Molecular Cloning and Characterization of the First Caspase in the Striped Stem Borer, Chilo suppressalis

Abstract
:1. Introduction
2. Results and Discussion
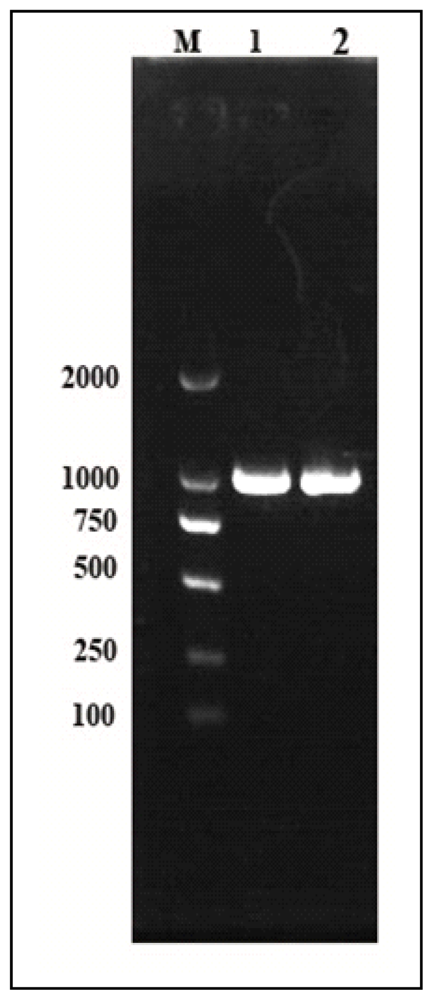
2.1. Isolation, Cloning, Sequencing and Structure of Cscaspase-1
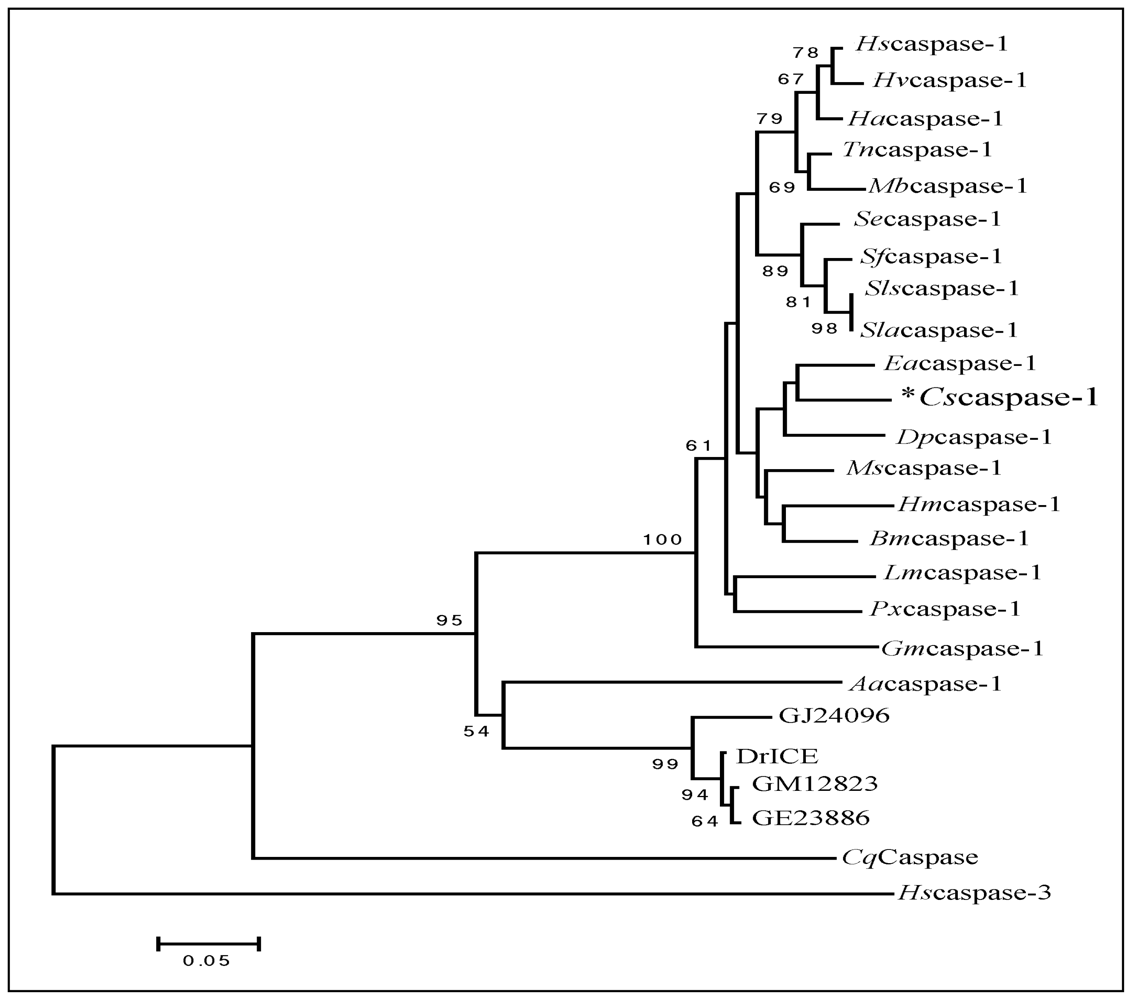
2.2. Phylogenetic Analysis
2.3. Genomic Analysis of Cscaspase-1
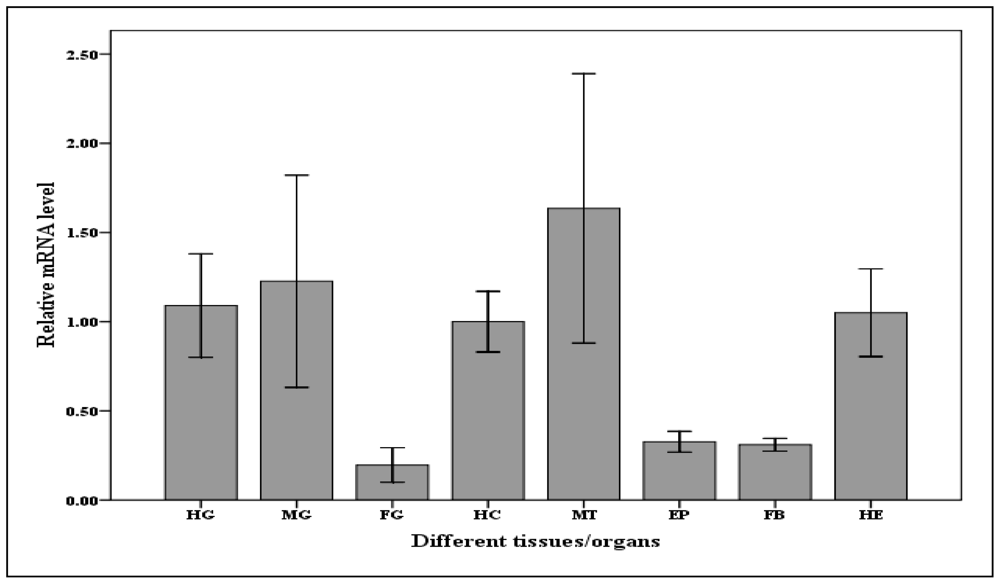
2.4. Tissues (organs) Distribution of Cscaspase-1mRNA Expression in C. suppressalis Larvae
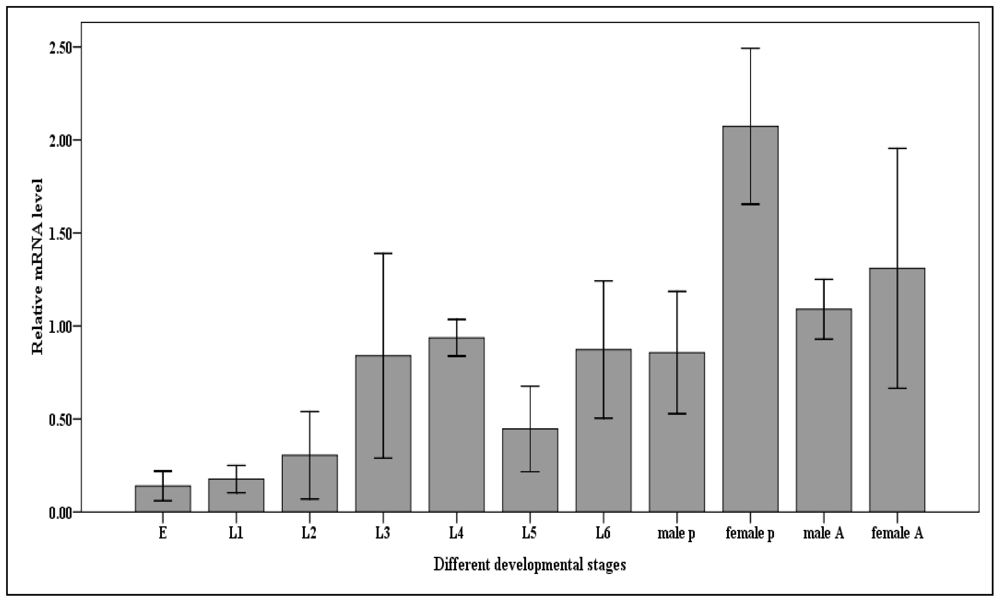
2.5. Expression of Cscaspase-1 in the Developmental Stages of the C. suppressalis
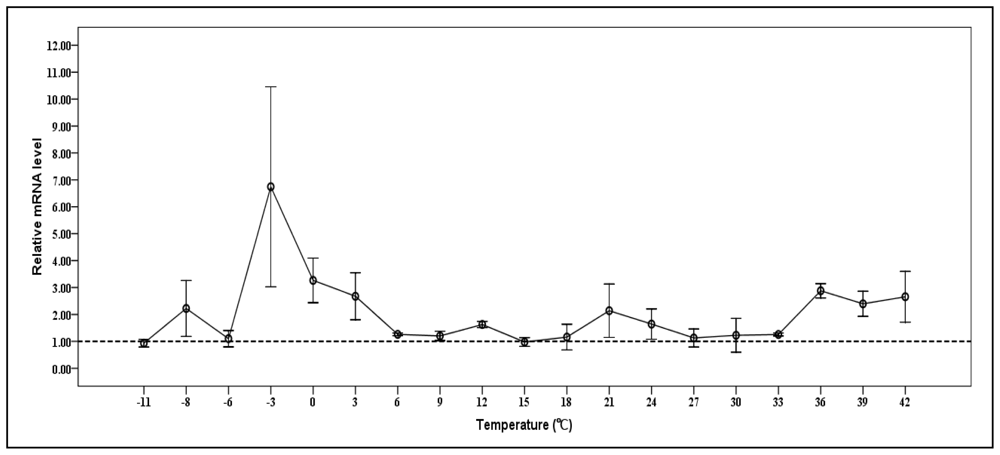
2.6. Expression of Cscaspase-1 mRNA under Various Temperatures
3. Experimental Section
3.1. Insects
3.2. Cloning and RACEs
3.3. Characterization of the Cscaspase-1 Genome
3.4. Sample Preparation
3.5. Temperature Stress
3.6. Quantitative Real-time PCR (qPCR) Analysis
3.7. Bioinformatic Analysis
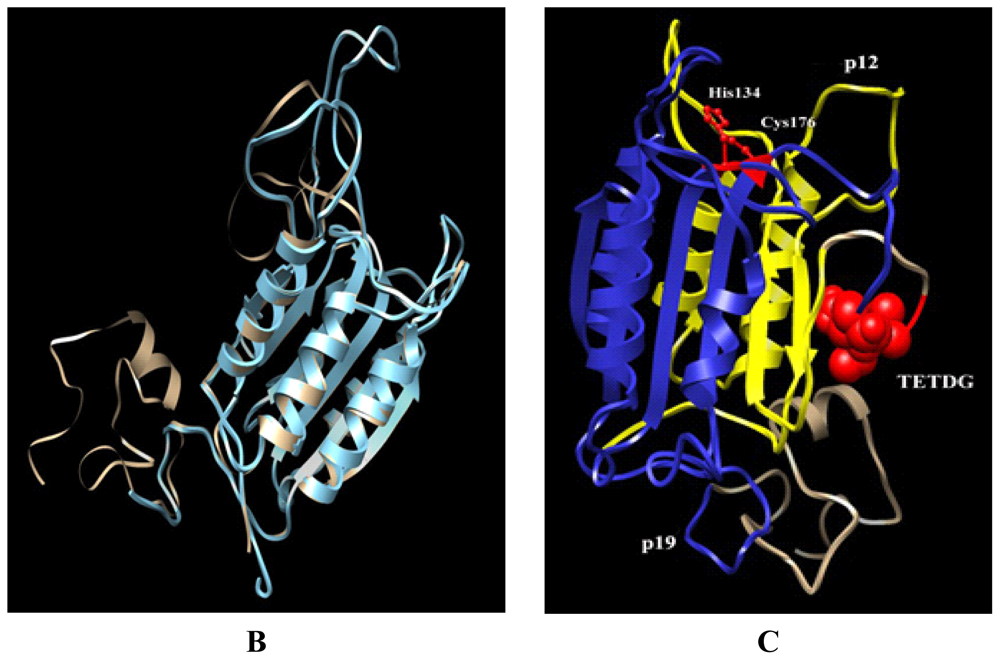
3.8. Computational Molecular Modeling
4. Conclusions
Supplementary Information
ijms-14-10229-s001.pdfAcknowledgments
References
- Kerr, J.F.R.; Wyllie, H.A.; Currie, R.A. Apoptosis: A basic biological phenomenon with wide-ranging Implications in Tissue Kinetics. Br. J. Cancer 1972, 26, 239–257. [Google Scholar]
- White, E. Life, death, and the pursuit of apoptosis. Gene. Dev 1996, 10, 1–15. [Google Scholar]
- Teodoro, J.G.; Branton, P.E. Regulation of apoptosis by viral gene products. J. Virol 1997, 71, 1739–1746. [Google Scholar]
- Benedict, C.A.; Norris, P.S.; Ware, C.F. To kill or be killed: Viral evasion of apoptosis. Nat. Immunol 2002, 3, 1013–1018. [Google Scholar]
- Yi, S.X.; Moore, C.W.; Lee, R.E., Jr. Rapid cold-hardening protects Drosophila melanogaster from cold-induced apoptosis. Apoptosis 2007, 12, 1183–1193. [Google Scholar]
- Cohen, G.M. Caspases: The executioners of apoptosis. Biochem. J 1997, 326, 1–16. [Google Scholar]
- Thornberry, N.A.; Lazebnik, Y. Caspases: Enemies within. Science 1998, 281, 1312–1316. [Google Scholar]
- Raff, M. Cell suicide for beginners. Nature 1998, 396, 119–122. [Google Scholar]
- Clem, R.J. The Role of Apoptosis in Defense against Baculovirus Infection in Insects. In Role of Apoptosis in Infection: Current Topics in Microbiology and Immunology; Griffin, D., Ed.; Springer: Heidelberg, Germany, 2005; Volume 289, pp. 113–129. [Google Scholar]
- Richardson, H.; Kumar, S. Death to flies: Drosophila as a model system to study programmed cell death. J. Immunol. Methods 2002, 265, 21–38. [Google Scholar]
- Cooper, D.M.; Thi, E.P.; Chamberlain, C.M.; Pio, F.; Lowenberger, C. Aedes Dronc: A novel ecdysone-inducible caspase in the yellow fever mosquito, Aedes aegypti. Insect Mol. Biol 2007, 16, 563–572. [Google Scholar]
- Zhang, J.Y.; Pan, M.H.; Sun, Z.Y.; Huang, S.J.; Yu, Z.S.; Liu, D.; Zhao, D.H.; Lu, C. The genomic underpinnings of apoptosis in the silkworm, Bombyx mori. BMC Genomics 2010, 11, 611. [Google Scholar]
- Courtiade, J.; Pauchet, Y.; Vogel, H.; Heckel, D.G. A comprehensive characterization of the caspase gene family in insects from the order Lepidoptera. BMC Genomics 2011, 12, 357. [Google Scholar]
- Forsyth, C.M.; Lemongello, D.; LaCount, D.J.; Friesen, P.D.; Fisher, A.J. Crystal Structure of an Invertebrate Caspase. J. Biol. Chem 2004, 279, 7001–7008. [Google Scholar]
- Yang, D.T.; Chai, L.Q.; Wang, J.X.; Zhao, X.F. Molecular cloning and characterization of Hearm caspase-1 from Helicoverpa armigera. Mol. Biol. Rep 2008, 35, 405–412. [Google Scholar]
- Zhuang, H.M.; Wang, K.F.; Miyata, T.; Wu, Z.J.; Wu, G.; Xie, L.H. Identification and expression of caspase-1 gene under heat stress in insecticide susceptible and-resistant Plutella xylostella (Lepidoptera: Plutellidae). Mol. Biol. Rep 2011, 38, 2529–2539. [Google Scholar]
- Khoa, D.B.; Trang, L.T.D.; Takeda, M. Expression analyses of caspase-1 and related activities in the midgut of Galleria mellonella during metamorphosis. Insect Mol. Biol 2012, 21, 247–256. [Google Scholar]
- Bowler, K.; Terblanche, J.S. Insect thermal tolerance: What is the role of ontogeny, ageing and senescence? Biol. Rev 2008, 83, 339–355. [Google Scholar]
- Doeppner, T.R.; Grune, T.; de Groot, H.; Rauen, U. Cold induced apoptosis of rat liver endothelial cells: Involvement of the proteasome. Transplantation 2003, 75, 1946–1953. [Google Scholar]
- Lu, M.X.; Liu, Z.X.; Wang, X.; Du, Y.Z. Seasonal Cold Tolerance of Chilo suppressalis (Walker) in Yangzhou, China. Ann. Entomol. Soc. Am 2012, 105, 479–483. [Google Scholar]
- Hurst, L.D.; McVean, G.; Moore, T. Imprinted genes have few and small introns. Nat. Genet 1996, 12, 234–237. [Google Scholar]
- Castillo-Davis, C.I.; Mekhedov, S.L.; Hartl, D.L.; Koonin, E.V.; Kondrashov, F.A. Selection for short introns in highly expressed genes. Nat. Genet 2002, 31, 415–418. [Google Scholar]
- Comeron, J.M. Selective and mutational patterns associated with gene expression in humans: Influences on synonymous composition and intron presence. Genetics 2004, 167, 1293–1304. [Google Scholar]
- Penaloza, C.; Orlanski, S.; Ye, Y.; Entezari-Zaher, T.; Javdan, M.; Zakeri, Z. Cell death in mammalian development. Curr. Pharm. Des 2008, 14, 184–196. [Google Scholar]
- Amundson, S.A.; Myers, T.G.; Fornace, A.J. Roles for p53 in growth arrest and apoptosis: Putting on the brakes after genotoxic stress. Oncogene 1998, 17, 3287–3299. [Google Scholar]
- McGavin, G.C. Essential Entomology: An Order by Order Introduction; Oxford University Press: New York, NY, USA, 2001. [Google Scholar]
- Uwo, M.F.; Ui-Tei, K.; Park, P.; Takeda, M. Replacement of midgut epithelium in the greater wax moth, Galleria mellonela, during larval–pupal moult. Cell Tissue Res 2002, 308, 319–331. [Google Scholar]
- Tettamanti, G.; Grimaldi, A.; Casartelli, M.; Ambrosetti, E.; Ponti, B.; Congiu, T.; Ferrarese, R.; Rivas-Pena, M.L.; Pennacchio, F.; Eguileor, M. Programmed cell death and stem cell differentiation are responsible for midgut replacement in Heliothis virescens during prepupal instar. Cell Tissue Res 2007, 330, 345–359. [Google Scholar]
- Vilaplana, L.; Pascual, N.; Perera, N.; Belles, X. Molecular characterization of an inhibitor of apoptosis in the Egyptian armyworm, Spodoptera littoralis, and midgut cell death during metamorphosis. Insect Biochem. Mol. Biol 2007, 37, 1241–1248. [Google Scholar]
- Hakim, R.S.; Baldwin, K.; Smagghe, G. Regulation of midgut growth, development, and metamorphosis. Annu. Rev. Entomol 2010, 55, 593–608. [Google Scholar]
- Kilpatrick, Z.E.; Cakouros, D.; Kumar, S. Ecdysone-mediated up-regulation of the effector caspase DRICE is required for hormone-dependent apoptosis in Drosophila cells. J. Biol. Chem 2005, 280, 981–986. [Google Scholar]
- Yin, V.P.; Thummel, C.S. Mechanisms of steroid-triggered programmed cell death in Drosophila. Semin. Cell Dev. Biol 2005, 16, 237–243. [Google Scholar]
- Baehrecke, E.H. Steroid regulation of programmed cell death during Drosophila development. Cell Death Differ 2000, 7, 1057–1062. [Google Scholar]
- Jiang, C.; Jiang, C.; Lamblin, A.F.; Steller, H.; Thummel, C.S. A steroid-triggered transcriptional hierarchy controls salivary gland cell death during Drosophila metamorphosis. Mol. Cell 2000, 5, 445–455. [Google Scholar]
- Lee, C.Y.; Cooksey, B.; Baehrecke, E.H. Steroid regulation of midgut cell death during Drosophila development. Dev. Biol 2002, 250, 101–111. [Google Scholar]
- Shang, Z.Z.; Wang, Y.S.; Zou, Y.H. Study on rearing method of rice stem borer Chilo suppressalis Walker (in Chinese). Acta Entomol. Sin 1979, 2, 164–167. [Google Scholar]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative CT method. Nat. Protoc 2008, 3, 1101–1108. [Google Scholar]
- ORF Finder. Available online: http://www.ncbi.nlm.nih.gov/gorf/gorf.html (accessed on 13 May 2013).
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular Evolutionary Genetics Analysis using Maximum Likelihood, Evolutionary Distance, and Maximum Parsimony Methods. Mol. Biol. Evol 2011, 28, 2731–2739. [Google Scholar]
- Kelley, L.A.; Sternberg, M.J.E. Protein structure prediction on the web: A case study using the phyre server. Nat. Protoc 2009, 4, 363–371. [Google Scholar]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera–a visualization system for exploratory research and analysis. J. Comput. Chem 2004, 25, 1605–1612. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primers | Sequences (5′–3′) |
|---|---|
| DP-F | AYCAYGARCATTTYGAHATTCACA |
| DP-R | DCCACCATCCRATTTATCACCTTG |
| caspase-1 5′ | CCTGTGCGAGATTTCAAGCTGTGA |
| caspase-1 3′ | ACTTTACTGCTGACAAGTGCCCTTCC |
| caspase-1-F | TGCATGTCATTTTCGGTTGTTG |
| caspase-1-R | CTCCGCCGTTTTCCCTTTG |
| GSP-1 | CGAAATGTCTGACGAAG |
| GSP-2 | AAACTAGGGCAAACTCA |
| 18SrRNA-F | CACGGGAAATCTCACCAGG |
| 18SrRNA-R | CAGACAAATCGCTCCACCAACTA |
| UPM(Long) | CTAATACGACTCACTATAGGGCAAGCAGTGGTATCAACGCAGAGT |
| UPM(Short) | CTAATACGACTCACTATAGGGC |
© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Lu, M.-X.; Du, Y.-Z.; Cao, S.-S.; Liu, P.; Li, J. Molecular Cloning and Characterization of the First Caspase in the Striped Stem Borer, Chilo suppressalis. Int. J. Mol. Sci. 2013, 14, 10229-10241. https://doi.org/10.3390/ijms140510229
Lu M-X, Du Y-Z, Cao S-S, Liu P, Li J. Molecular Cloning and Characterization of the First Caspase in the Striped Stem Borer, Chilo suppressalis. International Journal of Molecular Sciences. 2013; 14(5):10229-10241. https://doi.org/10.3390/ijms140510229
Chicago/Turabian StyleLu, Ming-Xing, Yu-Zhou Du, Shuang-Shuang Cao, Pingyang Liu, and Jianyong Li. 2013. "Molecular Cloning and Characterization of the First Caspase in the Striped Stem Borer, Chilo suppressalis" International Journal of Molecular Sciences 14, no. 5: 10229-10241. https://doi.org/10.3390/ijms140510229
APA StyleLu, M. -X., Du, Y. -Z., Cao, S. -S., Liu, P., & Li, J. (2013). Molecular Cloning and Characterization of the First Caspase in the Striped Stem Borer, Chilo suppressalis. International Journal of Molecular Sciences, 14(5), 10229-10241. https://doi.org/10.3390/ijms140510229