S-Layer Protein Self-Assembly



Abstract
:1. Introduction
2. Occurrence and Location of S-Layers
3. Ultrastructure of S-Layers
4. Genetic Engineering of S-Layer Proteins
5. Reassembly of S-Layer Proteins
5.1. Isolation Procedures
5.2. Reassembly in Solution
5.3. Reassembly on Mica and on Silicon Substrates
5.4. Reassembly on Self-Assembled Monolayers
5.5. Reassembly on Polyelectrolyte Layers
5.6. Reassembly at Lipid Interfaces
5.7. Reassembly at Liposomes and Nanocapsules
6. Summary
Acknowledgments
Conflict of Interest
References
- Sleytr, U.B. Regular arrays of macromolecules on bacterial cell walls: Structure, chemistry, assembly, and function. Int. Rev. Cytol 1978, 53, 1–64. [Google Scholar]
- Sleytr, U.B.; Glauert, A.M. Analysis of regular arrays of subunits on bacterial surfaces: Evidence for a dynamic process of assembly. J. Ultrastruct. Res 1975, 50, 103–116. [Google Scholar]
- Sleytr, U.B.; Beveridge, T.J. Bacterial S-layers. Trends Microbiol 1999, 7, 253–260. [Google Scholar]
- Sleytr, U.B.; Egelseer, E.M.; Ilk, N.; Messner, P.; Schäffer, C.; Pum, D.; Schuster, B. Nanobiotechnological Applications of S-Layers. In Prokaryotic Cell Wall Compounds—Structure and Biochemistry; König, H., Claus, H., Varma, A., Eds.; Springer-Verlag: Berlin, Germany, 2010; pp. 459–481. [Google Scholar]
- Sleytr, U.B.; Messner, P. Crystalline Cell Surface Layers (S layers). In Encyclopedia of Microbiology; Schaechter, M., Ed.; Academic Press/Elsevier Science: San Diego, CA, USA, 2009; Volume 1, pp. 89–98. [Google Scholar]
- Sleytr, U.B. Self-assembly of the hexagonally and tetragonally arranged subunits of bacterial surface layers and their reattachment to cell walls. J. Ultrastruct. Res 1976, 55, 360–377. [Google Scholar]
- Sleytr, U.B.; Messner, P.; Pum, D.; Sára, M. Crystalline bacterial cell surface layers (S layers): From supramolecular cell structure to biomimetics and nanotechnology. Angew. Chem. Int. Ed 1999, 38, 1035–1054. [Google Scholar]
- Sleytr, U.B.; Schuster, B.; Egelseer, E.M.; Pum, D.; Horejs, C.M.; Tscheliessnig, R.; Ilk, N. Nanobiotechnology with S-layer proteins as building blocks. Prog. Mol. Biol. Transl. Sci 2011, 103, 277–352. [Google Scholar]
- Sleytr, U.B.; Huber, C.; Ilk, N.; Pum, D.; Schuster, B.; Egelseer, E.M. S-layers as a tool kit for nanobiotechnological applications. FEMS Microbiol. Lett 2007, 267, 131–144. [Google Scholar]
- Sleytr, U.B.; Egelseer, E.M.; Ilk, N.; Pum, D.; Schuster, B. S-layers as a basic building block in a molecular construction kit. FEBS J 2007, 274, 323–334. [Google Scholar]
- Sleytr, U.B.; Sára, M.; Pum, D.; Schuster, B. Crystalline Bacterial Cell Surface Layers (S-Layers): A Versatile Self-Assembly System. In Supramolecular Polymers, 2nd ed; Ciferri, A., Ed.; Taylor and Francis: Boca Raton, FL, USA, 2005; pp. 583–616. [Google Scholar]
- Sleytr, U.B.; Sára, M.; Pum, D.; Schuster, B. Molecular Nanotechnology and Nanobiotechnology with Two-Dimensional Protein Crystals (S-Layers). In Nano-Surface Chemistry; Rosoff, M., Ed.; Marcel Dekker, Inc: New York, NY, USA, 2001; pp. 333–389. [Google Scholar]
- Shenton, W.; Pum, D.; Sleytr, U.B.; Mann, S. Synthesis of cadmium sulphide superlattices using self-assembled bacterial S-layers. Nature 1997, 389, 585–587. [Google Scholar]
- Dieluweit, S.; Pum, D.; Sleytr, U.B. Formation of a gold superlattice on an S-layer with square lattice symmetry. Supramol. Sci 1998, 5, 15–19. [Google Scholar]
- Sára, M.; Sleytr, U.B. S-layer proteins. J. Bacteriol 2000, 182, 859–868. [Google Scholar]
- Claus, H.; König, H. Cell Envelopes of Methanogens. In Prokaryotic Cell Wall Compounds—Structure and Biochemistry; König, H., Claus, H., Varma, A., Eds.; Springer-Verlag: Berlin, Germany, 2010; pp. 231–251. [Google Scholar]
- Eichler, J.; Abu-Qarn, M.; Konrad, Z.; Magidovich, H.; Plavner, N.; Yurist-Doutsch, S. The Cell Envelopes of Haloarchaea: Staying in Shape in a World of Salt. In Prokaryotic Cell Wall Compounds—Structure and Biochemistry; König, H., Claus, H., Varma, A., Eds.; Springer-Verlag: Berlin, Germany, 2010; pp. 253–270. [Google Scholar]
- Rachel, R. Cell Envelopes of Crenarchaeota and Nanoarchaeota. In Prokaryotic Cell Wall Compounds—Structure and Biochemistry; König, H., Claus, H., Varma, A., Eds.; Springer-Verlag: Berlin, Germany, 2010; pp. 271–291. [Google Scholar]
- Beveridge, T.J.; Sára, M.; Pum, D.; Sprott, G.D.; Stewart, M.; Sleytr, U.B. The Structure, Chemistry and Physicochemistry of the Methanospirillum Hungatei GP1 Sheath. In Crystalline Bacterial Cell Surface Layers; Sleytr, U.B., Messner, P., Pum, D., Sára, M., Eds.; Springer-Verlag: Berlin, Germany, 1988; pp. 26–30. [Google Scholar]
- Messner, P.; Pum, D.; Sára, M.; Stetter, K.O.; Sleytr, U.B. Ultrastructure of the cell envelope of the archaebacteria Thermoproteus tenax and Thermoproteus neutrophilus. J. Bacteriol 1986, 166, 1046–1054. [Google Scholar]
- Pum, D.; Messner, P.; Sleytr, U.B. Role of the S layer in morphogenesis and cell division of the archaebacterium Methanocorpusculum sinense. J. Bacteriol 1991, 173, 6865–6873. [Google Scholar]
- Messner, P.; Schäffer, C.; Egelseer, E.M.; Sleytr, U.B. Occurrence, Structure, Chemistry, Genetics, Morphogenesis, and Functions of S-layers. In Prokaryotic Cell Wall Compounds— Structure and Biochemistry; König, H., Claus, H., Varma, A., Eds.; Springer-Verlag: Berlin, Germany, 2010; pp. 53–109. [Google Scholar]
- Sára, M.; Dekitsch, C.; Mayer, H.F.; Egelseer, E.M.; Sleytr, U.B. Influence of the secondary cell wall polymer on the reassembly, recrystallization, and stability properties of the S-layer protein from Bacillus stearothermophilus PV72/p2. J. Bacteriol 1998, 180, 4146–4153. [Google Scholar]
- Huber, C.; Ilk, N.; Rünzler, D.; Egelseer, E.M.; Weigert, S.; Sleytr, U.B.; Sára, M. The three S-layer-like homology motifs of the S-layer protein SbpA of Bacillus sphaericus CCM 2177 are not sufficient for binding to the pyruvylated secondary cell wall polymer. Mol. Microbiol 2005, 55, 197–205. [Google Scholar]
- Lupas, A.; Engelhardt, H.; Peters, J.; Santarius, U.; Volker, V.; Baumeister, W. Domain structure of the Acetogenium kivui surface layer revealed by electron crystallography and sequence analysis. J. Bacteriol 1994, 176, 1224–1233. [Google Scholar]
- Sára, M. Conserved anchoring mechanisms between crystalline cell surface S-layer proteins and secondary cell wall polymers in gram-positive bacteria. Trends Microbiol 2001, 9, 47–49. [Google Scholar]
- Beveridge, T.J.; Pouwels, P.H.; Sára, M.; Kotiranta, A.; Lounatmaa, K.; Kari, K.; Kerosuo, E.; Haapasalo, M.; Egelseer, E.M.; Schocher, I.; et al. Functions of S-layers. FEMS Microbiol. Rev 1997, 20, 99–149. [Google Scholar]
- Mader, C.; Huber, C.; Moll, D.; Sleytr, U.B.; Sára, M. Interaction of the crystalline bacterial cell surface layer protein SbsB and the secondary cell wall polymer of Geobacillus stearothermophilus pv72 assessed by real-time surface plasmon resonance biosensor technology. J. Bacteriol 2004, 186, 1758–1768. [Google Scholar]
- Egelseer, E.M.; Sára, M.; Pum, D.; Schuster, B.; Sleytr, U.B. Genetically Engineered S-Layer Proteins and S-Layer-Specific Heteropolysaccharides as Components of a Versatile Molecular Construction Kit for Applications in Nanobiotechnology. In Nanobiotechnology: Bioinspired Devices and Materials of the Future; Shoseyov, O., Levy, I., Eds.; Humana Press: Totowa, NJ, USA, 2008; pp. 55–86. [Google Scholar]
- Badelt-Lichtblau, H.; Kainz, B.; Völlenkle, C.; Egelseer, E.M.; Sleytr, U.B.; Pum, D.; Ilk, N. Genetic engineering of the S-layer protein Sbpa of Lysinibacillus sphaericus CCM 2177 for the generation of functionalized nanoarrays. Bioconjugate Chem 2009, 20, 895–903. [Google Scholar]
- Sleytr, U.B.; Messner, P. Self-assemblies of crystalline bacterial cell surface layers. Electron Microsc. Subcell. Dyn. 1989, 13–31. [Google Scholar]
- Sleytr, U.B. Heterologous reattachment of regular arrays of glycoproteins on bacterial surfaces. Nature 1975, 257, 400–402. [Google Scholar]
- Amos, L.A.; Henderson, R.; Unwin, P.N.T. 3-dimensional structure determination by electron-microscopy of two-dimensional crystals. Prog. Biophys. Mol. Biol 1982, 39, 183–231. [Google Scholar]
- Baumeister, W.; Engelhardt, H. Three-Dimensional Structure of Bacterial Surface Layers; Harris, JR, Horne, RW, Eds.; Academic Press Inc.: London, UK, 1987; Volume 6, pp. 109–154. [Google Scholar]
- Hovmöller, S.; Sjögren, A.; Wang, D.N. The structure of crystalline bacterial surface layers. Prog. Biophys. Mol. Biol 1988, 51, 131–163. [Google Scholar]
- Pavkov-Keller, T.; Howorka, S.; Keller, W. The structure of bacterial S-layer proteins. Prog. Mol. Biol. Transl. Sci 2011, 103, 73–130. [Google Scholar]
- Sára, M.; Sleytr, U.B. Molecular sieving through S-layers of Bacillus stearothermophilus strains. J. Bacteriol 1987, 169, 4092–4098. [Google Scholar]
- Sára, M.; Sleytr, U.B. Production and characteristics of ultrafiltration membranes with uniform pores from two-dimensional arrays of proteins. J. Membr. Sci 1987, 33, 27–49. [Google Scholar]
- Sára, M.; Pum, D.; Sleytr, U.B. Permeability and charge-dependent adsorption properties of the S-layer lattice from Bacillus coagulans E38–66. J. Bacteriol 1992, 174, 3487–3493. [Google Scholar]
- Sára, M.; Sleytr, U.B. Charge Distribution of S-Layers and Importance of Charged Groups for Morphogenesis and Function. In Crystalline Bacterial Cell Surface Layers; Sleytr, U.B., Messner, P., Pum, D., Sára, M., Eds.; Springer-Verlag: Berlin, Germany, 1988; pp. 105–108. [Google Scholar]
- Györvary, E.; Schroedter, A.; Talapin, D.V.; Weller, H.; Pum, D.; Sleytr, U.B. Formation of nanoparticle arrays on S-layer protein lattices. J. Nanosci. Nanotech 2004, 4, 115–120. [Google Scholar]
- Egelseer, E.; Schocher, I.; Sára, M.; Sleytr, U.B. The S-layer from Bacillus stearothermophilus DSM 2358 functions as an adhesion site for a high-molecular weight amylase. J. Bacteriol 1995, 177, 1444–1451. [Google Scholar]
- Sára, M.; Pum, D.; Schuster, B.; Sleytr, U.B. S-layers as patterning elements for application in nanobiotechnology. J. Nanosci. Nanotech 2005, 5, 1939–1953. [Google Scholar]
- Pum, D.; Sleytr, U.B. S-Layer Proteins for Assembling Ordered Nanoparticle Arrays. In Nanobioelectronics—For Electronics, Biology, and Medicine; Offenhäuser, A., Rinaldi, R., Eds.; Springer Science + Business Media: Berlin, Germany, 2009; pp. 167–180. [Google Scholar]
- Moll, D.; Huber, C.; Schlegel, B.; Pum, D.; Sleytr, U.B.; Sára, M. S-layer-streptavidin fusion proteins as template for nanopatterned molecular arrays. Proc. Natl. Acad. Sci. USA 2002, 99, 14646–14651. [Google Scholar]
- Ilk, N.; Völlenkle, C.; Egelseer, E.M.; Breitwieser, A.; Sleytr, U.B.; Sára, M. Molecular characterization of the S-layer gene, Sbpa, of Bacillus sphaericus CCM 2177 and production of a functional S-layer fusion protein with the ability to recrystallize in a defined orientation while presenting the fused allergen. Appl. Environ. Microbiol 2002, 68, 3251–3260. [Google Scholar]
- Egelseer, E.M.; Ilk, N.; Pum, D.; Messner, P.; Schäffer, C.; Schuster, B.; Sleytr, U.B. S-layers, microbial, biotechnological applications. Encycl. Ind. Biotechnol 2010, 7, 4424–4448. [Google Scholar]
- Ilk, N.; Egelseer, E.M.; Sleytr, U.B. S-layer fusion proteins—Construction principles and applications. Curr. Opin. Biotechnol 2011, 22, 824–831. [Google Scholar]
- Sleytr, U.B.; Messner, P. Crystalline surface layers in procaryotes. J. Bacteriol 1988, 170, 2891–2897. [Google Scholar]
- Schuster, B.; Sleytr, U.B. Nanotechnology with S-Layer Proteins. In Protein Nanotechnology: Protocols, Instrumentation and Applications; Gerard, J., Ed.; Humana Press, Springer: Heidelberg, Germany, 2012. [Google Scholar]
- Ilk, N.; Schumi, C.T.; Bohle, B.; Egelseer, E.M.; Sleytr, U.B. Expression of an endotoxin-free S-layer/allergen fusion protein in gram-positive Bacillus subtilis 1012 for the potential application as vaccines for immunotherapy of atopic allergy. Microb. Cell Fact 2011, 10, 10–16. [Google Scholar]
- Pum, D.; Sleytr, U.B. Large-scale reconstruction of crystalline bacterial surface layer proteins at the air-water interface and on lipids. Thin Solid Films 1994, 244, 882–886. [Google Scholar]
- Pum, D.; Sleytr, U.B. Anisotropic crystal growth of the S-layer of Bacillus sphaericus CCM 2177 at the air/water interface. Colloids Surf. A 1995, 102, 99–104. [Google Scholar]
- Baranova, E.; Fronzes, R.; Garcia-Pino, A.; van Gerven, N.; Papapostolou, D.; Pehau-Arnaudet, G.; Pardon, E.; Steyaert, J.; Howorka, S.; Remaut, H. Sbsb structure and lattice reconstruction unveil Ca2+ triggered S-layer assembly. Nature 2012, 487, 119–124. [Google Scholar]
- Norville, J.E.; Kelly, D.F.; Knight, T.F., Jr; Belcher, A.M.; Walz, T. 7Å projection map of the S-layer protein Sbpa obtained with trehalose-embedded monolayer crystals. J. Struct. Biol. 2007, 160, 313–323. [Google Scholar]
- Sára, M.; Sleytr, U.B. Charge distribution on the S-layer of Bacillus stearothermophilus NRS 1536/3c and importance of charged groups for morphogenesis and function. J. Bacteriol 1987, 169, 2804–2809. [Google Scholar]
- Sleytr, U.B.; Glauert, A.M. Ultrastructure of the cell walls of two closely related clostridia that possess different regular arrays of surface subunits. J. Bacteriol 1976, 126, 869–882. [Google Scholar]
- Sleytr, U.B.; Plohberger, R. The Dynamic Process of Assembly of Two-Dimensional Arrays of Macromolecules on Bacterial Cell Walls. In Electron Microscopy at Molecular Dimensions; Baumeister, W., Vogell, W., Eds.; Springer-Verlag: Berlin, Germany, 1980; pp. 36–47. [Google Scholar]
- Messner, P.; Pum, D.; Sleytr, U.B. Characterization of the ultrastructure and the self-assembly of the surface layer of Bacillus stearothermophilus NRS 2004/3a. J. Ultrastruct. Mol. Struct. Res 1986, 97, 73–88. [Google Scholar]
- Jaenicke, R.; Welsch, R.; Sára, M.; Sleytr, U.B. Stability and self-assembly of the S-layer protein of the cell wall of Bacillus stearothermophilus. Biol. Chem. Hoppe-Seyler 1985, 366, 663–670. [Google Scholar]
- Pum, D.; Sleytr, U.B. Monomolecular reassembly of a crystalline bacterial cell surface layer (S-layer) on untreated and modified silicon surfaces. Supramol. Sci 1995, 2, 193–197. [Google Scholar]
- Györvary, E.S.; Stein, O.; Pum, D.; Sleytr, U.B. Self-assembly and recrystallization of bacterial S-layer proteins at silicon supports imaged in real time by atomic force microscopy. J. Microsc 2003, 212, 300–306. [Google Scholar]
- Moreno-Flores, S.; Kasry, A.; Butt, H.J.; Vavilala, C.; Schmittel, M.; Pum, D.; Sleytr, U.B.; Toca-Herrera, J.L. From native to non-native two-dimensional protein lattices through underlying hydrophilic/hydrophobic nanoprotrusions. Angew. Chem. Int. Ed. Edition 2008, 47, 4707–4710. [Google Scholar]
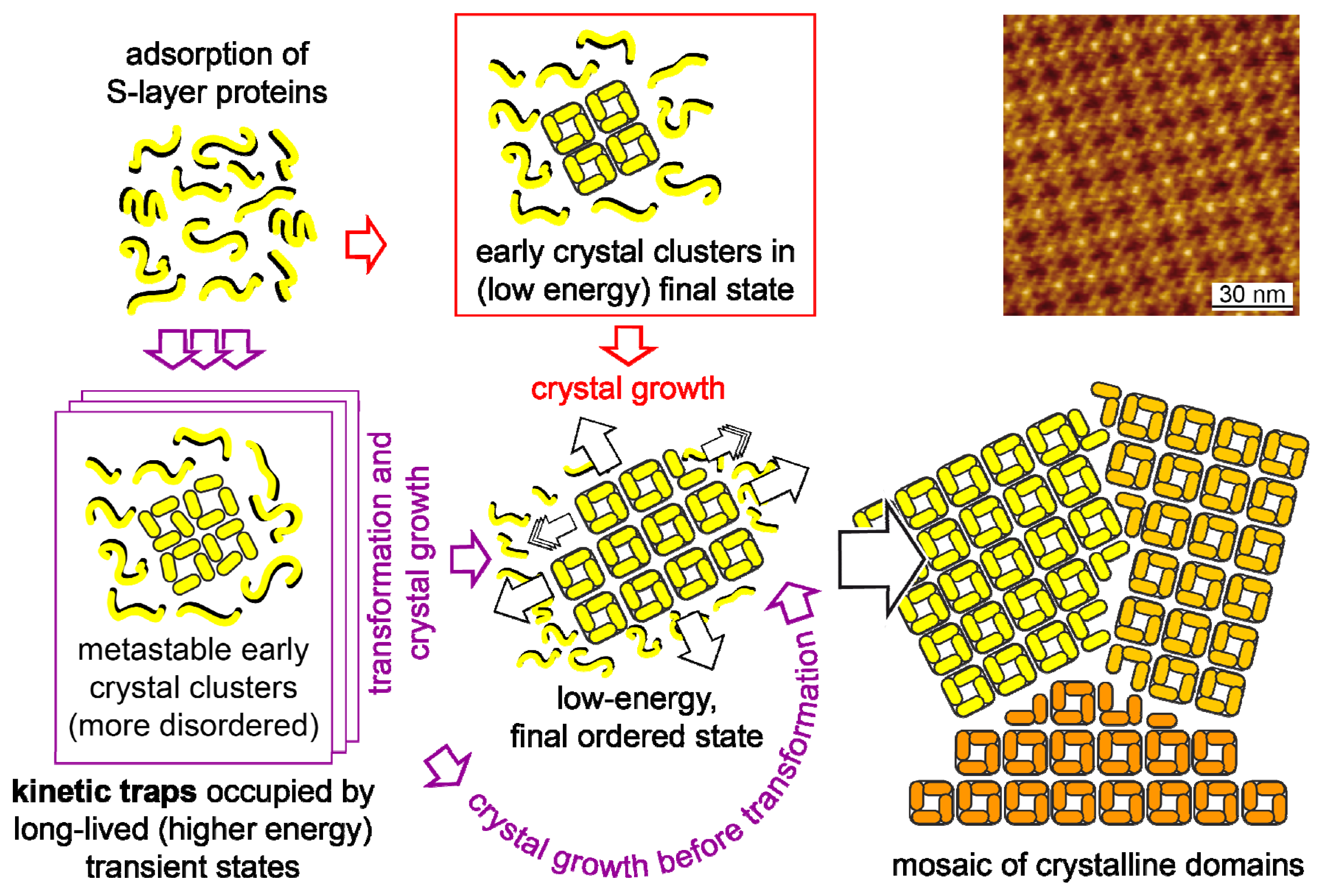
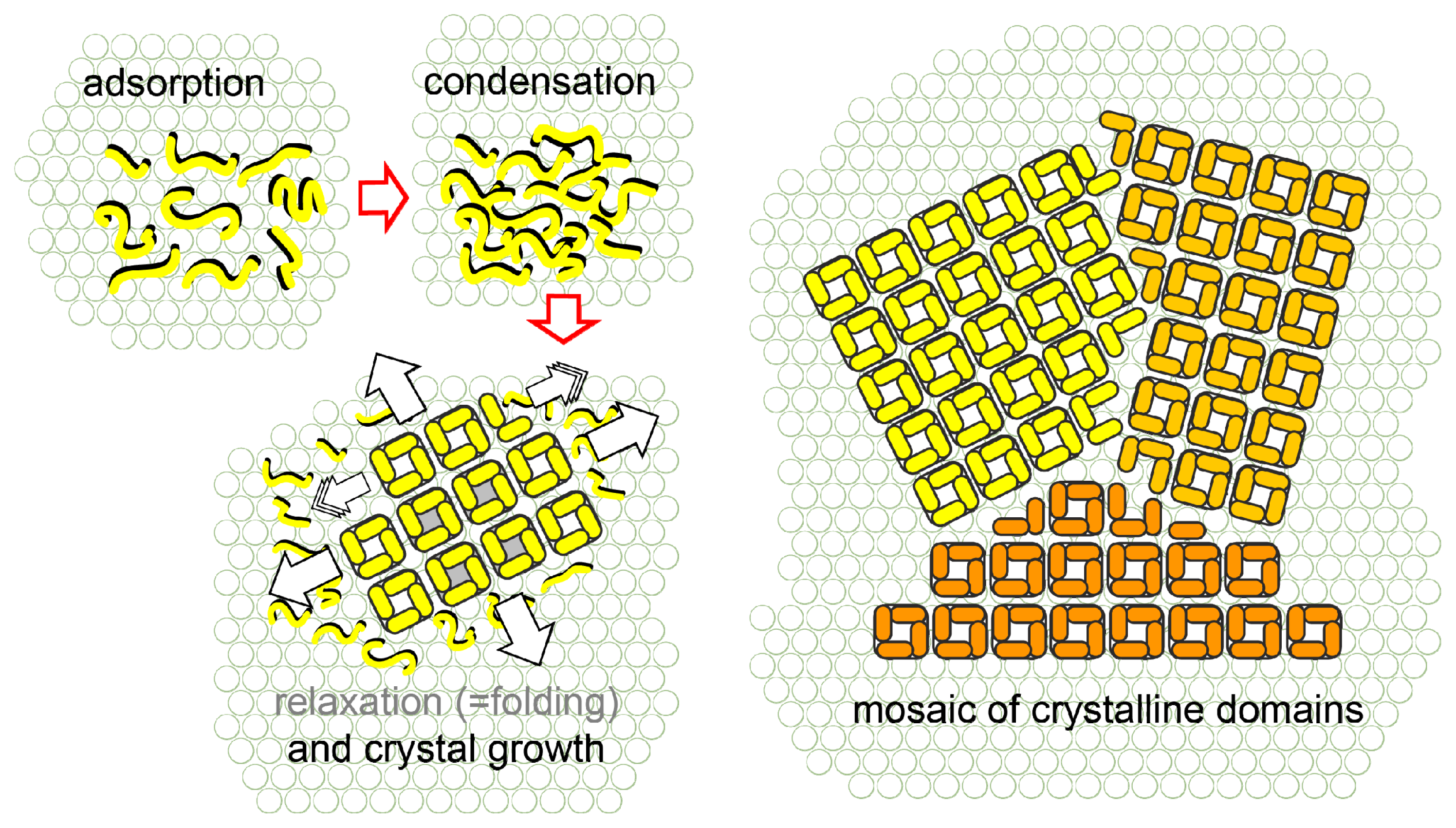
- Shin, S.-H.; Chung, S.; Sanii, B.; Comolli, L.R.; Bertozzi, C.R.; de Yoreo, J.J. Direct observation of kinetic traps associated with structural transformations leading to multiple pathways of S-layer assembly. Proc. Natl. Acad. Sci 2012, 109, 12968–12973. [Google Scholar]
- Pleschberger, M.; Neubauer, A.; Egelseer, E.M.; Weigert, S.; Lindner, B.; Sleytr, U.B.; Muyldermans, S.; Sára, M. Generation of a functional monomolecular protein lattice consisting of an S-layer fusion protein comprising the variable domain of a camel heavy chain antibody. Bioconjugate Chem 2003, 14, 440–448. [Google Scholar]
- Pleschberger, M.; Saerens, D.; Weigert, S.; Sleytr, U.B.; Muyldermans, S.; Sára, M.; Egelseer, E.M. An S-layer heavy chain camel antibody fusion protein for generation of a nanopatterned sensing layer to detect the prostate-specific antigen by surface plasmon resonance technology. Bioconjugate Chem 2004, 15, 664–671. [Google Scholar]
- Völlenkle, C.; Weigert, S.; Ilk, N.; Egelseer, E.; Weber, V.; Loth, F.; Falkenhagen, D.; Sleytr, U.B.; Sára, M. Construction of a functional S-layer fusion protein comprising an immunoglobulin G-binding domain for development of specific adsorbents for extracorporeal blood purification. Appl. Environ. Microbiol 2004, 70, 1514–1521. [Google Scholar]
- Lopez, A.E.; Moreno-Flores, S.; Pum, D.; Sleytr, U.B.; Toca-Herrera, J.L. Surface dependence of protein nanocrystal formation. Small 2010, 6, 396–403. [Google Scholar]
- López, A.E.; Pum, D.; Sleytr, U.B.; Toca-Herrera, J.L. Influence of surface chemistry and protein concentration on the adsorption rate and S-layer crystal formation. Phys. Chem. Chem. Phys 2011, 13, 11905–11913. [Google Scholar]
- Toca-Herrera, J.L.; Krastev, R.; Bosio, V.; Küpcü, S.; Pum, D.; Fery, A.; Sára, M.; Sleytr, U.B. Recrystallization of bacterial S-layers on flat polyelectrolyte surfaces and hollow polyelectrolyte capsules. Small 2005, 1, 339–348. [Google Scholar]
- Delcea, M.; Krastev, R.; Gutlebert, T.; Pum, D.; Sleytr, U.B.; Toca-Herrera, J.L. Mapping bacterial surface layers affinity to polyelectrolytes through the building of hybrid macromolecular structures. J. Nanosci. Nanotechnol 2007, 7, 4260–4266. [Google Scholar]
- Delcea, M.; Krastev, R.; Gutberlet, T.; Pum, D.; Sleytr, U.B.; Toca-Herrera, J.L. Thermal stability, mechanical properties and water content of bacterial protein layers recrystallized on polyelectrolyte multilayers. Soft Matter 2008, 4, 1414–1421. [Google Scholar]
- Schuster, B.; Sleytr, U.B. S-layer-supported lipid membranes. Rev. Mol. Biotechnol 2000, 74, 233–254. [Google Scholar]
- Schuster, B.; Sleytr, U.B. Composite S-layer lipid structures. J. Struct. Biol 2009, 168, 207–216. [Google Scholar]
- Györvary, E.; Wetzer, B.; Sleytr, U.B.; Sinner, A.; Offenhäuser, A.; Knoll, W. Lateral diffusion of lipids in silane-, dextrane- and S-layer protein-supported mono- and bilayers. Langmuir 1999, 15, 1337–1347. [Google Scholar]
- Diederich, A.; Sponer, C.; Pum, D.; Sleytr, U.B.; Lösche, M. Reciprocal influence between the protein and lipid components of a lipid-protein membrane model. Colloids Surf. B 1996, 6, 335–346. [Google Scholar]
- Weygand, M.; Wetzer, B.; Pum, D.; Sleytr, U.B.; Cuvillier, N.; Kjaer, K.; Howes, P.B.; Losche, M. Bacterial S-layer protein coupling to lipids: X-ray reflectivity and grazing incidence diffraction studies. Biophys J 1999, 76, 458–468. [Google Scholar]
- Weygand, M.; Schalke, M.; Howes, P.B.; Kjaer, K.; Friedmann, J.; Wetzer, B.; Pum, D.; Sleytr, U.B.; Lösche, M. Coupling of protein sheet crystals (S-layers) to phospholipid monolayers. J. Mater. Chem 2000, 10, 141–148. [Google Scholar]
- Weygand, M.; Kjaer, K.; Howes, P.B.; Wetzer, B.; Pum, D.; Sleytr, U.B.; Lösche, M. Structural reorganization of phospholipid headgroups upon recrystallization of an S-layer lattice. J. Phys. Chem 2002, 106, 5793–5799. [Google Scholar]
- Chung, S.; Shin, S.H.; Bertozzi, C.R.; De Yoreo, J.J. Self-catalyzed growth of S layers via an amorphous-to-crystalline transition limited by folding kinetics. Proc. Natl. Acad. Sci. USA 2010, 107, 16536–16541. [Google Scholar]
- Schuster, B.; Pum, D.; Braha, O.; Bayley, H.; Sleytr, U.B. Self-assembled α-hemolysin pores in an S-layer-supported lipid bilayer. Biochim. Biophys. Acta 1998, 1370, 280–288. [Google Scholar]
- Schuster, B.; Sleytr, U.B. Single channel recordings of α-hemolysin reconstituted in S-layer-supported lipid bilayers. Bioelectrochemistry 2002, 55, 5–7. [Google Scholar]
- Schuster, B.; Gufler, P.C.; Pum, D.; Sleytr, U.B. Interplay of phospholipase A2 with S-layer-supported lipid monolayers. Langmuir 2003, 19, 3393–3397. [Google Scholar]
- Schuster, B.; Pum, D.; Sleytr, U.B. S-layer stabilized lipid membranes. Biointerphases 2008, 3, FA3–11. [Google Scholar]
- Küpcü, S.; Sára, M.; Sleytr, U.B. Liposomes coated with crystalline bacterial cell surface protein (S-layers) as immobilization structures for macromolecules. Biochim. Biophys. Acta 1995, 1235, 263–269. [Google Scholar]
- Mader, C.; Küpcü, S.; Sára, M.; Sleytr, U.B. Stabilizing effect of an S-layer on liposomes towards thermal or mechanical stress. Biochim. Biophys. Acta 1999, 1418, 106–116. [Google Scholar]
- Mader, C.; Küpcü, S.; Sleytr, U.B.; Sára, M. S-layer-coated liposomes as a versatile system for entrapping and binding target molecules. Biochim. Biophys. Acta 2000, 1463, 142–150. [Google Scholar]
- Martín-Molina, A.; Moreno-Flores, S.; Perez, E.; Pum, D.; Sleytr, U.B.; Toca-Herrera, J.L. Structure, surface interactions, and compressibility of bacterial S-layers through scanning force microscopy and the surface force apparatus. Biophys. J 2006, 90, 1821–1829. [Google Scholar]
- Pavkov, T.; Egelseer, E.M.; Tesarz, M.; Svergun, D.I.; Sleytr, U.B.; Keller, W. The structure and binding behavior of the bacterial cell surface layer protein Sbsc. Structure 2008, 16, 1226–1237. [Google Scholar]
- Fagan, R.P.; Albesa-Jové, D.; Qazi, O.; Svergun, D.I.; Brown, K.A.; Fairweather, N.F. Structural insights into the molecular organization of the S-layer from Clostridium difficile. Mol. Microbiol 2009, 71, 1308–1322. [Google Scholar]
- Horejs, C.; Pum, D.; Sleytr, U.B.; Tscheliessnig, R. Structure prediction of an S-layer protein by the mean force method. J. Chem. Phys 2008, 128, 65106. [Google Scholar]
- Horejs, C.; Gollner, H.; Pum, H.; Sleytr, U.B.; Peterlik, H.; Jungbauer, A.; Tscheliessnig, R. Atomistic structure of monomolecular surface layer self-assemblies: Toward functionalized nanostructures. ACS Nano 2011, 5, 2288–2297. [Google Scholar]
- Horejs, C.; Pum, D.; Sleytr, U.B.; Peterlik, H.; Jungbauer, A.; Tscheliessnig, R. Surface layer protein characterization by small angle X-ray scattering and a fractal mean force concept: From protein structure to nanodisk assemblies. J. Chem. Phys 2010, 133, 175102. [Google Scholar]
- Horejs, C.; Mitra, M.K.; Pum, D.; Sleytr, U.B.; Muthukumar, M. Monte carlo study of the molecular mechanisms of surface-layer protein self-assembly. J. Chem. Phys 2011, 134, 125103. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Functionality | Application |
|---|---|
| Core streptavidin | Binding biotinylated ligands (DNA, protein), Biochip development |
| Major birch pollen allergen (Bet v1) | Vaccines, treatment of type 1 allergy |
| Strep-tag II, Affinity tag for streptavidin | Biochip development |
| ZZ, IgG-binding domain of Protein A | Extracorporeal blood purification |
| Enhanced green fluorescent protein (EGFP) | Liposomes, Drug and delivery systems |
| cAb, Heavy chain camel antibody | Diagnostic systems and sensing layer for labelfree detection systems |
| Hyperthermophilic enzyme laminarinase (LamA) | Immobilized biocatalysts |
| Cysteine mutants | Building of nanoparticle arrays |
| Mimotope of an Epstein-Barr virus (EBV) epitope (F1) | Vaccine development |
| M. tuberculosis antigen (mpt64) | Vaccine development |
| IgG-Binding domain of Protein G | Downstream processing |
| Glucose-1-phosphate thymidylyltransferase (RmlA) | Immobilized biocatalysts |
| Enhanced cyan (ECFP), green (EGFP), yellow (YFP), monomeric red (RFP1) fluorescent protein | pH biosensors in vivo or in vitro, fluor. markers for drug delivery systems |
| Metal, silica and titania precipitating peptides | Material sciences |
© 2013 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Pum, D.; Toca-Herrera, J.L.; Sleytr, U.B. S-Layer Protein Self-Assembly. Int. J. Mol. Sci. 2013, 14, 2484-2501. https://doi.org/10.3390/ijms14022484
Pum D, Toca-Herrera JL, Sleytr UB. S-Layer Protein Self-Assembly. International Journal of Molecular Sciences. 2013; 14(2):2484-2501. https://doi.org/10.3390/ijms14022484
Chicago/Turabian StylePum, Dietmar, Jose Luis Toca-Herrera, and Uwe B. Sleytr. 2013. "S-Layer Protein Self-Assembly" International Journal of Molecular Sciences 14, no. 2: 2484-2501. https://doi.org/10.3390/ijms14022484
APA StylePum, D., Toca-Herrera, J. L., & Sleytr, U. B. (2013). S-Layer Protein Self-Assembly. International Journal of Molecular Sciences, 14(2), 2484-2501. https://doi.org/10.3390/ijms14022484