The Ribonucleoprotein Csr Network

{kind=link}
{kind=link}
Abstract
:1. Introduction
1.1. A Plethora of Ribonucleoprotein Complexes in Bacteria
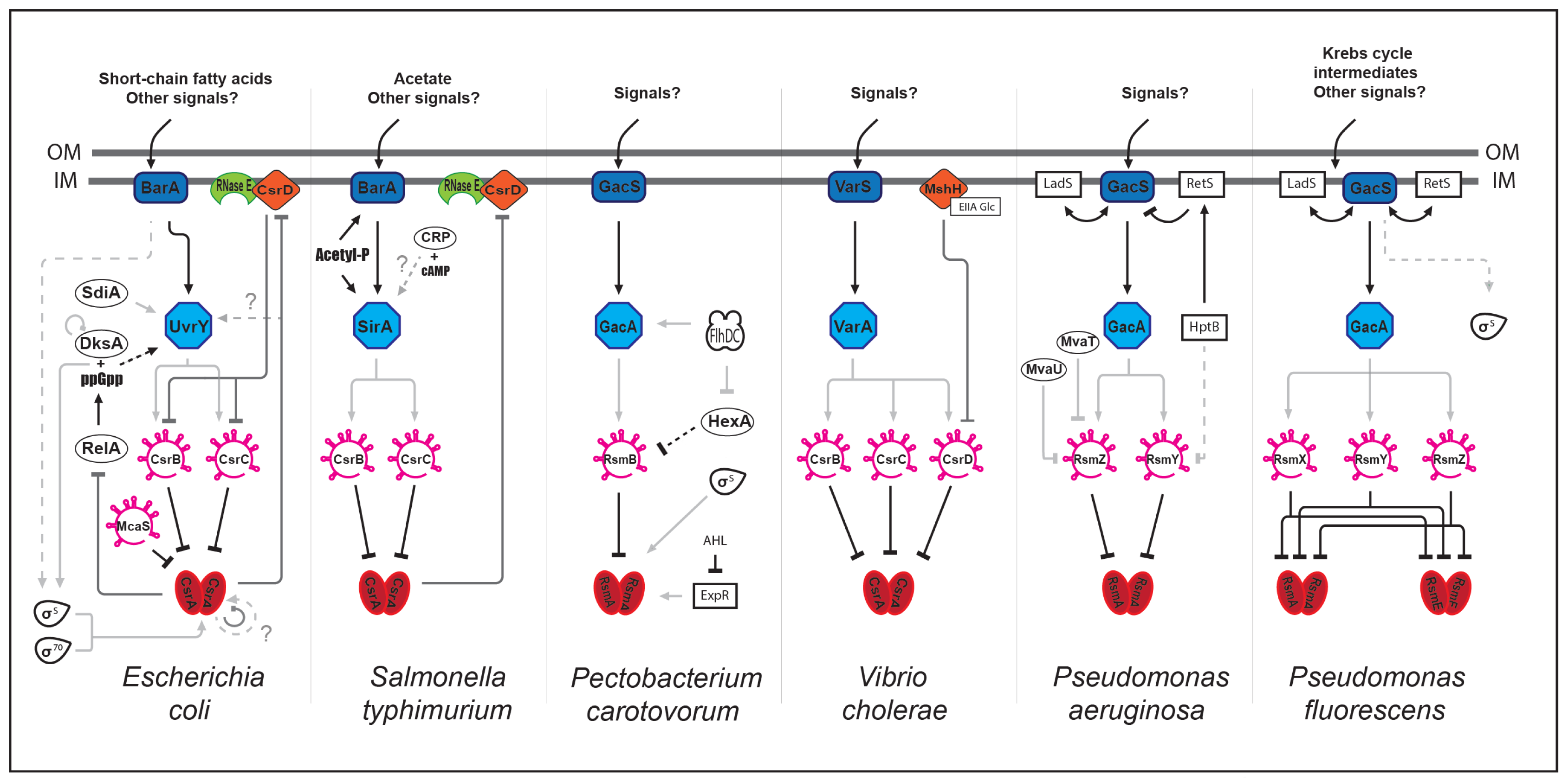
2. The Csr Network
2.1. CsrA Is a Global Regulator
2.2. Building up the Csr Network
2.3. Expanding the Csr Network
2.4. Are the Regulatory Csr/Rsm sRNAs Redundant?
3. The CsrA-Containing Ribonucleoprotein Complexes
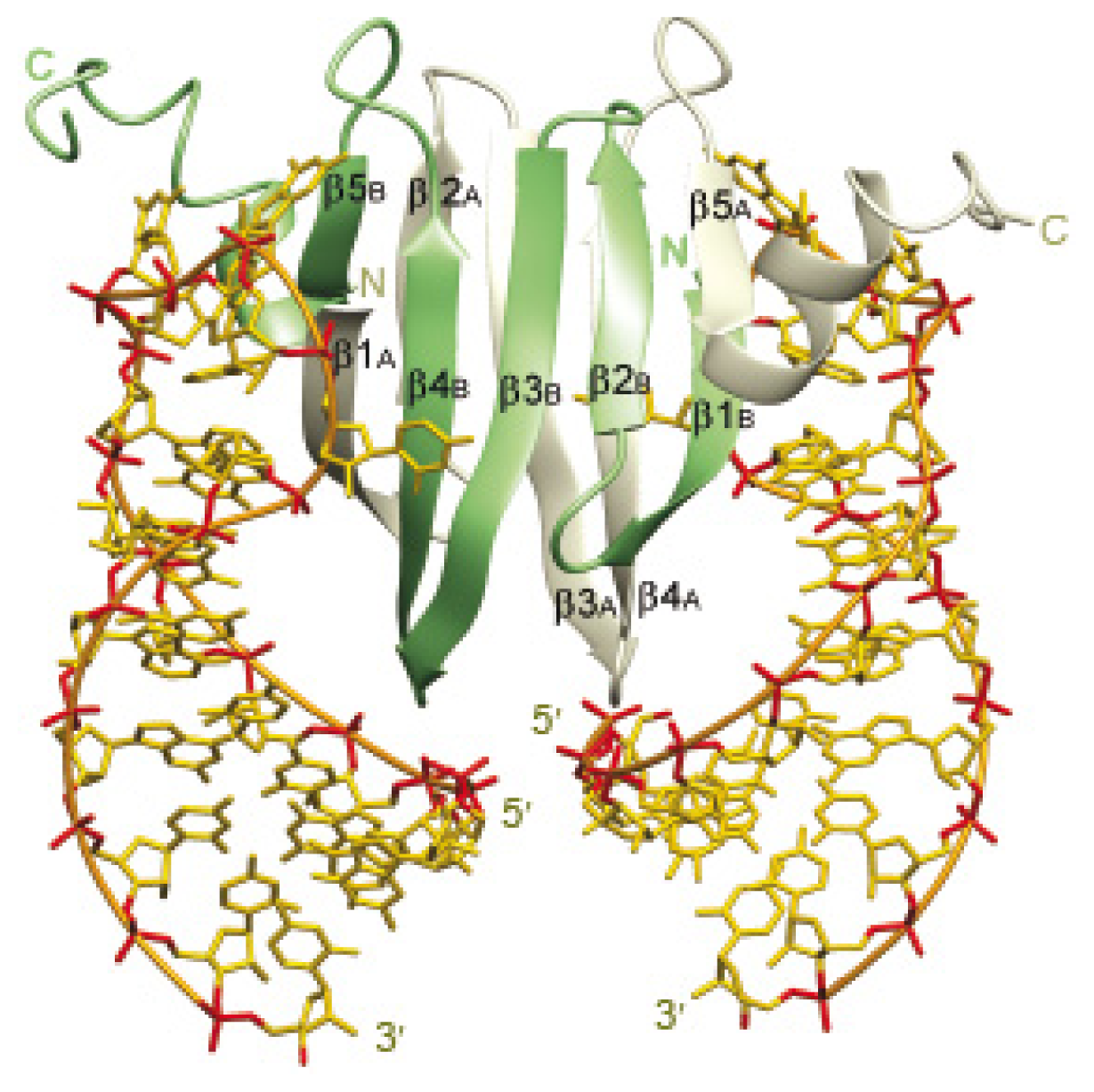
3.1. Structural Information
3.2. Mechanism of Action of CsrA on mRNAs Targets
3.3. Deeper Understanding of the Ribonucleoprotein Complexes Formed by CsrA
4. General Conclusions


Acknowledgments
Conflicts of Interest
References
- Byrgazov, K.; Vesper, O.; Moll, I. Ribosome heterogeneity: Another level of complexity in bacterial translation regulation. Curr. Opin. Microbiol 2013, 16, 133–139. [Google Scholar]
- Babitzke, P.; Baker, C.S.; Romeo, T. Regulation of translation initiation by RNA binding proteins. Annu. Rev. Microbiol 2009, 63, 27–44. [Google Scholar]
- Wassarman, K.M.; Storz, G. 6S RNA regulates E. coli RNA polymerase activity. Cell 2000, 101, 613–623. [Google Scholar]
- Grudnik, P.; Bange, G.; Sinning, I. Protein targeting by the signal recognition particle. Biol. Chem 2009, 390, 775–782. [Google Scholar]
- Vogel, J.; Luisi, B.F. Hfq and its constellation of RNA. Nat. Rev. Microbiol 2011, 9, 578–589. [Google Scholar]
- Jiang, W.; Hou, Y.; Inouye, M. CspA, the major cold-shock protein of Escherichia coli, is an RNA chaperone. J. Biol. Chem 1997, 272, 196–202. [Google Scholar]
- Pichon, C.; Felden, B. Proteins that interact with bacterial small RNA regulators. FEMS Microbiol. Rev 2007, 31, 614–625. [Google Scholar] [Green Version]
- Romeo, T.; Gong, M.; Liu, M.Y.; Brun-Zinkernagel, A.M. Identification and molecular characterization of csrA, a pleiotropic gene from Escherichia coli that affects glycogen biosynthesis, gluconeogenesis, cell size, and surface properties. J. Bacteriol 1993, 175, 4744–4755. [Google Scholar]
- Baker, C.S.; Morozov, I.; Suzuki, K.; Romeo, T.; Babitzke, P. CsrA regulates glycogen biosynthesis by preventing translation of glgC in Escherichia coli. Mol. Microbiol 2002, 44, 1599–1610. [Google Scholar]
- Dubey, A.K.; Baker, C.S.; Suzuki, K.; Jones, A.D.; Pandit, P.; Romeo, T.; Babitzke, P. CsrA regulates translation of the Escherichia coli carbon starvation gene, cstA, by blocking ribosome access to the cstA transcript. J. Bacteriol 2003, 185, 4450–4460. [Google Scholar]
- Yang, H. Pleiotropic regulation of central carbohydrate metabolism in Escherichia coli via the gene csrA. J. Biol. Chem. 1995, 270, 29096–29104. [Google Scholar]
- McKee, A.E.; Rutherford, B.J.; Chivian, D.C.; Baidoo, E.K.; Juminaga, D.; Kuo, D.; Benke, P.I.; Dietrich, J.A.; Ma, S.M.; Arkin, A.P.; et al. Manipulation of the carbon storage regulator system for metabolite remodeling and biofuel production in Escherichia coli. Microb. Cell Fact 2012, 11, 79. [Google Scholar]
- Wei, B.; Shin, S.; LaPorte, D.; Wolfe, A.J.; Romeo, T. Global regulatory mutations in csrA and rpoS cause severe central carbon stress in Escherichia coli in the presence of acetate. J. Bacteriol 2000, 182, 1632–1640. [Google Scholar]
- Lawhon, S.D.; Frye, J.G.; Suyemoto, M.; Porwollik, S.; McClelland, M.; Altier, C. Global regulation by CsrA inSalmonella typhimurium. Mol. Microbiol 2003, 48, 1633–1645. [Google Scholar]
- Jackson, D.W.; Suzuki, K.; Oakford, L.; Simecka, J.W.; Hart, M.E.; Romeo, T. Biofilm formation and dispersal under the influence of the global regulator CsrA of Escherichia coli. J. Bacteriol 2002, 184, 290–301. [Google Scholar]
- Wang, X.; Preston, J.F.; Romeo, T. The pgaABCD locus of Escherichia coli promotes the synthesis of a polysaccharide adhesin required for biofilm formation. J. Bacteriol 2004, 186, 2724–2734. [Google Scholar]
- Wang, X.; Dubey, A.K.; Suzuki, K.; Baker, C.S.; Babitzke, P.; Romeo, T. CsrA post-transcriptionally represses pgaABCD, responsible for synthesis of a biofilm polysaccharide adhesin of Escherichia coli. Mol. Microbiol 2005, 56, 1648–1663. [Google Scholar]
- Wei, B.L.; Brun-Zinkernagel, A.M.; Simecka, J.W.; Pruss, B.M.; Babitzke, P.; Romeo, T. Positive regulation of motility and flhDC expression by the RNA-binding protein CsrA of Escherichia coli. Mol. Microbiol 2001, 40, 245–256. [Google Scholar]
- Yakhnin, A.V.; Baker, C.S.; Vakulskas, C.A.; Yakhnin, H.; Berezin, I.; Romeo, T.; Babitzke, P. CsrA activates flhDC expression by protecting flhDC mRNA from RNase E-mediated cleavage. Mol. Microbiol 2013, 87, 851–866. [Google Scholar]
- Boyd, C.D.; O’Toole, G.A. Second messenger regulation of biofilm formation: Breakthroughs in understanding c-di-GMP effector systems. Annu. Rev. Cell Dev. Biol 2012, 28, 439–462. [Google Scholar]
- Jonas, K.; Edwards, A.N.; Simm, R.; Romeo, T.; Romling, U.; Melefors, O. The RNA binding protein CsrA controls cyclic di-GMP metabolism by directly regulating the expression of GGDEF proteins. Mol. Microbiol 2008, 70, 236–257. [Google Scholar]
- Cui, Y.; Chatterjee, A.; Liu, Y.; Dumenyo, C.K.; Chatterjee, A.K. Identification of a global repressor gene, rsmA, of Erwinia carotovora subsp. carotovora that controls extracellular enzymes, N-(3-oxohexanoyl)-l-homoserine lactone, and pathogenicity in soft-rotting Erwinia spp. J. Bacteriol 1995, 177, 5108–5115. [Google Scholar]
- Goodman, A.L.; Kulasekara, B.; Rietsch, A.; Boyd, D.; Smith, R.S.; Lory, S. A signaling network reciprocally regulates genes associated with acute infection and chronic persistence in Pseudomonas aeruginosa. Dev. Cell 2004, 7, 745–754. [Google Scholar]
- Molofsky, A.B.; Swanson, M.S. Legionella pneumophila CsrA is a pivotal repressor of transmission traits and activator of replication. Mol. Microbiol 2003, 50, 445–461. [Google Scholar]
- Martínez, L.C.; Yakhnin, H.; Camacho, M.I.; Georgellis, D.; Babitzke, P.; Puente, J.L.; Bustamante, V.H. Integration of a complex regulatory cascade involving the SirA/BarA and Csr global regulatory systems that controls expression of the Salmonella SPI-1 and SPI-2 virulence regulons through HilD. Mol. Microbiol 2011, 80, 1637–1656. [Google Scholar]
- Kõiv, V.; Andresen, L.; Broberg, M.; Frolova, J.; Somervuo, P.; Auvinen, P.; Pirhonen, M.; Tenson, T.; Mäe, A. Lack of RsmA-mediated control results in constant hypervirulence, cell elongation, and hyperflagellation in Pectobacterium wasabiae. PLoS One 2013, 8, e54248. [Google Scholar]
- Pessi, G.; Haas, D. Dual control of hydrogen cyanide biosynthesis by the global activator GacA in Pseudomonas aeruginosa PAO1. FEMS Microbiol. Lett 2001, 200, 73–78. [Google Scholar]
- Brencic, A.; Lory, S. Determination of the regulon and identification of novel mRNA targets of Pseudomonas aeruginosa RsmA. Mol. Microbiol 2009, 72, 612–632. [Google Scholar]
- Heurlier, K.; Pessi, G.; Williams, F.; Holden, M.T.G.; Mara, M.C.A.; Haas, D.; Williams, P. The global posttranscriptional regulator RsmA modulates production of virulence determinants and N-Acylhomoserine lactones in Pseudomonas aeruginosa. 2001, 183, 6676–6683. [Google Scholar]
- Heeb, S.; Blumer, C.; Haas, D. Regulatory RNA as mediator in GacA/RsmA-dependent global control of exoproduct formation in Pseudomonas regulatory RNA as mediator in GacA/RsmA-dependent global control of exoproduct formation in Pseudomonas fluorescens CHA0. J. Bacteriol 2002, 184, 1046–1056. [Google Scholar]
- Sacherer, P.; Défago, G.; Haas, D. Extracellular protease and phospholipase C are controlled by the global regulatory gene gacA in the biocontrol strain Pseudomonas fluorescens CHA0. FEMS Microbiol. Lett 1994, 116, 155–160. [Google Scholar]
- Timmermans, J.; van Melderen, L. Conditional essentiality of the csrA gene in Escherichia coli. J. Bacteriol 2009, 191, 1722–1724. [Google Scholar]
- Altier, C.; Suyemoto, M.; Lawhon, S.D. Regulation of Salmonella enterica serovar typhimurium invasion genes by csrA. Infect. Immun 2000, 68, 6790–6797. [Google Scholar]
- Mitra, A.; Palaniyandi, S.; Herren, C.D.; Zhu, X.; Mukhopadhyay, S. Pleiotropic roles of uvrY on biofilm formation, motility and virulence in uropathogenic Escherichia coli CFT073. PLoS One 2013, 8, e55492. [Google Scholar]
- Seyll, E.; Van Melderen, L. Université Libre de Bruxelles (ULB), Laboratoire de Génétique et Physiologie Bactérienne: Belgium; Unpublished work; 2013. [Google Scholar]
- Gutierrez, P.; Li, Y.; Osborne, M.J.; Pomerantseva, E.; Liu, Q.; Gehring, K. Solution structure of the carbon storage regulator protein CsrA from Escherichia coli. J. Bacteriol 2005, 187, 3496–3501. [Google Scholar]
- Liu, M.Y.; Gui, G.; Wei, B.; Preston, J.F., 3rd; Oakford, L.; Yuksel, U.; Giedroc, D.P.; Romeo, T. The RNA molecule CsrB binds to the global regulatory protein CsrA and antagonizes its activity in Escherichia coli. J. Biol. Chem 1997, 272, 17502–17510. [Google Scholar]
- Weilbacher, T.; Suzuki, K.; Dubey, A.K.; Wang, X.; Gudapaty, S.; Morozov, I.; Baker, C.S.; Georgellis, D.; Babitzke, P.; Romeo, T. A novel sRNA component of the carbon storage regulatory system of Escherichia coli. Mol. Microbiol 2003, 48, 657–670. [Google Scholar]
- Romeo, T. Global regulation by the small RNA-binding protein CsrA and the non-coding RNA molecule CsrB. Mol. Microbiol 1998, 29, 1321–1330. [Google Scholar]
- Babitzke, P.; Romeo, T. CsrB sRNA family: Sequestration of RNA-binding regulatory proteins. Curr. Opin. Microbiol 2007, 10, 156–163. [Google Scholar]
- Gudapaty, S.; Suzuki, K.; Wang, X.; Babitzke, P.; Romeo, T. Regulatory interactions of Csr components: The RNA binding protein CsrA activates csrB transcription in Escherichia coli. J. Bacteriol 2001, 183, 6017–6027. [Google Scholar]
- Schubert, M.; Lapouge, K.; Duss, O.; Oberstrass, F.C.; Jelesarov, I.; Haas, D.; Allain, F.H.-T. Molecular basis of messenger RNA recognition by the specific bacterial repressing clamp RsmA/CsrA. Nat. Struct. Mol. Biol 2007, 14, 807–813. [Google Scholar]
- Suzuki, K.; Wang, X.; Weilbacher, T.; Pernestig, A.K.; Melefors, O.; Georgellis, D.; Babitzke, P.; Romeo, T. Regulatory circuitry of the CsrA/CsrB and BarA/UvrY systems of Escherichia coli. J. Bacteriol 2002, 184, 5130–5140. [Google Scholar]
- Burrowes, E.; Abbas, A.; O’Neill, A.; Adams, C.; O’Gara, F. Characterisation of the regulatory RNA RsmB from Pseudomonas aeruginosa PAO1. Res. Microbiol 2005, 156, 7–16. [Google Scholar]
- Heurlier, K.; Williams, F.; Heeb, S.; Dormond, C.; Pessi, G.; Singer, D.; Camara, M.; Williams, P.; Haas, D. Positive control of swarming, rhamnolipid synthesis, and lipase production by the posttranscriptional RsmA/RsmZ system in Pseudomonas aeruginosa PAO1. J. Bacteriol 2004, 186, 2936–2945. [Google Scholar]
- Yakhnin, H.; Yakhnin, A.V.; Baker, C.S.; Sineva, E.; Berezin, I.; Romeo, T.; Babitzke, P. Complex regulation of the global regulatory gene csrA: CsrA-mediated translational repression, transcription from five promoters by Esigma(7)(0) and Esigma(S), and indirect transcriptional activation by CsrA. Mol. Microbiol 2011, 81, 689–704. [Google Scholar]
- Takeuchi, K.; Kiefer, P.; Reimmann, C.; Keel, C.; Dubuis, C.; Rolli, J.; Vorholt, J.A.; Haas, D. Small RNA-dependent expression of secondary metabolism is controlled by Krebs cycle function in Pseudomonas fluorescens. J. Biol. Chem 2009, 284, 34976–34985. [Google Scholar]
- Chavez, R.G.; Alvarez, A.F.; Romeo, T.; Georgellis, D. The physiological stimulus for the BarA sensor kinase. J. Bacteriol 2010, 192, 2009–2012. [Google Scholar]
- Suzuki, K.; Babitzke, P.; Kushner, S.R.; Romeo, T. Identification of a novel regulatory protein (CsrD) that targets the global regulatory RNAs CsrB and CsrC for degradation by RNase E. Genes Dev 2006, 20, 2605–2617. [Google Scholar]
- Jonas, K.; Edwards, A.N.; Ahmad, I.; Romeo, T.; Römling, U.; Melefors, O. Complex regulatory network encompassing the Csr, c-di-GMP and motility systems of Salmonella Typhimurium. Environ. Microbiol 2010, 12, 524–540. [Google Scholar]
- Mukherjee, S.; Yakhnin, H.; Kysela, D.; Sokoloski, J.; Babitzke, P.; Kearns, D.B. CsrA-FliW interaction governs flagellin homeostasis and a checkpoint on flagellar morphogenesis in Bacillus subtilis. Mol. Microbiol 2011, 82, 447–461. [Google Scholar]
- Reimmann, C.; Valverde, C.; Kay, E.; Haas, D. Posttranscriptional repression of GacS/GacA-controlled genes by the RNA-binding protein RsmE acting together with RsmA in the biocontrol strain Pseudomonas fluorescens CHA0. J. Bacteriol 2005, 187, 276–285. [Google Scholar]
- Lenz, D.H.; Miller, M.B.; Zhu, J.; Kulkarni, R.V.; Bassler, B.L. CsrA and three redundant small RNAs regulate quorum sensing in Vibrio cholerae. Mol. Microbiol 2005, 58, 1186–1202. [Google Scholar]
- Kay, E.; Dubuis, C.; Haas, D. Three small RNAs jointly ensure secondary metabolism and biocontrol in Pseudomonas fluorescens CHA0. Proc. Natl. Acad. Sci. USA 2005, 102, 17136–17141. [Google Scholar]
- Edwards, A.N.; Patterson-Fortin, L.M.; Vakulskas, C.A.; Mercante, J.W.; Potrykus, K.; Vinella, D.; Camacho, M.I.; Fields, J.A.; Thompson, S.A.; Georgellis, D.; et al. Circuitry linking the Csr and stringent response global regulatory systems. Mol. Microbiol 2011, 80, 1561–1580. [Google Scholar]
- Cui, Y.; Chatterjee, A.; Hasegawa, H.; Dixit, V.; Leigh, N.; Chatterjee, A.K. ExpR, a LuxR homolog of Erwinia carotovora subsp. carotovora, activates transcription of rsmA, which specifies a global regulatory RNA-binding protein. J. Bacteriol 2005, 187, 4792–4803. [Google Scholar]
- Pickering, B.S.; Smith, D.R.; Watnick, P.I. Glucose-specific enzyme IIA has unique binding partners in the Vibrio cholerae biofilm. MBio 2012, 3, e00228–e00212. [Google Scholar]
- Altier, C.; Suyemoto, M.; Ruiz, A.I.; Burnham, K.D.; Maurer, R. Characterization of two novel regulatory genes affecting Salmonella invasion gene expression. Mol. Microbiol 2000, 35, 635–646. [Google Scholar]
- Lawhon, S.D.; Maurer, R.; Suyemoto, M.; Altier, C. Intestinal short-chain fatty acids alter Salmonella typhimurium invasion gene expression and virulence through BarA/SirA. Mol. Microbiol 2002, 46, 1451–1464. [Google Scholar]
- Teplitski, M.; Goodier, R.I.; Ahmer, B.M. Pathways leading from BarA/SirA to motility and virulence gene expression in Salmonella. J. Bacteriol 2003, 185, 7257–7265. [Google Scholar]
- Teplitski, M.; Goodier, R.I.; Ahmer, B.M. Catabolite repression of the SirA regulatory cascade in Salmonella enterica. Int. J. Med. Microbiol 2006, 296, 449–466. [Google Scholar]
- Cui, Y.; Chatterjee, A.; Yang, H.; Chatterjee, A.K. Regulatory network controlling extracellular proteins in Erwinia carotovora subsp. carotovora: flhDC, the master regulator of flagellar genes, activates rsmB regulatory RNA production by affecting gacA and hexA (lrhA) expression. J. Bacteriol 2008, 190, 4610–4623. [Google Scholar]
- Mukherjee, A.; Cui, Y.; Ma, W.; Liu, Y.; Chatterjee, A.K. hexA of Erwinia carotovora ssp. carotovora strain Ecc71 negatively regulates production of RpoS and rsmB RNA, a global regulator of extracellular proteins, plant virulence and the quorum-sensing signal, N-(3-oxohexanoyl)-l-homoserine lactone. Environ. Microbiol 2000, 2, 203–215. [Google Scholar]
- Ventre, I.; Goodman, A.L.; Vallet-Gely, I.; Vasseur, P.; Soscia, C.; Molin, S.; Bleves, S.; Lazdunski, A.; Lory, S.; Filloux, A. Multiple sensors control reciprocal expression of Pseudomonas aeruginosa regulatory RNA and virulence genes. Proc. Natl. Acad. Sci. USA 2006, 103, 171–176. [Google Scholar]
- Workentine, M.L.; Chang, L.; Ceri, H.; Turner, R.J. The GacS-GacA two-component regulatory system of Pseudomonas fluorescens: A bacterial two-hybrid analysis. FEMS Microbiol. Lett 2009, 292, 50–56. [Google Scholar]
- Goodman, A.L.; Merighi, M.; Hyodo, M.; Ventre, I.; Filloux, A.; Lory, S. Direct interaction between sensor kinase proteins mediates acute and chronic disease phenotypes in a bacterial pathogen. Genes Dev 2009, 23, 249–259. [Google Scholar]
- Jorgensen, M.G.; Thomason, M.K.; Havelund, J.; Valentin-Hansen, P.; Storz, G. Dual function of the McaS small RNA in controlling biofilm formation. Genes Dev 2013, 27, 1132–1145. [Google Scholar]
- Mukherjee, A.; Cui, Y.; Ma, W.; Liu, Y.; Ishihama, A.; Eisenstark, A.; Chatterjee, A.K. RpoS (sigma-S) controls expression of rsmA, a global regulator of secondary metabolites, harpin, and extracellular proteins in Erwinia carotovora. J. Bacteriol 1998, 180, 3629–3634. [Google Scholar]
- Valverde, C.; Heeb, S.; Keel, C.; Haas, D. RsmY, a small regulatory RNA, is required in concert with RsmZ for GacA-dependent expression of biocontrol traits in Pseudomonas fluorescens CHA0. Mol. Microbiol 2003, 50, 1361–1379. [Google Scholar]
- Fortune, D.R.; Suyemoto, M.; Altier, C. Identification of CsrC and characterization of its role in epithelial cell invasion in Salmonella enterica serovar Typhimurium. Infect. Immun 2006, 74, 331–339. [Google Scholar]
- Mackie, G.A. RNase E: At the interface of bacterial RNA processing and decay. Nat. Rev. Microbiol 2013, 11, 45–57. [Google Scholar]
- Heroven, A.K.; Sest, M.; Pisano, F.; Scheb-Wetzel, M.; Steinmann, R.; Bohme, K.; Klein, J.; Munch, R.; Schomburg, D.; Dersch, P. Crp induces switching of the CsrB and CsrC RNAs in Yersinia pseudotuberculosis and links nutritional status to virulence. Front. Cell. Infect. Microbiol 2012, 2, 158. [Google Scholar]
- Brencic, A.; McFarland, K.A.; McManus, H.R.; Castang, S.; Mogno, I.; Dove, S.L.; Lory, S. The GacS/GacA signal transduction system of Pseudomonas aeruginosa acts exclusively through its control over the transcription of the RsmY and RsmZ regulatory small RNAs. Mol. Microbiol 2009, 73, 434–445. [Google Scholar]
- Bordi, C.; Lamy, M.C.; Ventre, I.; Termine, E.; Hachani, A.; Fillet, S.; Roche, B.; Bleves, S.; Mejean, V.; Lazdunski, A.; et al. Regulatory RNAs and the HptB/RetS signalling pathways fine-tune Pseudomonas aeruginosa pathogenesis. Mol. Microbiol 2010, 76, 1427–1443. [Google Scholar]
- Sonnleitner, E.; Schuster, M.; Sorger-Domenigg, T.; Greenberg, E.P.; Blasi, U. Hfq-dependent alterations of the transcriptome profile and effects on quorum sensing in Pseudomonas aeruginosa. Mol. Microbiol 2006, 59, 1542–1558. [Google Scholar]
- Liu, M.Y.; Yang, H.; Romeo, T. The product of the pleiotropic Escherichia coli gene csrA modulates glycogen biosynthesis via effects on mRNA stability. J. Bacteriol 1995, 177, 2663–2672. [Google Scholar]
- Heeb, S.; Kuehne, S.A.; Bycroft, M.; Crivii, S.; Allen, M.D.; Haas, D.; Cámara, M.; Williams, P. Functional analysis of the post-transcriptional regulator RsmA reveals a novel RNA-binding site. J. Mol. Biol 2006, 355, 1026–1036. [Google Scholar]
- Mercante, J.; Suzuki, K.; Cheng, X.; Babitzke, P.; Romeo, T. Comprehensive alanine-scanning mutagenesis of Escherichia coli CsrA defines two subdomains of critical functional importance. J. Biol. Chem 2006, 281, 31832–31842. [Google Scholar]
- Dubey, A.K.; Baker, C.S.; Romeo, T.; Babitzke, P. RNA sequence and secondary structure participate in high-affinity CsrA-RNA interaction. RNA 2005, 11, 1579–1587. [Google Scholar]
- Baker, C.S.; Eory, L.A.; Yakhnin, H.; Mercante, J.; Romeo, T.; Babitzke, P. CsrA inhibits translation initiation of Escherichia coli hfq by binding to a single site overlapping the Shine-Dalgarno sequence. J. Bacteriol 2007, 189, 5472–5481. [Google Scholar]
- Yakhnin, H.; Baker, C.S.; Berezin, I.; Evangelista, M.A.; Rassin, A.; Romeo, T.; Babitzke, P. CsrA represses translation of sdiA, which encodes the N-acylhomoserine-l-lactone receptor of Escherichia coli, by binding exclusively within the coding region of sdiA mRNA. J. Bacteriol 2011, 193, 6162–6170. [Google Scholar]
- Patterson-Fortin, L.M.; Vakulskas, C.A.; Yakhnin, H.; Babitzke, P.; Romeo, T. Dual posttranscriptional regulation via a cofactor-responsive mRNA leader. J. Mol. Biol 2013, 425, 3662–3677. [Google Scholar]
- Mercante, J.; Edwards, A.N.; Dubey, A.K.; Babitzke, P.; Romeo, T. Molecular geometry of CsrA (RsmA) binding to RNA and its implications for regulated expression. J. Mol. Biol 2009, 392, 511–528. [Google Scholar]
- Mitobe, J.; Yanagihara, I.; Ohnishi, K.; Yamamoto, S.; Ohnishi, M.; Ishihama, A.; Watanabe, H. RodZ regulates the post-transcriptional processing of the Shigella sonnei type III secretion system. EMBO Rep 2011, 12, 911–916. [Google Scholar]
© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Seyll, E.; Van Melderen, L. The Ribonucleoprotein Csr Network. Int. J. Mol. Sci. 2013, 14, 22117-22131. https://doi.org/10.3390/ijms141122117
Seyll E, Van Melderen L. The Ribonucleoprotein Csr Network. International Journal of Molecular Sciences. 2013; 14(11):22117-22131. https://doi.org/10.3390/ijms141122117
Chicago/Turabian StyleSeyll, Ethel, and Laurence Van Melderen. 2013. "The Ribonucleoprotein Csr Network" International Journal of Molecular Sciences 14, no. 11: 22117-22131. https://doi.org/10.3390/ijms141122117
APA StyleSeyll, E., & Van Melderen, L. (2013). The Ribonucleoprotein Csr Network. International Journal of Molecular Sciences, 14(11), 22117-22131. https://doi.org/10.3390/ijms141122117