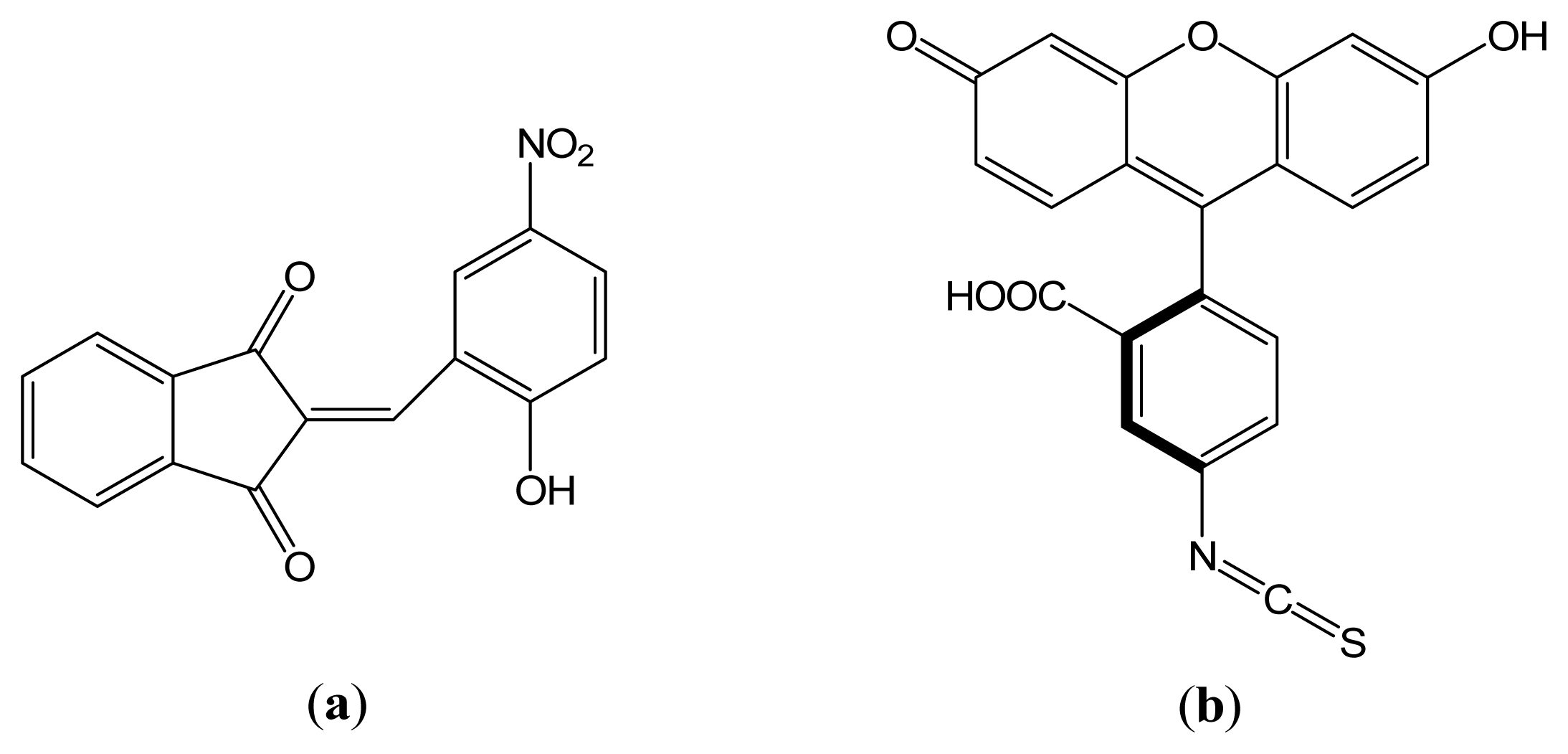
2-(2-Hydroxy-5-nitrobenzylidene)-1,3-indanedione versus Fluorescein Isothiocyanate in Interaction with Anti-hFABP Immunoglobulin G1: Fluorescence Quenching, Secondary Structure Alteration and Binding Sites Localization

Abstract
:1. Introduction
2. Results and Discussion
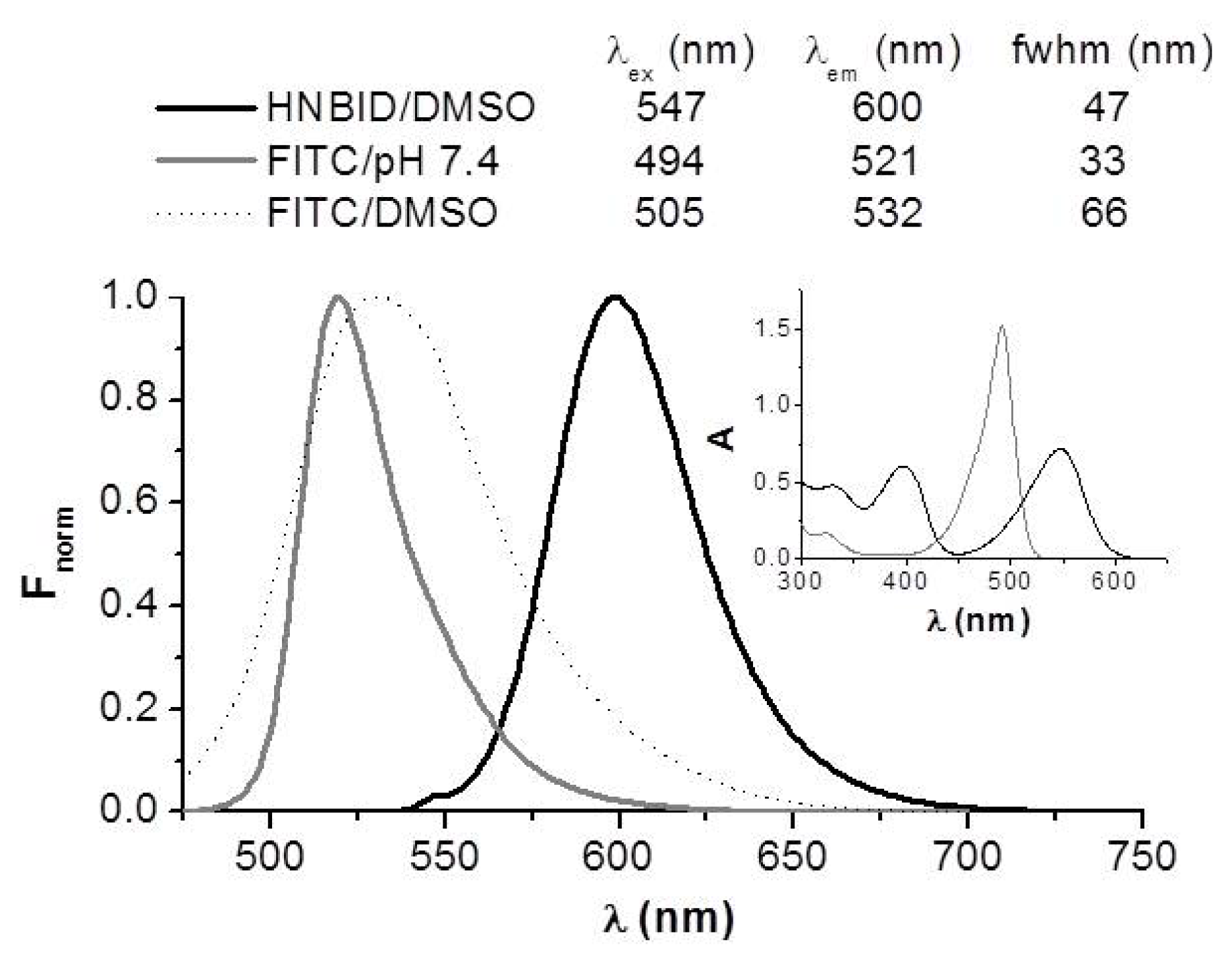
2.1. Effects of HNBID and FITC upon the Fluorescence Emission of Anti-hFABP
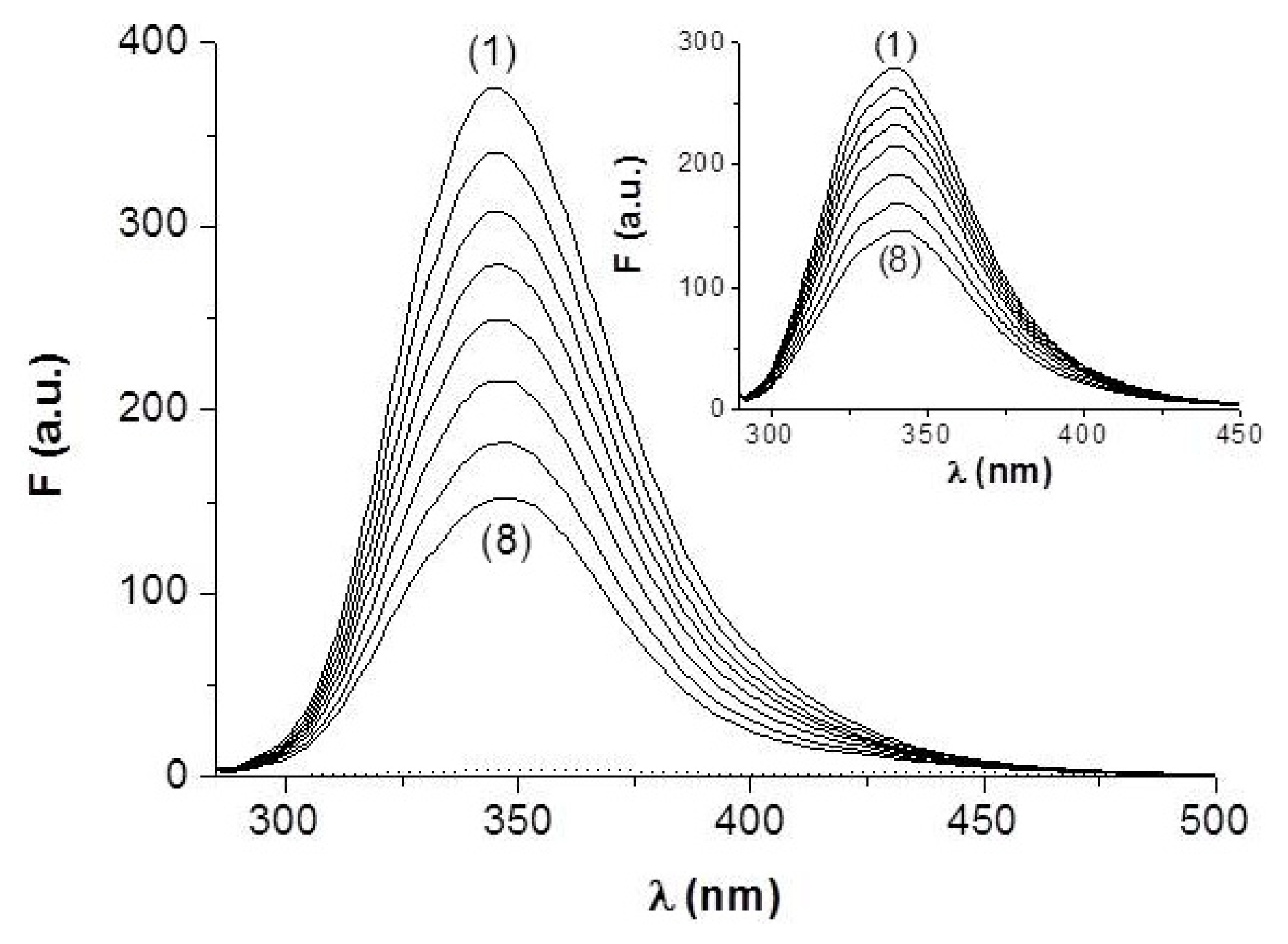
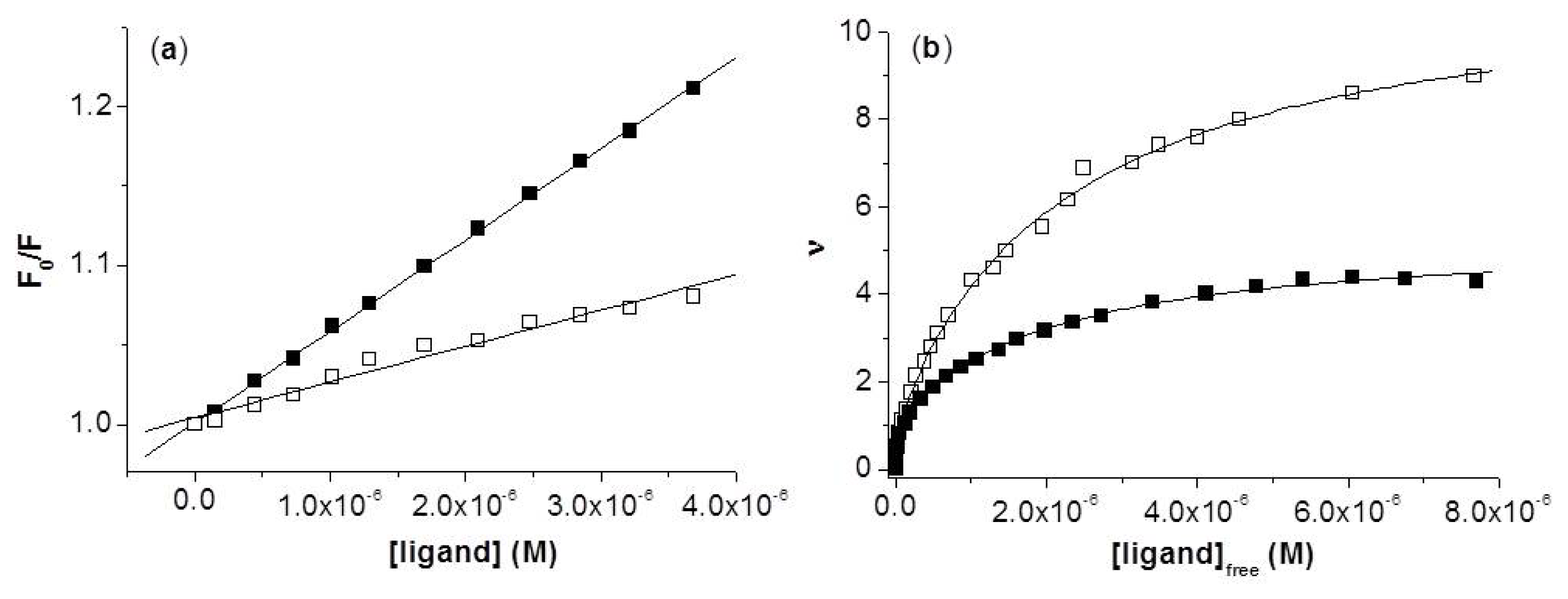
2.1.1. Steady-State Fluorescence
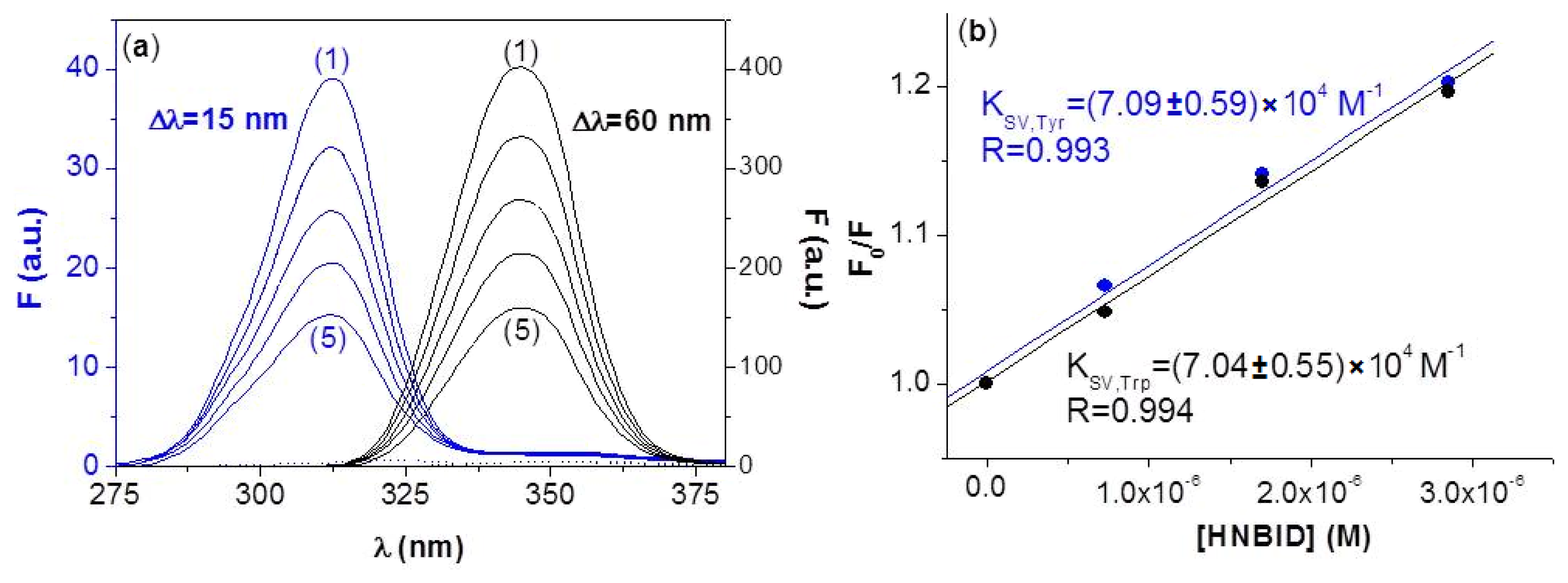
2.1.2. Synchronous Fluorescence
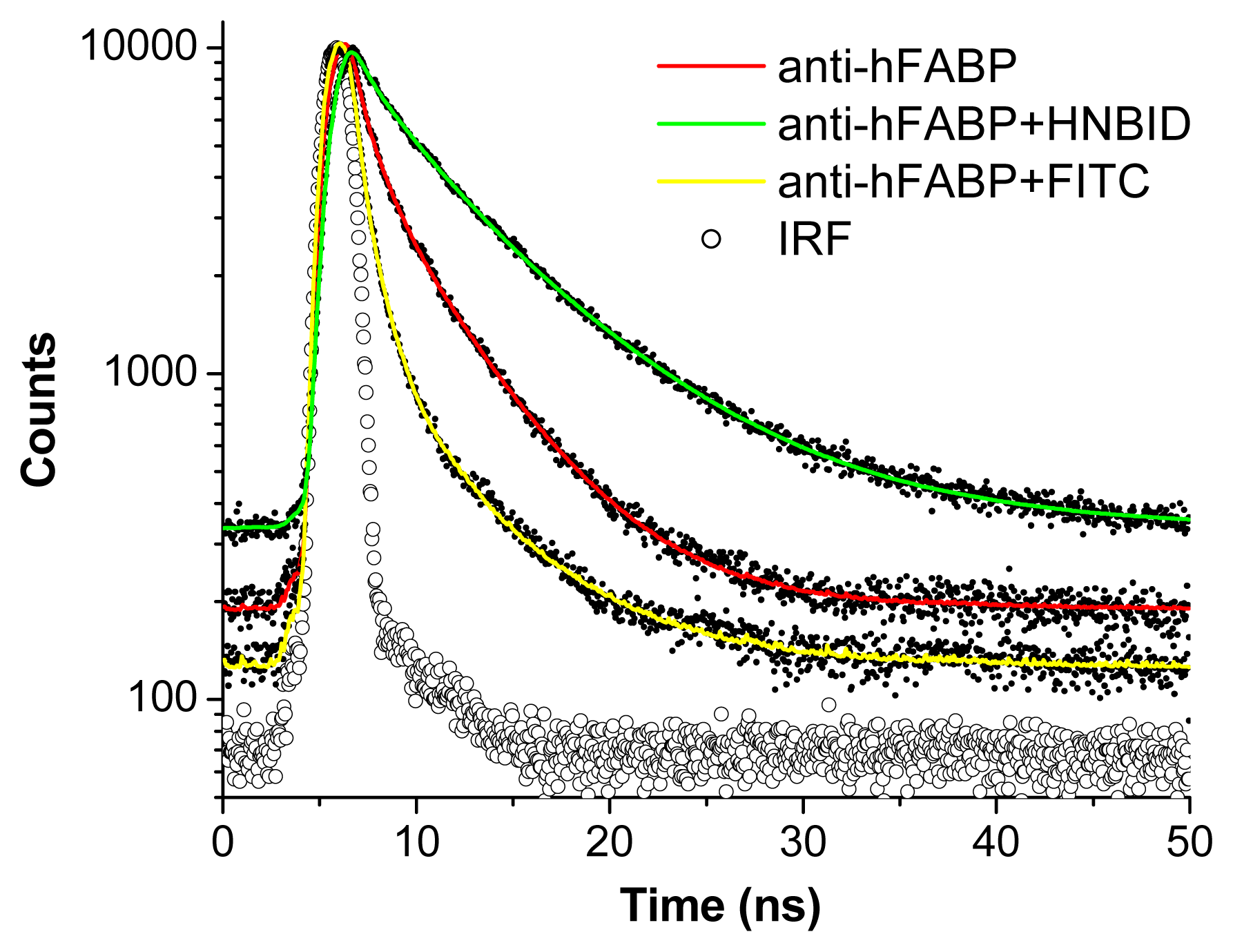
2.1.3. Time-Resolved Fluorescence
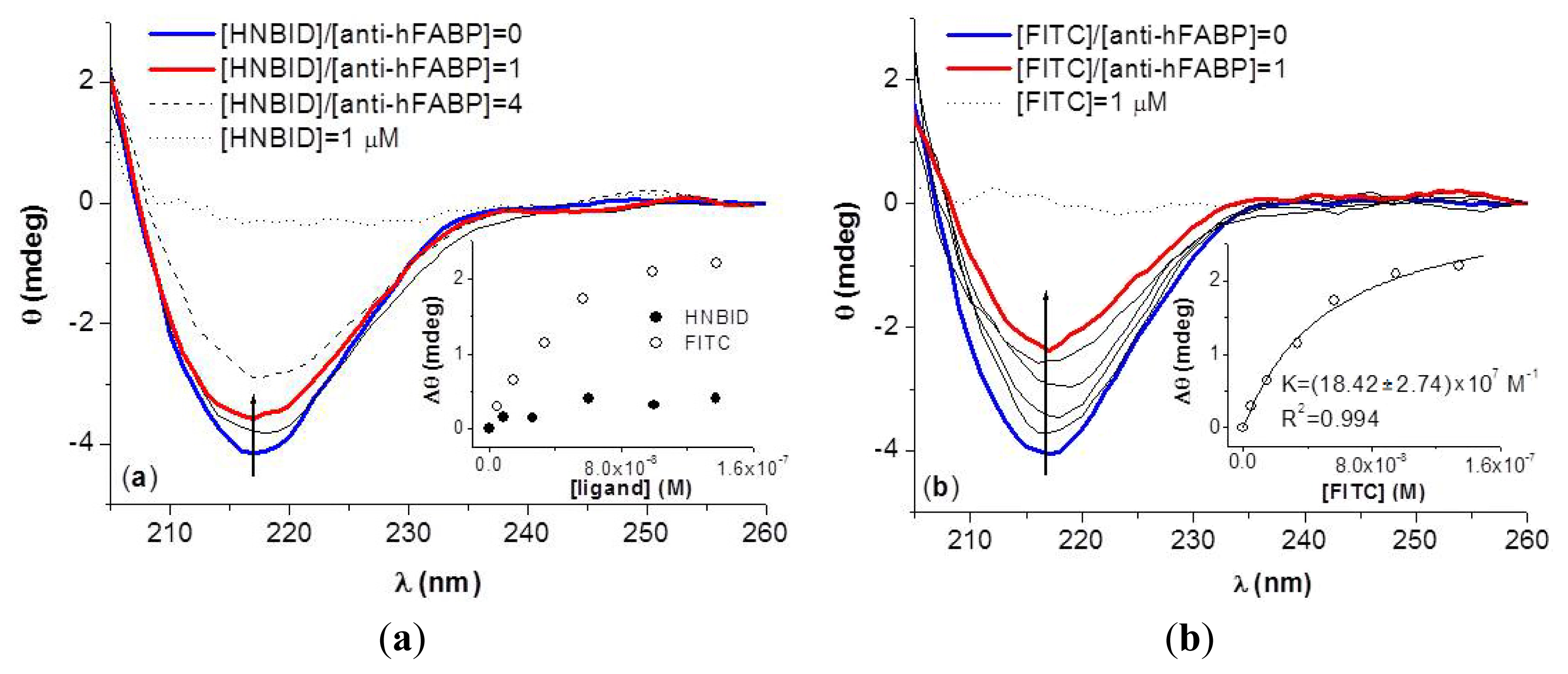
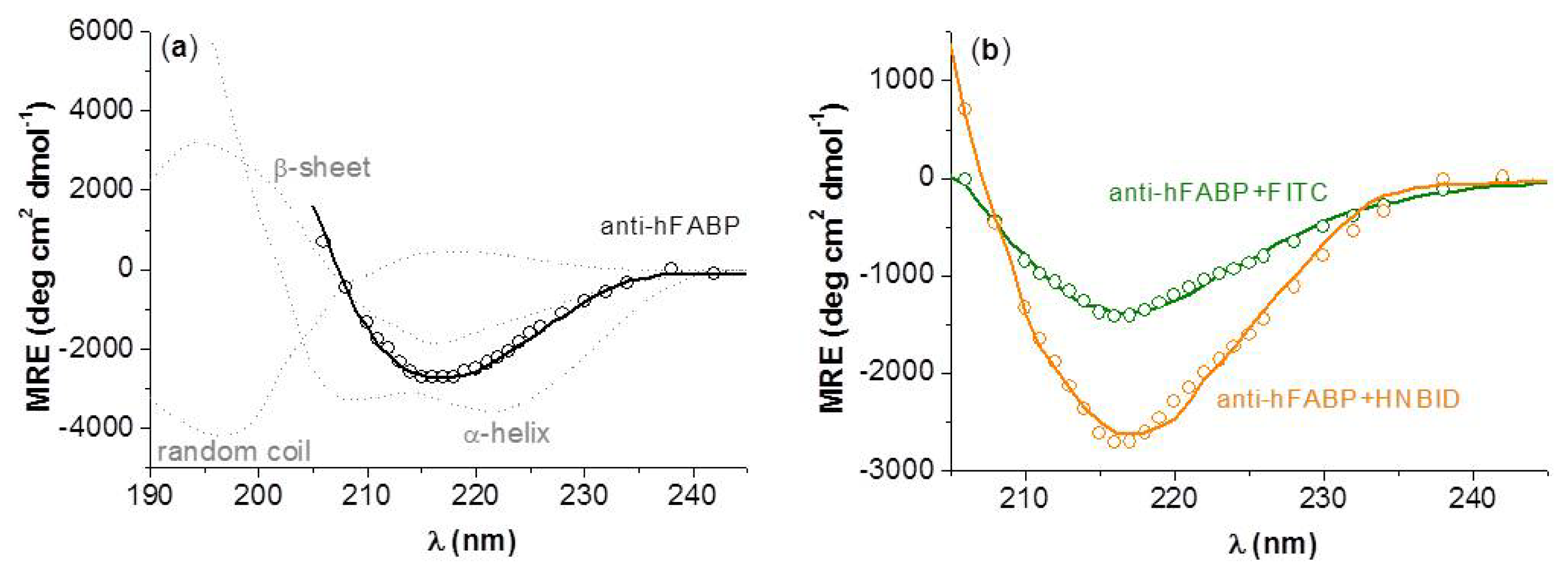
2.2. Effects of HNBID and FITC on the Anti-hFABP Secondary Structure
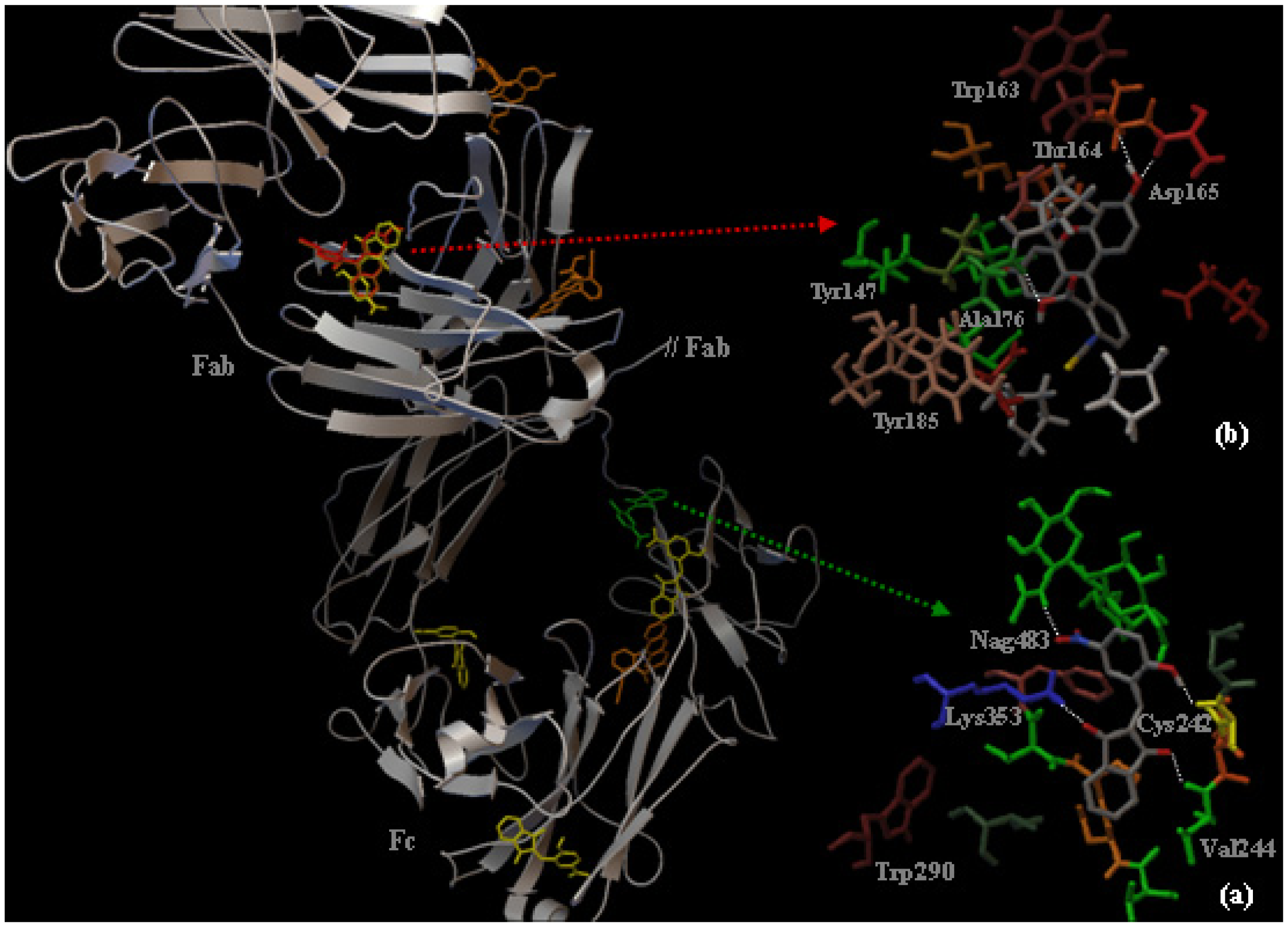
2.3. Localization of the HNBID and FITC Binding Sites on IgG1 Anti-hFABP
3. Experimental Section
4. Conclusions
Acknowledgments
Conflict of Interest
References
- Chen, L.; Guo, X.; Yang, F. Role of heart-type fatty acid binding protein in early detection of acute myocardial infarction in comparison with cTnI, CK-MB and myoglobin. J. Huazhong Univ. Sci. Technol. Med. Sci 2004, 24, 449–451. [Google Scholar]
- O’Donoghue, M.; de Lemos, J.A.; Morrow, D.A.; Murphy, S.A.; Buros, J.L.; Cannon, C.P.; Sabatine, M.S. Prognostic utility of heart-type fatty acid binding protein in patients with acute coronary syndromes. Circulation 2006, 114, 550–557. [Google Scholar]
- Kilcullen, N.; Viswanathan, K.; Das, R.; Morell, C.; Farrin, A.; Barth, J.H.; Hall, A.S. Heart-type fatty acid-binding protein predicts long-term mortality after acute coronary syndrome and identifies high-risk patients across the range of troponin values. J. Am. Coll. Cardiol 2007, 50, 2061–2067. [Google Scholar]
- Chan, D.; Ng, L.L. Biomarkers in acute myocardial infarction. BMC Med. 2010, 8. [Google Scholar] [CrossRef]
- Jobbágy, A.; Király, K. Chemical characterization of fluorescein isothiocyanate-protein conjugates. Biochim. Biophys. Acta 1966, 124, 166–175. [Google Scholar]
- Hermanson, G.T. Bioconjugate Techniques, 2nd ed; Academic Press: San Diego, CA, USA, 2008; pp. 783–787. [Google Scholar]
- Dayan, S.; Almog, J.; Khodzhaev, O.; Rozen, S. A novel synthesis of indanediones using the HOF.CH3CN complex. J. Org. Chem 1998, 63, 2752–2754. [Google Scholar]
- Ramotowski, R.; Cantu, A.A.; Joullié, M.M.; Petrovskaia, O. 1,2-Indanediones: A preliminary evaluation of a new class of amino acid visualizing compounds. Fingerpr. World 1997, 23, 131–140. [Google Scholar]
- Almog, J.; Levinton-Shamuilov, G.; Cohen, Y.; Azoury, M. Fingerprint reagents with dual action: Color and fluorescence. J. Forensic Sci 2007, 52, 330–334. [Google Scholar]
- Hall, I.; Wong, O.; Chi, L.; Chen, S. Cytotoxicity and mode of action of substituted indan-1,3-diones in murine and human tissue cultured cells. Anticancer Res 1994, 14, 2053–2058. [Google Scholar]
- Le Baut, G.; Loire, S.; Sparfel, L.; Creuzet, M.; Feniou, C.; Pontagnier, H.; Prat, G. N-substituted 2-aminomethylene-1,3-indanediones for Treating Arterial Hypertension and Spasmodic Conditions. U.S. Patent 4,758,561, 19 July 1988. [Google Scholar]
- Zalukaev, L.P.; Kozyreva, O.V. Allosteric effects in the mechanism of action of derivatives of indane-1,3-dione on proteins. Pharm. Chem. J 1979, 13, 1126–1128. [Google Scholar]
- Stan, D.; Matei, I.; Mihailescu, C.; Savin, M.; Hillebrand, M.; Baciu, I. Spectroscopic investigations of the binding interaction of a new indanedione derivative with human and bovine serum albumins. Molecules 2009, 14, 1614–1626. [Google Scholar]
- Liu, Y.C.; Yang, Z.Y.; Du, J.; Yao, X.J.; Lei, R.X.; Zheng, X.D.; Liu, J.N.; Hu, H.S.; Li, H. Study on the interactions of kaempferol and quercetin with intravenous immunoglobulin by fluorescence quenching, Fourier transformation infrared spectroscopy and circular dichroism spectroscopy. Chem. Pharm. Bull 2008, 56, 443–451. [Google Scholar]
- Liu, Y.; Yang, Z.; Du, J.; Yao, X.; Zheng, X.; Lei, R.; Liu, J.; Hu, H.; Li, H. Interaction of Taxol with intravenous immunoglobulin: An inhibition of Taxol from crystallizing in aqueous solution. Int. Immunopharmacol 2008, 8, 390–400. [Google Scholar]
- Santos, N.C.; Figueira-Coelho, J.; Martins-Silva, J.; Saldanha, C. Multidisciplinary utilization of dimethyl sulfoxide: Pharmacological, cellular, and molecular aspects. Biochem. Pharmacol 2003, 65, 1035–1041. [Google Scholar]
- Wiberg, K.; Sterner-Molin, A.; Jacobsson, S.P. Simultaneous determination of albumin and immunoglobulin G with fluorescence spectroscopy and multivariate calibration. Talanta 2004, 62, 567–574. [Google Scholar]
- Eftink, M.R. Fluorescence quenching. Theory and applications. In Principles of Fluorescence Spectroscopy; Lakowicz, J.R., Ed.; Kluwer Academic/Plenum Publishers: New York, NY, USA, 1999; pp. 53–127. [Google Scholar]
- Scatchard, G. The attractions of proteins for small molecules and ions. Ann. N.Y. Acad. Sci 1949, 51, 660–672. [Google Scholar]
- Höfliger, M.M.; Beck-Sickinger, A.G. Receptor–ligand interaction. In Protein-Ligand Interactions from Molecular Recognition to Drug Design; Böhm, H.J., Schneider, G., Eds.; Wiley-VCH Verlag: Weinheim, Germany, 2003; p. 113. [Google Scholar]
- Parsons, M.; Messent, A.J.; Humphries, J.D.; Deakin, N.O.; Humphries, M.J. Quantification of integrin receptor agonism by fluorescence lifetime imaging. J. Cell Sci 2008, 121, 265–271. [Google Scholar]
- Duncan, R.R. Fluorescence lifetime imaging microscopy (FLIM) to quantify protein–protein interactions inside cells. Biochem. Soc. Trans 2006, 34, 679–682. [Google Scholar]
- Van Hoek, A.; Visser, A.J.W.G. Time-resolved fluorescence techniques applied to biological interfaces. Appl. Spectrosc. Rev 2000, 35, 255–276. [Google Scholar]
- Klimtchuk, E.; Venyaminov, S.; Kurian, E.; Wessels, W.; Kirk, W.; Prendergast, F.G. Photophysics of ANS. I. Protein–ANS complexes: Intestinal fatty acid binding protein and single-trp mutants. Biophys. Chem 2007, 125, 1–12. [Google Scholar]
- Amzel, L.M.; Poljak, R.J. Three-dimensional structure of immunoglobulins. Annu. Rev. Biochem 1979, 48, 961–997. [Google Scholar]
- Vermeer, A.W.P.; Norde, W. The thermal stability of immunoglobulin: Unfolding and aggregation of a multi-domain protein. Biophys. J 2000, 78, 394–404. [Google Scholar]
- Matei, I.; Soare, L.; Tablet, C.; Hillebrand, M. Characterization of simvastatin and its cyclodextrin inclusion complexes by absorption and circular dichroism spectroscopies and molecular mechanics calculations. Rev. Roum. Chim 2009, 54, 133–141. [Google Scholar]
- Mian, I.S.; Bradwell, A.R.; Olson, A.J. Structure, function and properties of antibody binding sites. J. Mol. Biol 1991, 217, 133–151. [Google Scholar]
- Yang, H.; Gurgel, P.V.; Williams, D.K., Jr; Bobay, B.G.; Cavanagh, J.; Muddiman, D.C.; Carbonell, R.G. Binding site on human immunoglobulin G for the affinity ligand HWRGWV. J. Mol. Recognit. 2010, 23, 271–282. [Google Scholar]
- Silverton, E.W.; Navia, M.A.; Davies, D.R. Three-dimensional structure of an intact human immunoglobulin. Proc. Natl. Acad. Sci. USA 1977, 74, 5140–5144. [Google Scholar]
- Ionescu, M.V. New reaction for aromatic aldehydes. Bull. Soc. Chim 1930, 47, 210–214. [Google Scholar]
- Epps, D.E.; Raub, T.J.; Caiolfa, V.; Chiari, A.; Zamai, M. Determination of the affinity of drugs toward serum albumin by measurement of the quenching of the intrinsic tryptophan fluorescence of the protein. J. Pharm. Pharmacol 1999, 51, 41–48. [Google Scholar]
- Otosu, T.; Nishimoto, E.; Yamashita, S. Multiple conformational state of human serum albumin around single tryptophan residue at various pH revealed by time-resolved fluorescence spectroscopy. J. Biochem 2010, 147, 191–200. [Google Scholar]
- Greenfield, N.J.; Fasman, G.D. Computed circular dichroism spectra for the evaluation of protein conformation. Biochemistry 1969, 8, 4108–4116. [Google Scholar]
- Kelly, S.M.; Jess, T.J.; Price, N.C. How to study proteins by circular dichroism. Biochim. Biophys. Acta 2005, 1751, 119–139. [Google Scholar]
- Schmidt, M.W.; Baldridge, K.K.; Boatz, J.A.; Elbert, S.T.; Gordon, M.S.; Jensen, J.H.; Koseki, S.; Matsunaga, N.; Nguyen, K.A.; Su, S.; Windus, T.L.; Dupuis, M.; Montgomery, J.A. General atomic and molecular electronic structure system. J. Comput. Chem 1993, 14, 1347–1363. [Google Scholar]
- Tomasi, J.; Mennucci, B.; Cammi, R. Quantum mechanical continuum solvation models. Chem. Rev 2005, 105, 2999–3093. [Google Scholar]
- Morris, G.M.; Goodsell, D.S.; Halliday, R.S.; Huey, R.; Hart, W.E.; Belew, R.K.; Olson, A.J. Automated docking using a Lamarckian genetic algorithm and empirical binding free energy function. J. Comput. Chem 1998, 19, 1639–1662. [Google Scholar]
- Sanner, M.F. Python: A programming language for software integration and development. J. Mol. Graphics Model 1999, 17, 57–61. [Google Scholar]
- RCSB Protein Data Bank. Available online: http://www.rcsb.org/pdb accessed on 1 October 2012.
- Harris, L.J.; Skaletsky, E.; McPherson, A. Crystallographic structure of an intact IgG1 monoclonal antibody1. J. Mol. Biol 1998, 275, 861–872. [Google Scholar]
- Goodsell, D.S.; Morris, G.M.; Olson, A.J. Automated docking of flexible ligands: Applications of AutoDock. J. Mol. Recognit 1996, 9, 1–5. [Google Scholar]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ligand | KSV × 10−4 (M−1) | Ra; SD b; Nc | K × 10−6 (M−1) | n | R; SD; N |
|---|---|---|---|---|---|
| HNBID | 5.75 ± 0.04 | 0.999; 0.002; 12 | 41.35 ± 16.85 5.22 ± 0.60 | 1.00 4.36 | 0.997; 0.080; 23 |
| FITC | 2.25 ± 0.12 | 0.986; 0.005; 12 | 50.41 ± 30.59 0.44 ± 0.04 | 1.01 10.41 | 0.998; 0.142; 23 |
| System | τ1 (ns) | B1 | τ2 (ns) | B2 | τ3 (ns) | B3 | <τ> (ns) | χ2a |
|---|---|---|---|---|---|---|---|---|
| anti-hFABP | <0.10 * | – * | 1.15 | 0.02 | 4.47 | 0.01 | 2.91 | 1.684 |
| HNBID–anti-hFABP | 0.37 | 0.02 | 2.69 | 0.01 | 7.33 | 0.02 | 6.14 | 1.373 |
| FITC–anti-hFABP | <0.10 * | – * | 1.00 | 0.03 | 5.43 | 0.01 | 1.74 | 1.663 |
| System | % β-sheet | % α-helix | % random coil | R2 |
|---|---|---|---|---|
| anti-hFABP | 88.67 | 2.09 | 9.24 | 0.987 |
| HNBID–anti-hFABP | 86.54 | 2.10 | 11.36 | 0.985 |
| FITC–anti-hFABP | 82.58 | 1.09 | 16.33 | 0.991 |
Supplementary Files
© 2013 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Stan, D.; Mihailescu, C.-M.; Savin, M.; Matei, I. 2-(2-Hydroxy-5-nitrobenzylidene)-1,3-indanedione versus Fluorescein Isothiocyanate in Interaction with Anti-hFABP Immunoglobulin G1: Fluorescence Quenching, Secondary Structure Alteration and Binding Sites Localization. Int. J. Mol. Sci. 2013, 14, 3011-3025. https://doi.org/10.3390/ijms14023011
Stan D, Mihailescu C-M, Savin M, Matei I. 2-(2-Hydroxy-5-nitrobenzylidene)-1,3-indanedione versus Fluorescein Isothiocyanate in Interaction with Anti-hFABP Immunoglobulin G1: Fluorescence Quenching, Secondary Structure Alteration and Binding Sites Localization. International Journal of Molecular Sciences. 2013; 14(2):3011-3025. https://doi.org/10.3390/ijms14023011
Chicago/Turabian StyleStan, Dana, Carmen-Marinela Mihailescu, Mihaela Savin, and Iulia Matei. 2013. "2-(2-Hydroxy-5-nitrobenzylidene)-1,3-indanedione versus Fluorescein Isothiocyanate in Interaction with Anti-hFABP Immunoglobulin G1: Fluorescence Quenching, Secondary Structure Alteration and Binding Sites Localization" International Journal of Molecular Sciences 14, no. 2: 3011-3025. https://doi.org/10.3390/ijms14023011
APA StyleStan, D., Mihailescu, C. -M., Savin, M., & Matei, I. (2013). 2-(2-Hydroxy-5-nitrobenzylidene)-1,3-indanedione versus Fluorescein Isothiocyanate in Interaction with Anti-hFABP Immunoglobulin G1: Fluorescence Quenching, Secondary Structure Alteration and Binding Sites Localization. International Journal of Molecular Sciences, 14(2), 3011-3025. https://doi.org/10.3390/ijms14023011