Role of Homeodomain Leucine Zipper (HD-Zip) IV Transcription Factors in Plant Development and Plant Protection from Deleterious Environmental Factors

Abstract
:1. Background
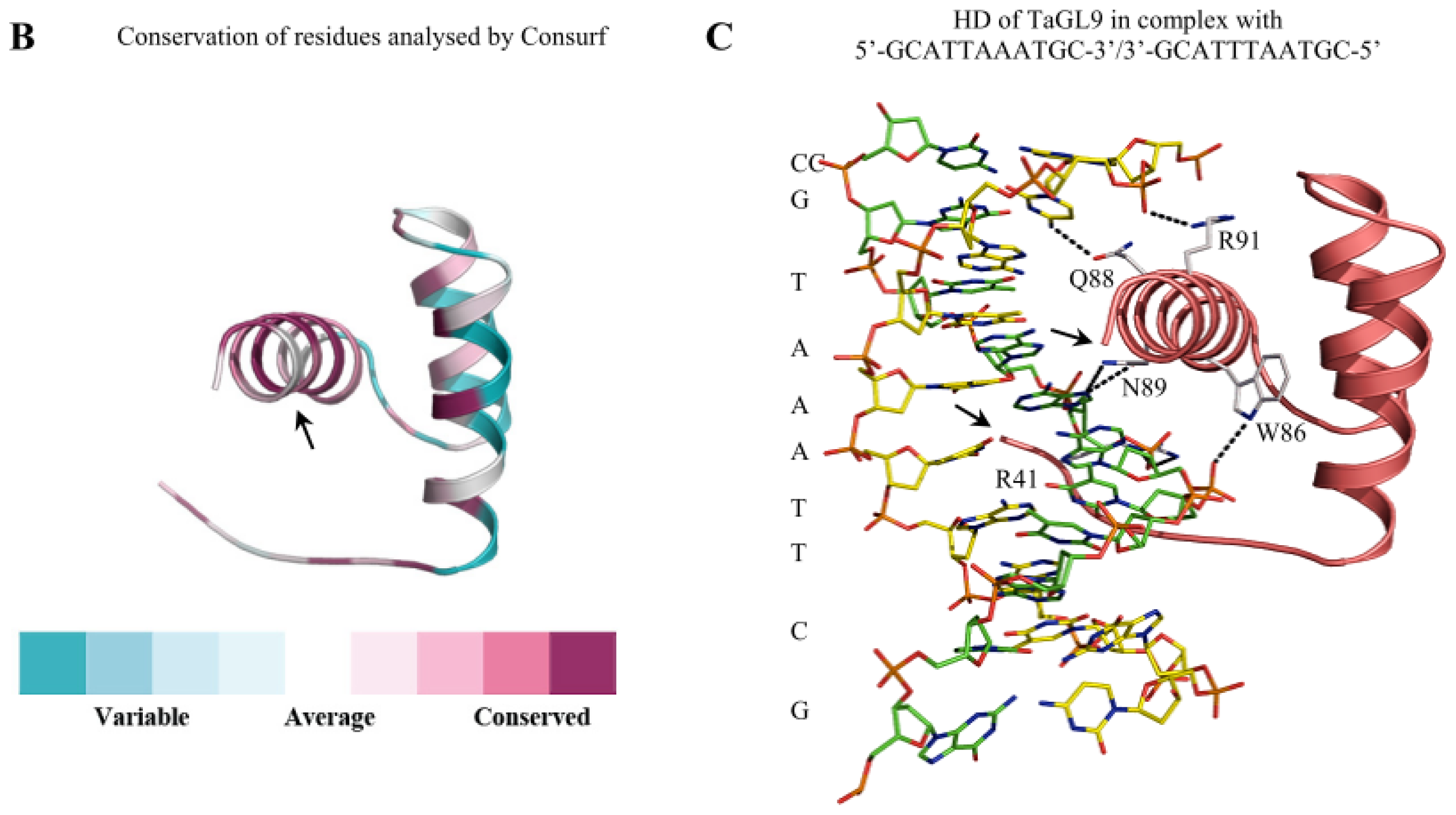
2. Structure of HD-Zip IV TFs and Function of Identified Domains
3. Regulation of HD-Zip IV Gene Expression
4. Roles of HD-Zip IV TFs
4.1. Development and Maintenance of Epidermal Cell Layers
4.2. Development of Sub-Epidermal Cell Layers and Control of Anthocyanin Pigmentation
4.3. Drought Tolerance
4.4. Function of HD-Zip IV Genes in Commercially Important Plant Species
4.5. Regulation of Cuticle Formation and Plant Protection
5. Conclusions and Future Perspectives
Acknowledgments
Conflict of Interest
References
- Wang, R.L.; Stec, A.; Hey, J.; Lukens, L.; Doebley, J. The limits of selection during maize domestication. Nature 1999, 398, 236–239. [Google Scholar]
- Bustamante, C.D.; Fledel-Alon, A.; Williamson, S.; Nielsen, R.; Hubisz, M.T.; Glanowski, S.; Tanenbaum, D.M.; White, T.J.; Sninsky, J.J.; Hernandez, R.D.; et al. Natural selection on protein-coding genes in the human genome. Nature 2005, 437, 1153–1157. [Google Scholar]
- Ma, J.; Ptashne, M. A new class of yeast transcriptional activators. Cell 1987, 51, 113–119. [Google Scholar]
- Garber, R.L.; Kuroiwa, A.; Gehring, W.J. Genomic and cDNA clones of the homeotic locus antennapedia in Drosophila. EMBO J 1983, 2, 2027–2036. [Google Scholar]
- Gehring, W.J.; Affolter, M.; Burglin, T. Homeodomain proteins. Annu. Rev. Biochem 1994, 63, 487–526. [Google Scholar]
- Chan, R.L.; Gago, G.M.; Palena, C.M.; Gonzalez, D.H. Homeoboxes in plant development. Biochim. Biophys. Acta 1998, 23, 1–19. [Google Scholar]
- Khaled, A.S.; Vernoud, V.; Ingram, G.C.; Perez, P.; Sarda, X.; Rogowsky, P.M. Engrailed-ZmOCL1 fusions cause a transient reduction of kernel size in maize. Plant Mol. Biol 2005, 58, 123–139. [Google Scholar]
- Vollbrecht, E.; Veit, B.; Sinha, N.; Hake, S. The developmental gene Knotted-1 is a member of a maize homeobox gene family. Nature 1991, 350, 241–243. [Google Scholar]
- Ariel, F.D.; Manavella, P.A.; Dezar, C.A.; Chan, R.L. The true story of the HD-Zip family. Trends Plant Sci 2007, 12, 419–426. [Google Scholar]
- Mukherjee, K.; Brocchieri, L.; Bürglin, T.R. A comprehensive classification and evolutionary analysis of plant homeobox genes. Mol. Biol. Evol 2009, 26, 2775–2794. [Google Scholar]
- Ruberti, I.; Sessa, G.; Lucchetti, S.; Morelli, G. A novel class of plant proteins containing a homeodomain with a closely linked leucine zipper motif. EMBO J 1991, 10, 1787–1791. [Google Scholar]
- Riechmann, J.L.; Heard, J.; Martin, G.; Reuber, L.; Jiang, C.; Keddie, J.; Adam, L.; Pineda, O.; Ratcliffe, O.J.; Samaha, R.R.; et al. Arabidopsis transcription factors: Genome-wide comparative analysis among eukaryotes. Science 2000, 290, 2105–2110. [Google Scholar]
- Schrick, K.; Nguyen, D.; Karlowski, W.M.; Mayer, K.F. START lipid/sterol-binding domains are amplified in plants and are predominantly associated with homeodomain transcription factors. Genome Biol. 2004, 5. [Google Scholar] [CrossRef] [Green Version]
- Mukherjee, K.; Bürglin, T.R. MEKHLA, a novel domain with similarity to PAS domains, is fused to plant homeodomain-leucine zipper III proteins. Plant Physiol 2006, 140, 1142–1150. [Google Scholar]
- Sessa, G.; Morelli, G.; Ruberti, I. The Athb-1 and -2 HD-Zip domains homodimerize forming complexes of different DNA binding specificities. EMBO J 1993, 12, 3507–3517. [Google Scholar]
- Harris, J.C.; Hrmova, M.; Lopato, S.; Langridge, P. Modulation of plant growth by HD-Zip class I and II transcription factors in response to environmental stimuli. New Phytol 2011, 190, 823–837. [Google Scholar]
- Yu, H.; Chen, X.; Hong, Y.-Y.; Wang, Y.; Xu, P.; Ke, S.-D.; Liu, H.-Y.; Zhu, J.-K.; Oliver, D.J.; Xiang, C.-B. Activated expression of an Arabidopsis HD-START protein confers drought tolerance with improved root system and reduced stomatal density. Plant Cell 2008, 20, 1134–1151. [Google Scholar]
- Zhang, S.; Haider, I.; Kohlen, W.; Jiang, L.; Bouwmeester, H.; Meijer, A.; Schluepmann, H.; Liu, C.-M.; Ouwerkerk, P.F. Function of the HD-Zip I gene Oshox22 in ABA-mediated drought and salt tolerances in rice. Plant Mol. Biol 2012, 80, 571–585. [Google Scholar]
- Cabello, J.V.; Chan, R.L. The homologous homeodomain-leucine zipper transcription factors HaHB1 and AtHB13 confer tolerance to drought and salinity stresses via the induction of proteins that stabilize membranes. Plant Biotech. J 2012, 10, 815–825. [Google Scholar]
- Nadakuduti, S.S.; Pollard, M.; Kosma, D.K.; Allen, C., Jr; Ohlrogge, J.B.; Barry, CS. Pleiotropic phenotypes of the sticky peel mutant provide new insight into the role of CUTIN DEFICIENT2 in epidermal cell function in tomato. Plant Physiol 2012, 159, 945–960. [Google Scholar]
- Valdés, A.; Övernäs, E.; Johansson, H.; Rada-Iglesias, A.; Engström, P. The homeodomain-leucine zipper (HD-Zip) class I transcription factors ATHB7 and ATHB12 modulate abscisic acid signalling by regulating protein phosphatase 2C and abscisic acid receptor gene activities. Plant Mol. Biol 2012, 80, 405–418. [Google Scholar]
- Lechner, E.; Leonhardt, N.; Eisler, H.; Parmentier, Y.; Alioua, M.; Jacquet, H.; Leung, J.; Genschik, P. MATH/BTB CRL3 receptors target the homeodomain-leucine zipper ATHB6 to modulate abscisic acid signaling. Dev. Cell 2011, 21, 1116–1128. [Google Scholar]
- Javelle, M.; Vernoud, V.; Rogowsky, P.M.; Ingram, G.C. Epidermis: The formation and functions of a fundamental plant tissue. New Phytol 2011, 189, 17–39. [Google Scholar]
- Nakamura, M.; Katsumata, H.; Abe, M.; Yabe, N.; Komeda, Y.; Yamamoto, K.T.; Takahashi, T. Characterization of the class IV homeodomain-leucine zipper gene family in Arabidopsis. Plant Physiol 2006, 141, 1363–7375. [Google Scholar]
- Abe, M.; Takahashi, T.; Komeda, Y. Identification of a cis-regulatory element for L1 layer-specific gene expression, which is targeted by an L1-specific homeodomain protein. Plant J 2001, 26, 487–494. [Google Scholar]
- Ohashi, Y.; Oka, A.; Rodrigues-Pousada, R.; Possenti, M.; Ruberti, I.; Morelli, G.; Aoyama, T. Modulation of phospholipid signaling by GLABRA2 in root-hair pattern formation. Science 2003, 300, 1427–1430. [Google Scholar]
- Graba, Y.; Aragnol, D.; Pradel, J. Drosophila Hox complex downstream targets and the function of homeotic genes. BioEssays 1997, 19, 379–388. [Google Scholar]
- Ekker, S.C.; von Kessler, D.P.; Beachy, P.A. Differential DNA sequence recognition is a determinant of specificity in homeotic gene action. EMBO J 1992, 11, 4059–4072. [Google Scholar]
- Wu, R.; Li, S.; He, S.; Wassmann, F.; Yu, C.; Qin, G.; Schreiber, L.; Qu, L.J.; Gu, H. CFL1, a WW domain protein, regulates cuticle development by modulating the function of HDG1, a class IV homeodomain transcription factor, in rice and Arabidopsis. Plant Cell 2011, 23, 3392–3411. [Google Scholar]
- Kinoshita, T.; Miura, A.; Choi, Y.; Kinoshita, Y.; Cao, X.; Jacobsen, S.E.; Fischer, R.L.; Kakutani, T. One-Way Control of FWA Imprinting in Arabidopsis Endosperm by DNA Methylation. Science 2004, 303, 521–523. [Google Scholar]
- Di Cristina, M.; Sessa, G.; Dolan, L.; Linstead, P.; Baima, S.; Ruberti, I.; Morelli, G. The Arabidopsis Athb-10 (GLABRA2) is an HD-Zip protein required for regulation of root hair development. Plant J 1996, 10, 393–402. [Google Scholar]
- Kubo, H.; Peeters, A.J.; Aarts, M.G.; Pereira, A.; Koornneef, M. ANTHOCYANINLESS2, a homeobox gene affecting anthocyanin distribution and root development in Arabidopsis. Plant Cell 1999, 11, 1217–1226. [Google Scholar]
- Abe, M.; Katsumata, H.; Komeda, Y.; Takahashi, T. Regulation of shoot epidermal cell differentiation by a pair of homeodomain proteins in Arabidopsis. Development 2003, 130, 635–643. [Google Scholar]
- Ingouff, M.; Farbos, I.; Lagercrantz, U.; von Arnold, S. PaHB1 is an evolutionary conserved HD-GL2 homeobox gene expressed in the protoderm during Norway spruce embryo development. Genesis 2001, 30, 220–230. [Google Scholar]
- Ingouff, M.; Farbos, I.; Wiweger, M.; von Arnold, S. The molecular characterization of PaHB2, a homeobox gene of the HD-GL2 family expressed during embryo development in Norway spruce. J. Exp. Bot 2003, 54, 1343–1350. [Google Scholar]
- Guan, X.Y.; Li, Q.J.; Shan, C.M.; Wang, S.; Mao, Y.B.; Wang, L.J.; Chen, X.Y. The HD-Zip IV gene GaHOX1 from cotton is a functional homologue of the Arabidopsis GLABRA2. Plant Physiol 2008, 134, 174–182. [Google Scholar]
- Walford, S.A.; Wu, Y.; Llewellyn, D.J.; Dennis, E.S. Epidermal cell differentiation in cotton mediated by the homeodomain leucine zipper gene, GhHD-1. Plant J 2012, 71, 464–478. [Google Scholar]
- Ingram, G.C.; Magnard, J.L.; Vergne, P.; Dumas, C.; Rogowsky, P.M. Expression patterns of genes encoding HD-Zip IV homeodomain proteins define specific domains in maize embryos and meristems. Plant J 2000, 22, 401–414. [Google Scholar]
- Vernoud, V.; Laigle, G.; Rozier, F.; Meeley, R.B.; Perez, P.; Rogowsky, P.M. The HD-ZIP IV transcription factor OCL4 is necessary for trichome patterning and anther development in maize. Plant J 2009, 59, 883–894. [Google Scholar]
- Javelle, M.; Klein-Cosson, C.; Vernoud, V.; Boltz, V.; Maher, C.; Timmermans, M.; Depege-Fargeix, N.; Rogowsky, P.M. Genome-wide characterization of the HD-ZIP IV transcription factor family in maize: Preferential expression in the epidermis. Plant Physiol 2011, 157, 790–803. [Google Scholar]
- Ito, M.; Sentoku, N.; Nishimura, A.; Hong, S.-K.; Sato, Y.; Matsuoka, M. Position dependent expression of GL2-type homeobox gene, Roc1: Significance for protoderm differentiation and radial pattern formation in early rice embryogenesis. Plant J 2002, 29, 497–507. [Google Scholar]
- Ito, M.; Sentoku, N.; Nishimura, A.; Hong, S.-K.; Sato, Y.; Matsuoka, M. Roles of rice GL-2 type homeobox genes in epidermis differentiation. Breed. Sci 2003, 53, 245–253. [Google Scholar]
- Sasaki, T.; Matsumoto, T.; Yamamoto, K.; Sakata, K.; Baba, T.; Katayose, Y.; Wu, J.; Niimura, Y.; Cheng, Z.; Nagamura, Y.; et al. The genome sequence and structure of rice chromosome 1. Nature 2002, 420, 312–316. [Google Scholar]
- Yu, J.; Wang, J.; Lin, W.; Li, S.; Li, H.; Zhou, J.; Ni, P.; Dong, W.; Hu, S.; Zeng, C.; et al. The genomes of Oryza sativa: A history of duplications. PLoS Biol 2005, 3, e38. [Google Scholar]
- Kovalchuk, N.; Wu, W.; Eini, O.; Bazanova, N.; Pallotta, M.; Shirley, N.; Singh, R.; Ismagul, A.; Eliby, S.; Johnson, A.; et al. The scutellar vascular bundle–specific promoter of the wheat HD-Zip IV transcription factor shows similar spatial and temporal activity in transgenic wheat, barley and rice. Plant Biotech. J 2012, 10, 43–53. [Google Scholar]
- Prigge, M.J.; Clark, S.E. Evolution of the class III HD-Zip gene family in land plants. Evol. Dev 2006, 8, 350–361. [Google Scholar]
- Thompson, J.D.; Gibson, T.J.; Plewniak, F.; Jeanmougin, F.; Higgins, D.G. The CLUSTAL_X windows interface: Flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res 1997, 25, 4876–4882. [Google Scholar]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol 1987, 4, 406–425. [Google Scholar]
- Schultz, J.; Milpetz, F.; Bork, P.; Ponting, C.P. SMART, a simple modular architecture research tool: Identification of signaling domains. Proc. Natl. Acad. Sci. USA 1998, 95, 5857–5864. [Google Scholar]
- LaRonde-LeBlanc, N.A.; Wolberger, C. Structure of HoxA9 and Pbx1 bound to DNA: Hox hexapeptide and DNA recognition anterior to posterior. Genes Dev 2003, 17, 2060–2072. [Google Scholar]
- Pei, J.; Kim, B.H.; Grishin, N.V. PROMALS3D: A tool for multiple protein sequence and structure alignments. Nucleic Acids Res 2008, 36, 2295–2300. [Google Scholar]
- Landau, M.; Mayrose, I.; Rosenberg, Y.; Glaser, F.; Martz, E.; Pupko, T.; Ben-Tal, N. ConSurf 2005: The projection of evolutionary conservation scores of residues on protein structures. Nucleic Acids Res 2005, 33, W299–W302. [Google Scholar]
- Biegert, A.; Soding, J. Sequence context-specific profiles for homology searching. Proc. Natl. Acad. Sci. USA 2009, 106, 3770–3775. [Google Scholar]
- Ashkenazy, H.; Erez, E.; Martz, E.; Pupko, T.; Ben-Tal, N. ConSurf 2010: Calculating evolutionary conservation in sequence and structure of proteins and nucleic acids. Nucleic Acids Res 2010, 38, W529–W533. [Google Scholar]
- Tron, A.E.; Bertoncini, C.W.; Palena, C.M.; Chan, R.L.; Gonzalez, D.H. Combinatorial interactions of two amino acids with a single base pair define target site specificity in plant dimeric homeodomain proteins. Nucleic Acids Res 2001, 29, 4866–4872. [Google Scholar]
- Bru, C.; Courcelle, E.; Carrère, S.; Beausse, Y.; Dalmar, S.; Kahn, D. The ProDom database of protein domain families: More emphasis on 3D. Nucleic Acids Res 2005, 33, D212–D215. [Google Scholar]
- Cotsaftis, O.; Plett, D.; Shirley, N.; Tester, M.; Hrmova, M. A two-staged model of Na+ exclusion in rice explained by 3D modeling of HKT transporters and alternative splicing. PLoS One 2012, 7, e39865. [Google Scholar]
- Palena, C.M.; Tron, A.E.; Bertoncini, C.W.; Gonzalez, D.H.; Chan, R.L. Positively charged residues at the N-terminal arm of the homeodomain are required for efficient DNA binding by homeodomain-leucine zipper proteins. J. Mol. Biol 2001, 308, 39–47. [Google Scholar]
- Ponting, C.P.; Aravind, L. START: A lipid-binding domain in StAR, HD-ZIP and signalling proteins. Trends Biochem. Sci 1999, 24, 130–132. [Google Scholar]
- Kumar, R.; Thompson, E.B. Gene regulation by the glucocorticoid receptor: Structure:Function relationship. J. Steroid Biochem. Mol. Biol 2005, 94, 383–394. [Google Scholar]
- Szymanski, D.B.; Jilk, R.A.; Pollock, S.M.; Marks, M.D. Control of GL2 expression in Arabidopsis leaves and trichomes. Development 1998, 125, 1161–1171. [Google Scholar]
- Depege-Fargeix, N.; Javelle, M.; Chambrier, P.; Frangne, N.; Gerentes, D.; Perez, P.; Rogowsky, P.M.; Vernoud, V. Functional characterization of the HD-ZIP IV transcription factor OCL1 from maize. J. Exp. Bot 2011, 62, 293–305. [Google Scholar]
- Zhang, F.; Zuo, K.; Zhang, J.; Liu, X.; Zhang, L.; Sun, X.; Tang, K. An L1 box binding protein, GbML1, interacts with GbMYB25 to control cotton fibre development. J. Exp. Bot 2010, 61, 3599–3613. [Google Scholar]
- Sarnowski, T.J.; Ríos, G.; Jásik, J.; Œwieżewskia, S.; Kaczanowskia, S.; Lib, Y.; Kwiatkowskac, A.; Pawlikowskaa, K.; KoŸbiałc, M.; KoŸbiałc, P.; et al. SWI3 subunits of putative SWI/SNF chromatin-remodeling complexes play distinct roles during Arabidopsis development. Plant Cell 2005, 17, 2454–2472. [Google Scholar]
- Neely, K.E.; Hassan, A.H.; Brown, C.E.; Howe, L.; Workman, J.L. Transcription activator interactions with multiple SWI/SNF subunits. Mol. Cell Biol 2002, 22, 1615–1625. [Google Scholar]
- Simone, C. SWI/SNF: The crossroads where extracellular signaling pathways meet chromatin. J. Cell Physiol 2006, 207, 309–314. [Google Scholar]
- Takada, S.; Jurgens, G. Transcriptional regulation of epidermal cell fate in the Arabidopsis embryo. Development 2007, 134, 1141–1150. [Google Scholar]
- Lu, P.; Porat, R.; Nadeau, J.A.; O’Neill, S.D. Identification of a meristem L1 layer-specific gene in Arabidopsis that is expressed during embryonic pattern formation and defines a new class of homeobox genes. Plant Cell 1996, 8, 2155–2168. [Google Scholar]
- Bowman, J.L. Class III HD-Zip gene regulation, the golden fleece of ARGONAUTE activity? BioEssays 2004, 26, 938–942. [Google Scholar]
- Soppe, W.J.; Jacobsen, S.E.; Alonso-Blanco, C.; Jackson, J.P.; Kakutani, T.; Koornneef, M.; Peeters, A.J. The late flowering phenotype of FWA mutants is caused by gain-of-function epigenetic alleles of a homeodomain gene. Mol. Cell 2000, 6, 791–802. [Google Scholar]
- Kakutani, T. Genetic characterization of late-flowering traits induced by DNA hypomethylation mutation in Arabidopsis thaliana. Plant J 1997, 12, 1447–1451. [Google Scholar]
- Ikeda, Y.; Kobayashi, Y.; Yamaguchi, A.; Abe, M.; Araki, T. Molecular basis of late-flowering phenotype caused by dominant epi-alleles of the FWA locus in Arabidopsis. Plant Cell Physiol 2007, 48, 205–220. [Google Scholar]
- Fujimoto, R.; Kinoshita, Y.; Kawabe, A.; Kinoshita, T.; Takashima, K.; Nordborg, M.; Nasrallah, M.E.; Shimizu, K.K.; Kudoh, H.; Kakutani, T. Evolution and control of imprinted FWA genes in the genus Arabidopsis. PLoS Genet 2008, 4, e1000048. [Google Scholar]
- Fujimoto, R.; Sasaki, T.; Kudoh, H.; Taylor, J.M.; Kakutani, T.; Dennis, E.S. Epigenetic variation in the FWA gene within the genus Arabidopsis. Plant J 2011, 66, 831–843. [Google Scholar]
- Prigge, M.J.; Otsuga, D.; Alonso, J.M.; Ecker, J.R.; Drews, G.N.; Clark, S.E. Class III homeodomain-leucine zipper gene family members have overlapping, antagonistic, and distinct roles in Arabidopsis development. Plant Cell 2005, 17, 61–76. [Google Scholar]
- Bommert, P.; Werr, W. Gene expression patterns in the maize caryopsis: Clues to decisions in embryo and endosperm development. Gene 2001, 271, 131–142. [Google Scholar]
- Hülskamp, M.; Miséra, S.; Jürgens, G. Genetic dissection of trichome cell development in Arabidopsis. Cell 1994, 76, 555–566. [Google Scholar]
- Masucci, J.D.; Rerie, W.G.; Foreman, D.R.; Zhang, M.; Galway, M.E.; Marks, M.D.; Schiefelbein, J.W. The homeobox gene GLABRA2 is required for position-dependent cell differentiation in the root epidermis of Arabidopsis thaliana. Development 1996, 122, 1253–1260. [Google Scholar]
- Wu, Y.; Machado, A.C.; White, R.G.; Llewellyn, D.J.; Dennis, E.S. Expression profiling identifies genes expressed early during lint fibre initiation in cotton. Plant Cell Physiol 2006, 47, 107–127. [Google Scholar]
- Suo, J.; Liang, X.; Pu, L.; Zhang, Y.; Xue, Y. Identification of GhMYB109 encoding a R2R3 MYB transcription factor that expressed specifically in fiber initials and elongating fibers of cotton (Gossypium hirsutum L.). Biochim. Biophys. Acta 2003, 20, 25–34. [Google Scholar]
- Machado, A.; Wu, Y.; Yang, Y.; Llewellyn, D.J.; Dennis, E.S. The MYB transcription factor GhMYB25 regulates early fibre and trichome development. Plant J 2009, 59, 52–62. [Google Scholar]
- Ingram, G.C.; Magnard, J.L.; Vergne, P.; Dumas, C.; Rogowsky, P.M. ZmOCL1, an HD-GL2 family homeobox gene, is expressed in the outer cell layer throughout maize development. Plant Mol. Biol 1999, 40, 343–354. [Google Scholar]
- Zou, L.-P.; Sun, X.-H.; Zhang, Z.-G.; Liu, P.; Wu, J.-X.; Tian, C.-J.; Qiu, J.-L.; Lu, T.-G. Leaf rolling controlled by the homeodomain leucine zipper class IV gene Roc5 in rice. Plant Physiol 2011, 156, 1589–1602. [Google Scholar]
- Metcalfe, C.R. Anatomy of the Monocotyledons; Clarendon Press: Oxford, UK, 1960. [Google Scholar]
- Jane, W.; Chiang, S. Morphology and development of bulliform cells in arundo formosana hack. Taiwania 1991, 36, 85–97. [Google Scholar]
- Yang, J.Y.; Chung, M.C.; Tu, C.Y.; Leu, W.M. OSTF1: A HD-GL2 family homeobox gene is developmentally regulated during early embryogenesis in rice. Plant Cell Physiol 2002, 43, 628–638. [Google Scholar]
- Boutrot, F.; Meynard, D.; Guiderdoni, E.; Joudrier, P.; Gautier, M. The Triticum aestivum non-specific lipid transfer protein (TaLtp) gene family: Comparative promoter activity of six TaLtp genes in transgenic rice. Planta 2007, 225, 843–862. [Google Scholar]
- Jeffree, C.E. The Fine Structure of the Plant Cuticle. In Annual Plant Reviews Volume 23: Biology of the Plant Cuticle; Blackwell Publishing Ltd: Oxford, UK, 2007; pp. 11–125. [Google Scholar]
- Kerstiens, G. Water transport in plant cuticles: An update. J. Exp. Bot 2006, 57, 2493–2499. [Google Scholar]
- Kosma, D.; Jenks, M. Eco-Physiological and Molecular-Genetic Determinants of Plant Cuticle Function in Drought and Salt Stress Tolerance. In Advances in Molecular Breeding Toward Drought and Salt Tolerant Crops; Jenks, M., Hasegawa, P., Jain, S.M., Eds.; Springer: Dordrecht, The Netherlands, 2007; pp. 91–120. [Google Scholar]
- Burghardt, M.; Riederer, M. 9 Cuticular transpiration. In Annual Plant Reviews, Biololgy of the Plant Cuticle 23; Blackwell Publishing: Oxford, UK, 2006; Volume 292, pp. 292–312. [Google Scholar]
- Shepherd, T.; Wynne Griffiths, D. The effects of stress on plant cuticular waxes. New Phytol 2006, 171, 469–499. [Google Scholar]
- Broun, P.; Poindexter, P.; Osborne, E.; Jiang, C.-Z.; Riechmann, J.L. WIN1, a transcriptional activator of epidermal wax accumulation in Arabidopsis. Proc. Natl. Acad. Sci. USA 2004, 101, 4706–4711. [Google Scholar]
- Aharoni, A.; Dixit, S.; Jetter, R.; Thoenes, E.; van Arkel, G.; Pereira, A. The SHINE clade of ap2 domain transcription factors activates wax biosynthesis, alters cuticle properties, and confers drought tolerance when overexpressed in Arabidopsis. Plant Cell 2004, 16, 2463–2480. [Google Scholar]
- Seo, P.J.; Lee, S.B.; Suh, M.C.; Park, M.-J.; Go, Y.S.; Park, C.-M. The MYB96 transcription factor regulates cuticular wax biosynthesis under drought conditions in Arabidopsis. Plant Cell 2011, 23, 1138–1152. [Google Scholar]
- Zhang, J.Y.; Broeckling, C.D.; Sumner, L.W.; Wang, Z.Y. Heterologous expression of two Medicago truncatula putative ERF transcription factor genes, WXP1 and WXP2, in Arabidopsis led to increased leaf wax accumulation and improved drought tolerance, but differential response in freezing tolerance. Plant Mol. Biol 2007, 64, 265–278. [Google Scholar]
- Cominelli, E.; Galbiati, M.; Tonelli, C. Integration of water stress response: Cell expansion and cuticle deposition in Arabidopsis thaliana. Plant Signal Behav 2008, 3, 556–557. [Google Scholar]
- Chen, H.I.; Sudol, M. The WW domain of Yes-associated protein binds a proline-rich ligand that differs from the consensus established for Src homology 3-binding modules. Proc. Natl. Acad. Sci. USA 1995, 92, 7819–7823. [Google Scholar]
- Kagale, S.; Links, M.G.; Rozwadowski, K. Genome-wide analysis of ethylene-responsive element binding factor-associated amphiphilic repression motif-containing transcriptional regulators in Arabidopsis. Plant Physiol 2010, 152, 1109–1134. [Google Scholar]
- Hiratsu, K.; Matsui, K.; Koyama, T.; Ohme-Takagi, M. Dominant repression of target genes by chimeric repressors that include the EAR motif, a repression domain, in Arabidopsis. Plant J 2003, 34, 733–739. [Google Scholar]
- Javelle, M.; Vernoud, V.; Depege-Fargeix, N.; Arnould, C.; Oursel, D.; Domergue, F.; Sarda, X.; Rogowsky, P.M. Overexpression of the epidermis-specific homeodomain-leucine zipper IV transcription factor Outer Cell Layer1 in maize identifies target genes involved in lipid metabolism and cuticle biosynthesis. Plant Physiol 2010, 154, 273–286. [Google Scholar]
- Rowland, O.; Zheng, H.; Hepworth, S.R.; Lam, P.; Jetter, R.; Kunst, L. CER4 encodes an alcohol-forming fatty acyl-coenzyme a reductase involved in cuticular wax production in Arabidopsis. Plant Physiol 2006, 142, 866–877. [Google Scholar]
- Cummins, I.; Landrum, M.; Steel, P.; Edwards, R. Structure activity studies with xenobiotic substrates using carboxylesterases isolated from Arabidopsis thaliana. Phytochemistry 2007, 68, 811–818. [Google Scholar]
- Frey, M.; Stettner, C.; Paré, P.W.; Schmelz, E.A.; Tumlinson, J.H.; Gierl, A. An herbivore elicitor activates the gene for indole emission in maize. Proc. Natl. Acad. Sci. USA 2000, 97, 14801–14806. [Google Scholar]
- Curwin, A.J.; Fairn, G.D.; McMaster, C.R. Phospholipid transfer protein Sec14 is required for trafficking from endosomes and regulates distinct trans-Golgi export pathways. J. Biol. Chem 2009, 284, 7364–7375. [Google Scholar]
- Bankaitis, V.A.; Phillips, S.; Yanagisawa, L.; Li, X.; Routt, S.; Xie, Z. Phosphatidylinositol transfer protein function in the yeast Saccharomyces cerevisiae. Adv. Enzyme Regul 2005, 45, 155–170. [Google Scholar]
- Velamakanni, S.; Wei, S.L.; Janvilisri, T.; van Veen, H.W. ABCG transporters: Structure, substrate specificities and physiological roles: A brief overview. J. Bioenerg. Biomembr 2007, 39, 465–471. [Google Scholar]
- Pighin, J.A.; Zheng, H.; Balakshin, L.J.; Goodman, I.P.; Western, T.L.; Jetter, R.; Kunst, L.; Samuels, A.L. Plant cuticular lipid export requires an ABC transporter. Science 2004, 306, 702–704. [Google Scholar]
- Bird, D.; Beisson, F.; Brigham, A.; Shin, J.; Greer, S.; Jetter, R.; Kunst, L.; Wu, X.; Yephremov, A.; Samuels, L. Characterization of Arabidopsis ABCG11/WBC11, an ATP binding cassette (ABC) transporter that is required for cuticular lipid secretion. Plant J 2007, 52, 485–498. [Google Scholar]
- Kader, J. Lipid-transfer proteins in plants. Annu. Rev. Plant Physiol. Plant Mol. Biol 1996, 47, 627–654. [Google Scholar]
- DeBono, A.; Yeats, T.H.; Rose, J.K.C.; Bird, D.; Jetter, R.; Kunst, L.; Samuels, L. Arabidopsis LTPG is a glycosylphosphatidylinositol-anchored lipid transfer protein required for export of lipids to the plant surface. Plant Cell 2009, 21, 1230–1238. [Google Scholar]
- Lee, S.B.; Go, Y.S.; Bae, H.J.; Park, J.H.; Cho, S.H.; Cho, H.J.; Lee, D.S.; Park, O.K.; Hwang, I.; Suh, M.C. Disruption of glycosylphosphatidylinositol-anchored lipid transfer protein gene altered cuticular lipid composition, increased plastoglobules, and enhanced susceptibility to infection by the fungal pathogen Alternaria brassicicola. Plant Physiol 2009, 150, 42–54. [Google Scholar]
- Boutrot, F.; Chantret, N.; Gautier, M.-F. Genome-wide analysis of the rice and Arabidopsis non-specific lipid transfer protein (nsLtp) gene families and identification of wheat nsLtp genes by EST data mining. BMC Genomics 2008, 9, 86, :1–86:19.. [Google Scholar]
- Zottich, U.; da Cunha, M.; Carvalho, A.O.; Dias, G.B.; Silva, N.C.; Santos, I.S.; do Nacimento, V.V.; Miguel, E.C.; Machado, O.L.; Gomes, V.M. Purification, biochemical characterization and antifungal activity of a new lipid transfer protein (LTP) from Coffea canephora seeds with alpha-amylase inhibitor properties. Biochim. Biophys. Acta 2011, 4, 375–383. [Google Scholar]
- Molina, A.; Garcia-Olmedo, F. Enhanced tolerance to bacterial pathogens caused by the transgenic expression of barley lipid transfer protein LTP2. Plant J 1997, 12, 669–675. [Google Scholar]
- Lay, F.T.; Mills, G.D.; Poon, I.K.; Cowieson, N.P.; Kirby, N.; Baxter, A.A.; van der Weerden, N.L.; Dogovski, C.; Perugini, M.A.; Anderson, M.A.; et al. Dimerization of plant defensin NaD1 enhances its antifungal activity. J. Biol. Chem 2012, 287, 19961–19972. [Google Scholar]
- Rigano, M.M.; Romanelli, A.; Fulgione, A.; Nocerino, N.; D’Agostino, N.; Avitabile, C.; Frusciante, L.; Barone, A.; Capuano, F.; Capparelli, R. A novel synthetic peptide from a tomato defensin exhibits antibacterial activities against Helicobacter pylori. J. Pept. Sci 2012, 18, 755–762. [Google Scholar]
- Picart, P.; Pirttila, A.; Raventos, D.; Kristensen, H.-H.; Sahl, H.-G. Identification of defensin-encoding genes of Picea glauca: Characterization of PgD5, a conserved spruce defensin with strong antifungal activity. BMC Plant Biol 2012, 12, 180, :1–180:14.. [Google Scholar]
- Kovalchuk, N.; Li, M.; Wittek, F.; Reid, N.; Singh, R.; Shirley, N.; Ismagul, A.; Eliby, S.; Johnson, A.; Milligan, A.S.; et al. Defensin promoters as potential tools for engineering disease resistance in cereal grains. Plant Biotech. J 2010, 8, 47–64. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Plant | Gene name | Accession | Tissue localisation | Suggested function* | Cited |
|---|---|---|---|---|---|
| Arabidopsis thaliana | HDG1 | NP_191674 | In trichomes at the base of young leaf, endodermal cell lines around emergent lateral root, stamen filament | Cuticle development | [24,29] |
| Arabidopsis thaliana | HDG2 | NP_172015 | Outer cell layers of shoot apical meristems, early flower primordia, nucellus, seed coat, epidermal layers of young leaves, hairless cell files of hypocotyl epidermis, primary root tips | Embryo development | [24] |
| Arabidopsis thaliana | HDG3 | NP_180796 | Siliques and seedlings | Cotyledon development | [24] |
| Arabidopsis thaliana | HDG4 | NP_193506 | Flowers | Flower organ development | [24] |
| Arabidopsis thaliana | HDG5 | NP_199499 | All plant organs except root, outer cell layers of shoot apical meristems, early flower primordia, nucellus, epidermis of the stamen filament, stomatal guard cells of the carpel | Not determined | [24] |
| Arabidopsis thaliana | FWA/HDG6 | NP_567722 | Endosperm | Endosperm development | [30] |
| Arabidopsis thaliana | HDG7 | NP_200030 | Base of leaf primordia, apical region of the heart-stage embryo, lateral root primordia and tips, seedlings | Not determined | [24] |
| Arabidopsis thaliana | HDG8 | NP_186976 | Endosperm and embryo at early stages of development, after flower organ development | Flower, seed development | [24] |
| Arabidopsis thaliana | HDG9 | NP_197234 | Developed flowers, chalazal, embryo sac | Embryonic development | [24] |
| Arabidopsis thaliana | HDG12 | NP_564041 | Apical meristem, stamen filament, nucellus, embryo, young epidermal tissues, lateral root tip | Trichome development | [24] |
| Arabidopsis thaliana | GL2 | NP_565223 | Epidermis of leaves and roots, developing trichomes and surrounding epidermal cells, hairless cell files of hypocotyl epidermis | Trichome and root hair development | [31] |
| Arabidopsis thaliana | ANL2 | NP_567183 | Leaves, stems, buds, roots | Anthocyanin distribution and root development | [32] |
| Arabidopsis thaliana | ATML1 | NP_193906 | Flower bud, lower parts of siliques | Shoot epidermal cell differentiation | [33] |
| Arabidopsis thaliana | PDF2 | NP_567274 | Flower buds, shoot apices | Shoot epidermal cell differentiation, cotyledon development | [33] |
| Picea abies | PaHB1 | AAG43405 | Embryo | Embryo development | [34] |
| Picea abies | PaHB2 | AAL83725 | Embryo | Embryo development | [35] |
| Gossypium spp. | GaHOX1 | ABY41242 | Fibre cells | Cotton fibre development | [36] |
| Gossypium spp. | GaHOX2 | ABY67263 | Fibre cells at early developmental stages | Seed coat development | [36] |
| Gossypium hirsutum | GhHD-1A | AFO11041 | Epidermal cells | Epidermal cell determination | [37] |
| Gossypium hirsutum | GhHD-1D | AFO11042 | Epidermal cells | Epidermal cell determination | [37] |
| Zea mays | ZmOCL1 | CAG38614 | Immature ears, epidermis specific expression in embryo, meristems, young organ primordia, juvenile leaves | Kernel development, cuticle deposition | [7] |
| Zea mays | ZmOCL2 | CAB96422 | Apical meristem, sub-epidermal layer of floral meristems immature ears, young female gametophyte, kernels, root | Not determined | [38] |
| Zea mays | ZmOCL3 | CAB96423 | Developing embryo and endosperm, epidermis specific expression in embryo, meristems, young organ primordial, juvenile leaves | Specification of organ identity | [38] |
| Zea mays | ZmOCL4 | CAB96424.2 | Shoot epidermis, meristems and young organ primordia, immature tassels | Anther and trichome development | [39] |
| Zea mays | ZmOCL5 | CAB96425 | Immature tassels, epidermis specific expression in embryo, meristems, young organ primordia, | Not determined | [40] |
| Zea mays | ZmOCL6 | DAA34955 | Immature tassels, immature ears, epidermal cells of juvenile leaves | Not determined | [40] |
| Zea mays | ZmOCL7 | DAA34956 | Immature ears, epidermal cells | Not determined | [40] |
| Zea mays | ZmOCL8 | DAA34957 | Immature tassels, epidermal cells | Not determined | [40] |
| Zea mays | ZmOCL9 | DAA34958 | Young developing kernels, L2 cells of shoot apical meristem | Not determined | [40] |
| Zea mays | ZmOCL10 | DAA34959 | Immature tassels, epidermal layer of juvenile leaves | Not determined | [40] |
| Zea mays | ZmOCL11 | DAA34960 | Mature tassels that include pollen | Not determined | [40] |
| Zea mays | ZmOCL12 | DAA34961 | Not determined | Not determined | [40] |
| Zea mays | ZmOCL13 | DAA34962 | Immature tassels, epidermis of shoot apical meristem | Not determined | [40] |
| Zea mays | ZmOCL14 | DAA34963 | Immature tassels, epidermal cells | Not determined | [40] |
| Zea mays | ZmOCL15 | DAA34964 | Immature tassels, epidermis of shoot apical meristem | Not determined | [40] |
| Zea mays | ZmOCL16 | DAA34965 | Young developing kernels, epidermal cells | Not determined | [40] |
| Zea mays | ZmOCL17 | DAA34966 | Immature tassels, epidermis of shoot apical meristem | Not determined | [40] |
| Oryza sativa | OsRoc1 | BAB85750 | Shoot apex, inflorescence apex, leaf blade | Embryogenesis | [41] |
| Oryza sativa | OsRoc2 | BAC77155 | Shoot apex, inflorescence apex, leaf blade | Not determined | [42] |
| Oryza sativa | OsRoc3 | BAC77156 | Shoot apex, inflorescence | Not determined | [42] |
| Oryza sativa | OsRoc4 | BAC77157 | Roots | Not determined | [42] |
| Oryza sativa | OsRoc5 | BAC77158 | Leaf epidermis | Epidermal differentiation, bulliform cells development | [42] |
| Oryza sativa | OsROC6 | Q7Y0V7 | Not determined | [42] | |
| Oryza sativa | OsRoc7 | BAC77160 | Epidermis | Epidermal differentiation | [42] |
| Oryza sativa | OsRoc8 | BAC77161 | Epidermis | Epidermal differentiation | [42] |
| Oryza sativa | OsRoc9 | BAC77162 | Epidermis | Epidermal differentiation | [42] |
| Oryza sativa | OsTF1 | Q5ZAY0 | Grain | Embryogenesis | [43] |
| Oryza sativa | OsHOX9 | A2Z8L4 | Not determined | [44] | |
| Triticum aestivum | TaGL9 | AEI99592 | Grain specific expression | Not determined | [45] |
| Triticum durum | TdGL9H | AEI99593 | Early embryo, endosperm around embryo, later in scutellar vascular bundle | Maintenance of scutellar vascular bundle | [45] |
| Solanum lycopersicum | SlCD2 | NP_001234657 | Epidermal cell and cuticle | Epidermal cell and cuticle development | [20] |
| Physcomitrella patens | PpHDZ41 | DAA05775 | Not determined | Not determined | [46] |
| Selaginella moellendorffii | SmHDZ44 | DAA05774 | Not determined | Not determined | [46] |
© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Chew, W.; Hrmova, M.; Lopato, S. Role of Homeodomain Leucine Zipper (HD-Zip) IV Transcription Factors in Plant Development and Plant Protection from Deleterious Environmental Factors. Int. J. Mol. Sci. 2013, 14, 8122-8147. https://doi.org/10.3390/ijms14048122
Chew W, Hrmova M, Lopato S. Role of Homeodomain Leucine Zipper (HD-Zip) IV Transcription Factors in Plant Development and Plant Protection from Deleterious Environmental Factors. International Journal of Molecular Sciences. 2013; 14(4):8122-8147. https://doi.org/10.3390/ijms14048122
Chicago/Turabian StyleChew, William, Maria Hrmova, and Sergiy Lopato. 2013. "Role of Homeodomain Leucine Zipper (HD-Zip) IV Transcription Factors in Plant Development and Plant Protection from Deleterious Environmental Factors" International Journal of Molecular Sciences 14, no. 4: 8122-8147. https://doi.org/10.3390/ijms14048122
APA StyleChew, W., Hrmova, M., & Lopato, S. (2013). Role of Homeodomain Leucine Zipper (HD-Zip) IV Transcription Factors in Plant Development and Plant Protection from Deleterious Environmental Factors. International Journal of Molecular Sciences, 14(4), 8122-8147. https://doi.org/10.3390/ijms14048122