferredoxin. J. Biol. Chem 1998, 273, 35102–35108. [Google Scholar]
- Reinbothe, C.; Bartsch, S.; Eggink, L.L.; Hoober, J.K.; Brusslan, J.; Andrade-Paz, R.; Monnet, J.; Reinbothe, S. A role for chlorophyllide a oxygenase in the regulated import and stabilization of light-harvesting chlorophyll a/b proteins. Proc. Natl. Acad. Sci. USA 2006, 103, 4777–4782. [Google Scholar]
- Tanaka, A.; Ito, H.; Tanaka, R.; Tanaka, N.K.; Yoshida, K.; Okada, K. Chlorophyll a oxygenase (CAO) is involved in chlorophyll b formation from chlorophyll a. Proc. Natl. Acad. Sci. USA 1998, 95, 12719–12723. [Google Scholar]
- Rodoni, S.; Muhlecker, W.; Anderl, M.; Krautler, B.; Moser, D.; Thomas, H.; Matile, P.; Hortensteiner, S. Chlorophyll breakdown in senescent chloroplasts (cleavage of pheophorbide a in two enzymic steps). Plant Physiol 1997, 115, 669–676. [Google Scholar]
- Op den Camp, R.G.; Przybyla, D.; Ochsenbein, C.; Laloi, C.; Kim, C.; Danon, A.; Wagner, D.; Hideg, E.; Gobel, C.; Feussner, I.; et al. Rapid induction of distinct stress responses after the release of singlet oxygen in Arabidopsis. Plant Cell 2003, 15, 2320–2332. [Google Scholar]
- Larkindale, J.; Huang, B. Thermotolerance and antioxidant systems in Agrostis stolonifera: Involvement of salicylic acid, abscisic acid, calcium, hydrogen peroxide, and ethylene. J. Plant Physiol 2004, 161, 405–413. [Google Scholar]
- Benning, C. Mechanisms of lipid transport involved in organelle biogenesis in plant cells. Ann. Rev. Cell Dev. Biol 2009, 25, 71–91. [Google Scholar]
- Ohlrogge, J.; Browse, J. Lipid biosynthesis. Plant Cell 1995, 7, 957–970. [Google Scholar]
- Jouhet, J.; Marechal, E.; Block, M.A. Glycerolipid transfer for the building of membranes in plant cells. Prog. Lipid Res 2007, 46, 37–55. [Google Scholar] [Green Version]
- Meguro, M.; Ito, H.; Takabayashi, A.; Tanaka, R.; Tanaka, A. Identification of the 7-hydroxymethyl chlorophyll a reductase of the chlorophyll cycle in Arabidopsis. Plant Cell 2011, 23, 3442–3453. [Google Scholar]
- Suzuki, N.; Koussevitzky, S.; Mittler, R.; Miller, G. ROS and redox signalling in the response of plants to abiotic stress. Plant Cell Environ 2012, 35, 259–270. [Google Scholar]
- Fini, A.; Brunetti, C.; di Ferdinando, M.; Ferrini, F.; Tattini, M. Stress-induced flavonoid biosynthesis and the antioxidant machinery of plants. Plant Signal. Behav 2011, 6, 709–711. [Google Scholar]
- Galvez-Valdivieso, G.; Mullineaux, P.M. The role of reactive oxygen species in signalling from chloroplasts to the nucleus. Physiol. Plant 2010, 138, 430–439. [Google Scholar]
- Rodriguez, R.E.; Lodeyro, A.; Poli, H.O.; Zurbriggen, M.; Peisker, M.; Palatnik, J.F.; Tognetti, V.B.; Tschiersch, H.; Hajirezaei, M.R.; Valle, E.M.; et al. Transgenic tobacco plants overexpressing chloroplastic ferredoxin-NADP(H) reductase display normal rates of photosynthesis and increased tolerance to oxidative stress. Plant Physiol 2007, 143, 639–649. [Google Scholar]
- Mediavilla, M.G.; di Venanzio, G.A.; Guibert, E.E.; Tiribelli, C. Heterologous ferredoxin reductase and flavodoxin protect Cos-7 cells from oxidative stress. PLoS One 2010, 5, e13501. [Google Scholar]
- Mano, J.; Ohno, C.; Domae, Y.; Asada, K. Chloroplastic ascorbate peroxidase is the primary target of methylviologen-induced photooxidative stress in spinach leaves: Its relevance to monodehydroascorbate radical detected with in vivo ESR. Biochim. Biophys. Acta 2001, 1504, 275–287. [Google Scholar]
- Giacomelli, L.; Masi, A.; Ripoll, D.R.; Lee, M.J.; van Wijk, K.J. Arabidopsis thaliana deficient in two chloroplast ascorbate peroxidases shows accelerated light-induced necrosis when levels of cellular ascorbate are low. Plant Mol. Biol 2007, 65, 627–644. [Google Scholar]
- Smirnoff, N. Ascorbate biosynthesis and function in photoprotection. Philos. Trans. R. Soc. Lond. B 2000, 355, 1455–1464. [Google Scholar]
- Gorman, D.S.; Levine, R.P. Cytochrome f and plastocyanin: Their sequence in the photosynthetic electron transport chain of Chlamydomonas reinhardtii. Proc. Natl. Acad. Sci. USA 1965, 54, 1665–1669. [Google Scholar]
- Stern, D.; Witman, G.; Harris, E.H. The Chlamydomonas Sourcebook, 2nd ed.; Academic Press: Waltham, MA, USA, 2008. [Google Scholar]
- Berthold, P.; Schmitt, R.; Mages, W. An engineered Streptomyces hygroscopicus aph 7" gene mediates dominant resistance against hygromycin B in Chlamydomonas reinhardtii. Protist 2002, 153, 401–412. [Google Scholar]
- Shimogawara, K.; Fujiwara, S.; Grossman, A.; Usuda, H. High-efficiency transformation of Chlamydomonas reinhardtii by electroporation. Genetics 1998, 148, 1821–1828. [Google Scholar]
- Strober, W. Trypan blue exclusion test of cell viability. Curr. Protoc. Immunol 2001. [Google Scholar] [CrossRef]
- Gillespie, K.M.; Ainsworth, E.A. Measurement of reduced, oxidized and total ascorbate content in plants. Nat. Protoc 2007, 2, 871–874. [Google Scholar]
- Voss, I.; Koelmann, M.; Wojtera, J.; Holtgrefe, S.; Kitzmann, C.; Backhausen, J.E.; Scheibe, R. Knockout of major leaf ferredoxin reveals new redox-regulatory adaptations in Arabidopsis thaliana. Physiol. Plant 2008, 133, 584–598. [Google Scholar]
- Funes, S.; Franzén, L.-G.; González-Halphen, D. Chlamydomonas reinhardtii: The model of choice to study mitochondria from unicellular photosynthetic organisms. Methods Mol. Biol 2007, 372, 137–149. [Google Scholar]
© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Lin, Y.-H.; Pan, K.-Y.; Hung, C.-H.; Huang, H.-E.; Chen, C.-L.; Feng, T.-Y.; Huang, L.-F. Overexpression of Ferredoxin, PETF, Enhances Tolerance to Heat Stress in Chlamydomonas reinhardtii. Int. J. Mol. Sci. 2013, 14, 20913-20929. https://doi.org/10.3390/ijms141020913
Lin Y-H, Pan K-Y, Hung C-H, Huang H-E, Chen C-L, Feng T-Y, Huang L-F. Overexpression of Ferredoxin, PETF, Enhances Tolerance to Heat Stress in Chlamydomonas reinhardtii. International Journal of Molecular Sciences. 2013; 14(10):20913-20929. https://doi.org/10.3390/ijms141020913
Chicago/Turabian StyleLin, Yi-Hsien, Kui-You Pan, Ching-Hui Hung, Hsiang-En Huang, Ching-Lian Chen, Teng-Yung Feng, and Li-Fen Huang. 2013. "Overexpression of Ferredoxin, PETF, Enhances Tolerance to Heat Stress in Chlamydomonas reinhardtii" International Journal of Molecular Sciences 14, no. 10: 20913-20929. https://doi.org/10.3390/ijms141020913
APA StyleLin, Y.-H., Pan, K.-Y., Hung, C.-H., Huang, H.-E., Chen, C.-L., Feng, T.-Y., & Huang, L.-F. (2013). Overexpression of Ferredoxin, PETF, Enhances Tolerance to Heat Stress in Chlamydomonas reinhardtii. International Journal of Molecular Sciences, 14(10), 20913-20929. https://doi.org/10.3390/ijms141020913