Physiological, Biochemical, and Molecular Mechanisms of Heat Stress Tolerance in Plants

 ,
, 
 and
and 
Abstract
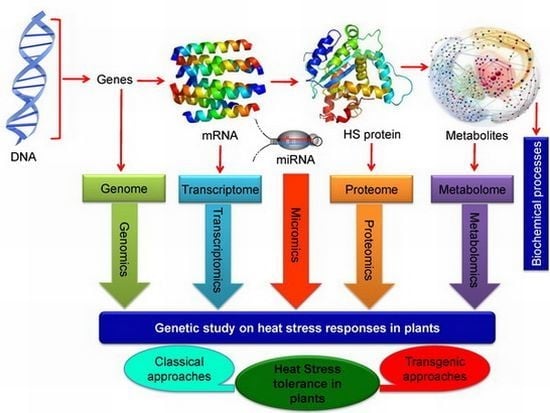
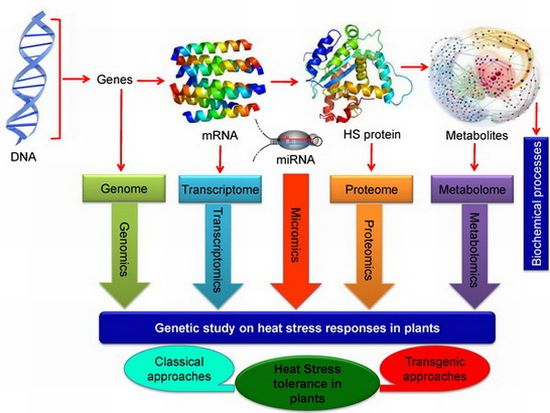
:
1. Introduction
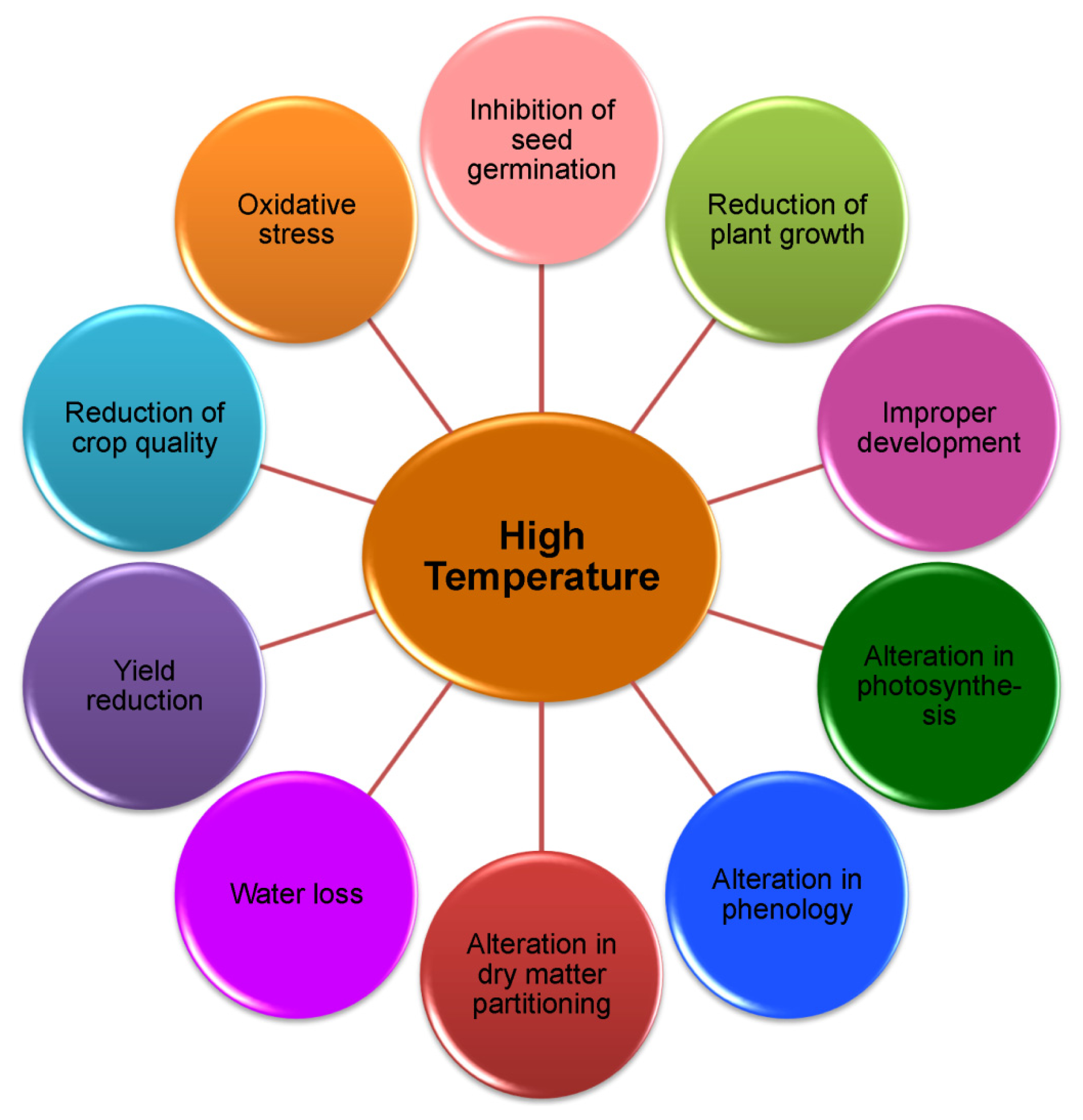
2. Plant Response to Heat Stress
2.1. Growth
2.2. Photosynthesis
2.3. Reproductive Development
2.4. Yield
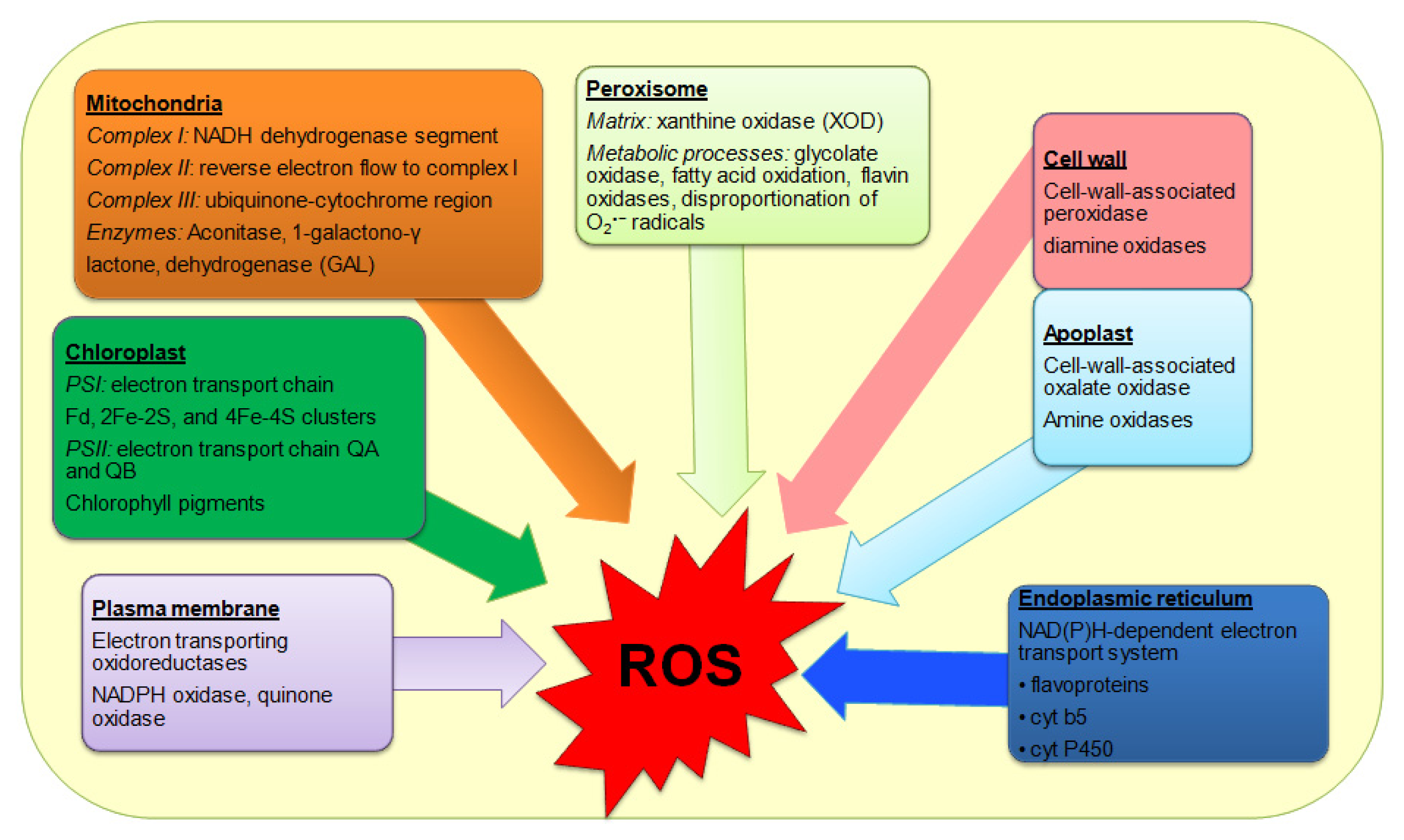
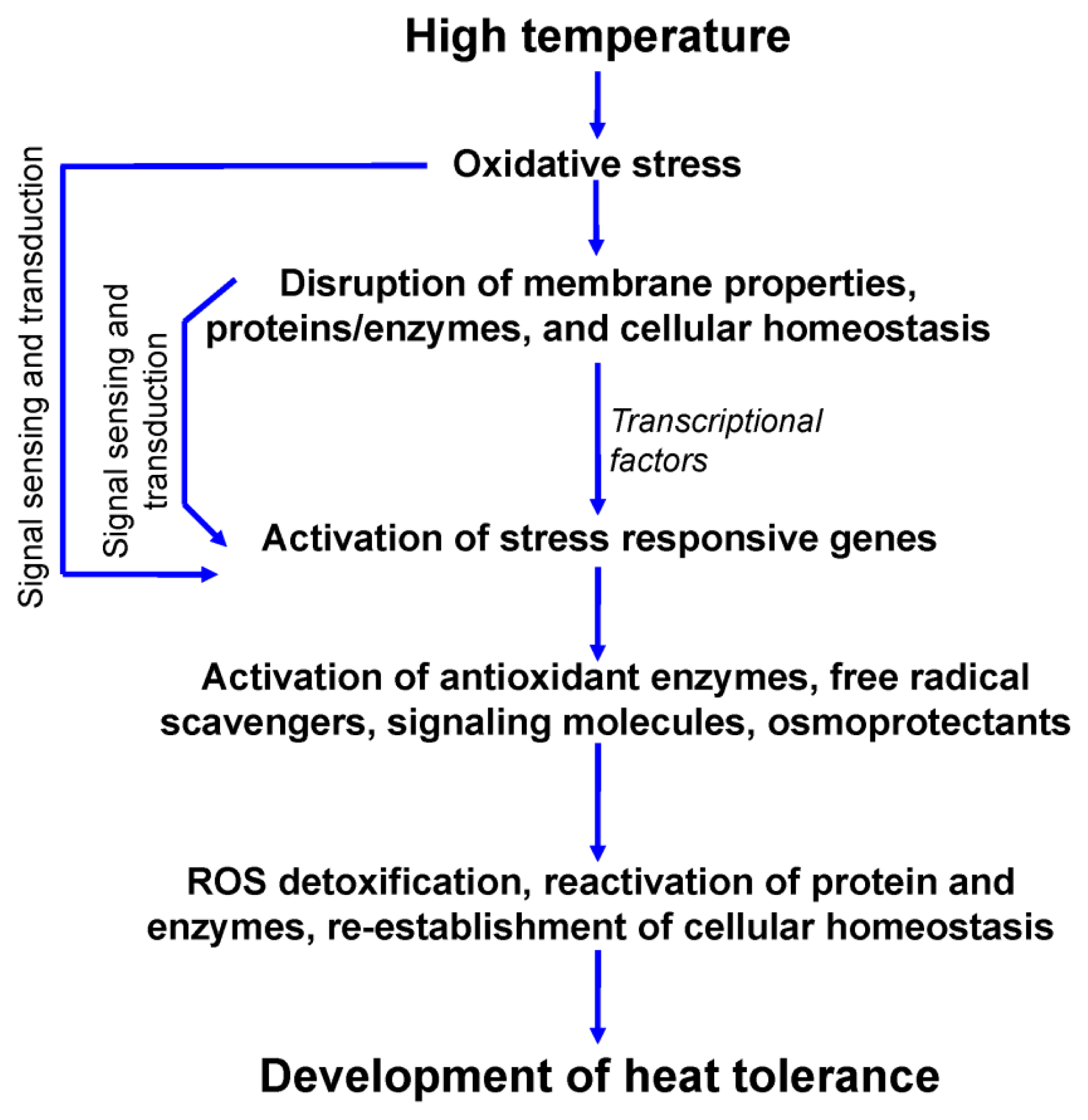
2.5. Oxidative Stress
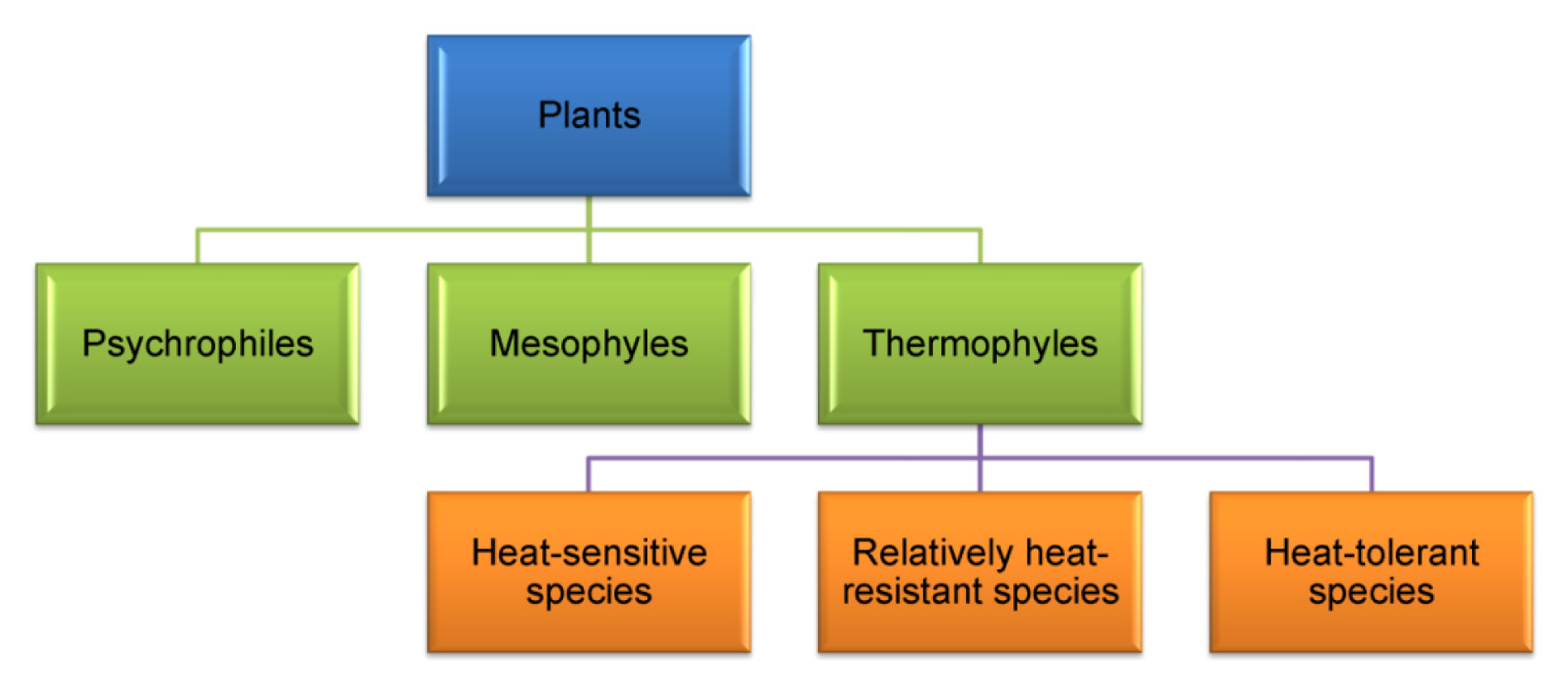
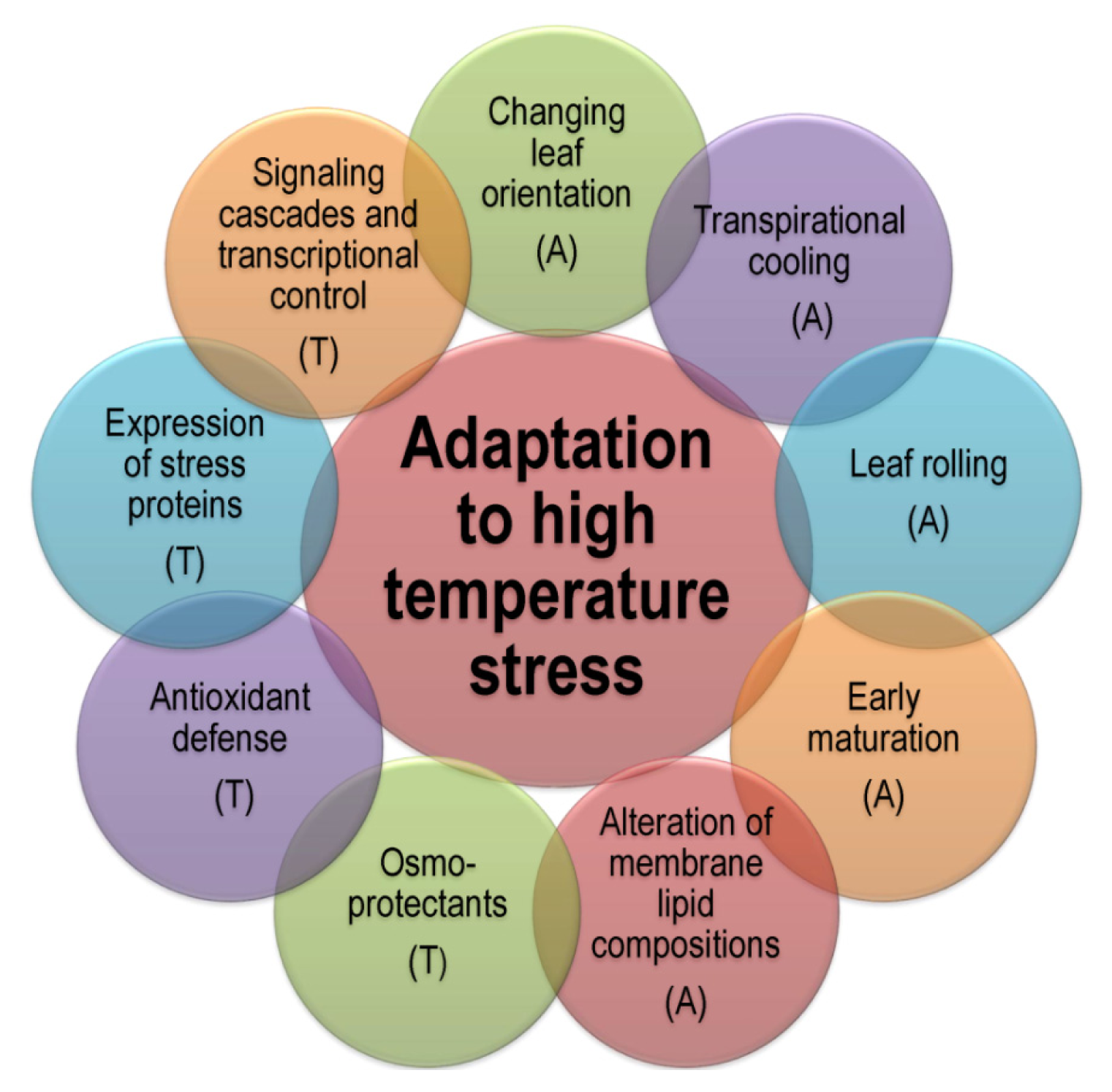
3. Plant Adaptation to Heat Stress
3.1. Avoidance Mechanisms
3.2. Tolerance Mechanisms
4. Antioxidant Defense in Response to Heat-Induced Oxidative Stress
5. Mechanism of Signal Transduction and Development of Heat Tolerance
6. Use of Exogenous Protectants in Mitigating Heat-Induced Damages
7. Molecular and Biotechnological Strategies for Development of Heat Stress Tolerance in Plants
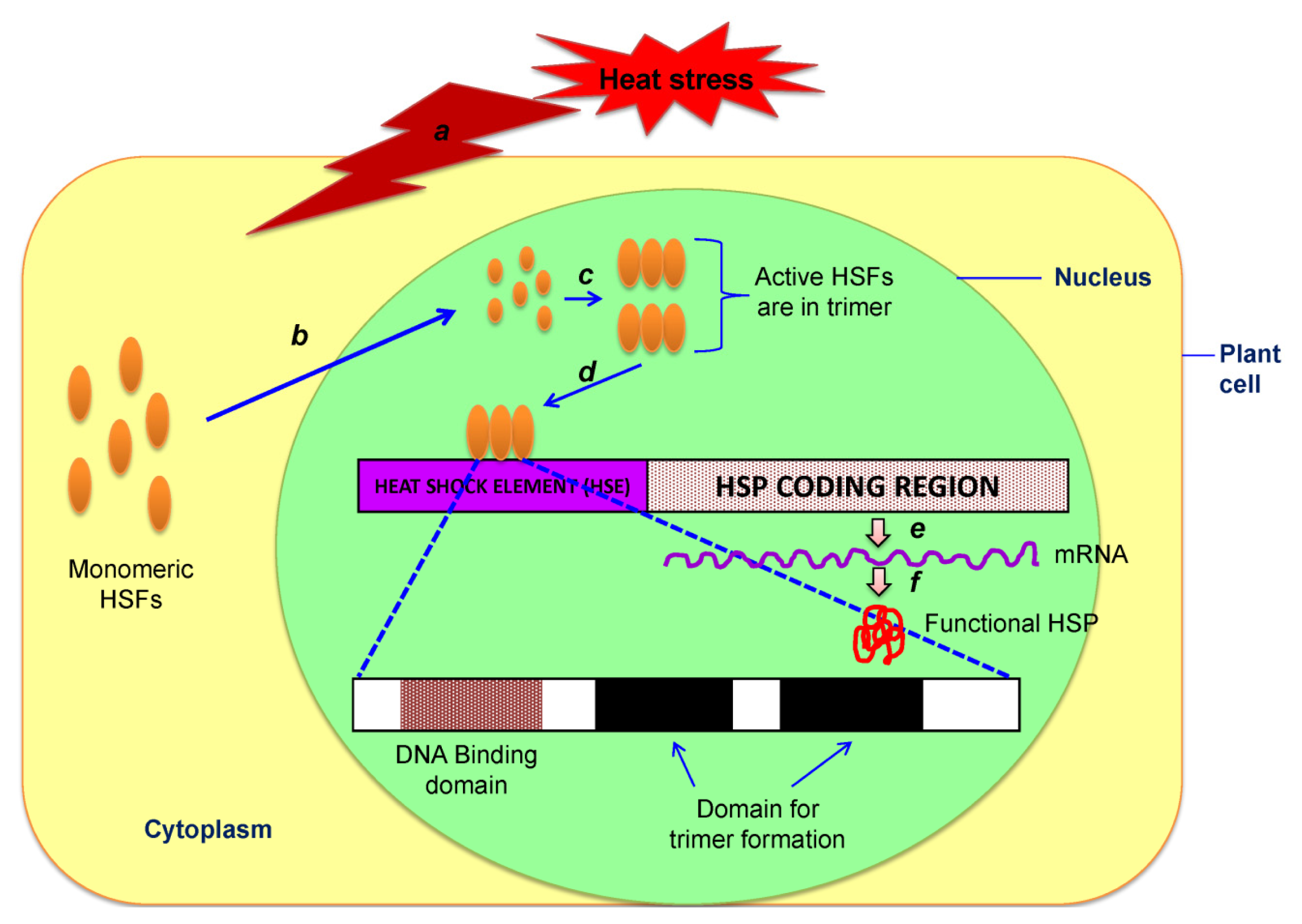
7.1. Heat-Shock Proteins (HSPs): Master Players for Heat Stress Tolerance
7.2. Genetic Engineering and Transgenic Approaches in Conferring Heat Stress Tolerance in Plants
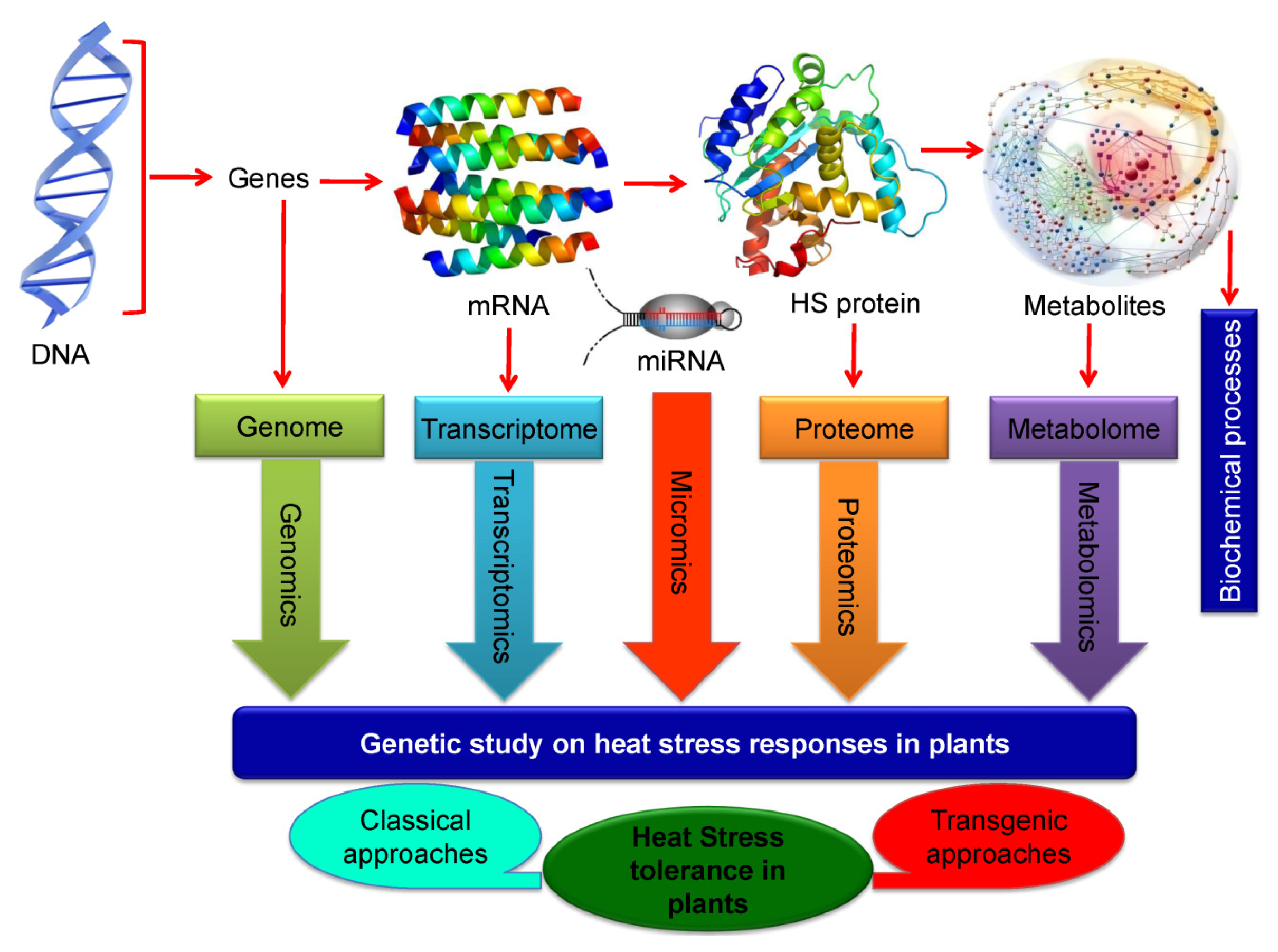
7.3. Omics Approaches in Developing Heat Stress Tolerance
8. Conclusion and Future Perspective
Acknowledgments
Conflict of Interest
References
- Intergovernmental Panel on Climate Change (IPCC), Climate change 2007–The physical science basis. In Contribution of Working Group I to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK, 2007.
- Lobell, D.B.; Asner, G.P. Climate and management contributions to recent trends in U.S. agricultural yields. Science 2003, 299. [Google Scholar] [CrossRef]
- Lobell, D.B.; Field, C.B. Global scale climate–Crop yield relationships and the impacts of recent warming. Environ. Res. Lett. 2007, 2. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Hossain, M.A.; da Silva, J.A.T.; Fujita, M. Plant Responses and Tolerance to Abiotic Oxidative Stress: Antioxidant Defenses is a Key Factor. In Crop Stress and Its Management: Perspectives and Strategies; Bandi, V., Shanker, A.K., Shanker, C., Mandapaka, M., Eds.; Springer: Berlin, Germany, 2012; pp. 261–316. [Google Scholar]
- Hasanuzzaman, M.; Nahar, K.; Fujita, M. Extreme Temperatures, Oxidative Stress and Antioxidant Defense in Plants. In Abiotic Stress—Plant Responses and Applications in Agriculture; Vahdati, K., Leslie, C., Eds.; InTech: Rijeka, Croatia, 2013; pp. 169–205. [Google Scholar]
- Valliyodan, B.; Nguyen, H.T. Understanding regulatory networks and engineering for enhanced drought tolerance in plants. Curr. Opin. Plant Biol 2006, 9, 189–195. [Google Scholar]
- Munns, R.; Tester, M. Mechanisms of salinity tolerance. Ann. Rev. Plant Biol 2008, 59, 651–681. [Google Scholar]
- Janska, A.; Marsik, P.; Zelenkova, S.; Ovesna, J. Cold stress and acclimation: What is important for metabolic adjustment? Plant Biol 2010, 12, 395–405. [Google Scholar]
- Chinnusamy, V.; Zhu, J.; Zhou, T.; Zhu, J.K. Small RNAs: Big Role in Abiotic Stress Tolerance of Plants. In Advances in Molecular Breeding toward Drought and Salt Tolerant Crops; Jenks, M.A., Hasegawa, P.M., Jain, S.M., Eds.; Springer: Dordrecht, The Netherland, 2007; pp. 223–260. [Google Scholar]
- Shinozaki, K.; Yamaguchi-Shinozaki, K. Gene networks involved in drought stress response and tolerance. J. Exp. Bot 2007, 58, 221–227. [Google Scholar]
- Krasensky, J.; Jonak, C. Drought, salt, and temperature stress-induced metabolic rearrangements and regulatory networks. J. Exp. Bot. 2012. [Google Scholar] [CrossRef]
- Semenov, M.A.; Halford, N.G. Identifying target traits and molecular mechanisms for wheat breeding under a changing climate. J. Exp. Bot 2009, 60, 2791–2804. [Google Scholar]
- Moreno, A.A.; Orellana, A. The physiological role of the unfolded protein response in plants. Biol. Res 2011, 44, 75–80. [Google Scholar]
- Hasanuzzaman, M.; Hossain, M.A.; Fujita, M. Physiological and biochemical mechanisms of nitric oxide induced abiotic stress tolerance in plants. Am. J. Plant Physiol 2010, 5, 295–324. [Google Scholar]
- Hasanuzzaman, M.; Hossain, M.A.; Fujita, M. Selenium in higher plants: Physiological role, antioxidant metabolism and abiotic stress tolerance. J. Plant Sci 2010, 5, 354–375. [Google Scholar]
- Hasanuzzaman, M.; Hossain, M.A.; Fujita, M. Exogenous selenium pretreatment protects rapeseed seedlings from cadmium-induced oxidative stress by upregulating the antioxidant defense and methylglyoxal detoxification systems. Biol. Trace Elem. Res 2012, 149, 248–261. [Google Scholar]
- Hasanuzzaman, M.; Nahar, K.; Alam, M.M.; Fujita, M. Exogenous nitric oxide alleviates high temperature induced oxidative stress in wheat (Triticum aestivum) seedlings by modulating the antioxidant defense and glyoxalase system. Aust. J. Crop Sci 2012, 6, 1314–1323. [Google Scholar]
- Hasanuzzaman, M.; Nahar, K.; Fujita, M. Plant response to salt stress and role of exogenous protectants to mitigate salt-induced damages. In Ecophysiology and Responses of Plants under Salt Stress; Ahmad, P., Azooz, M.M., Prasad, M.N.V., Eds.; Springer: New York, NY, USA, 2013; pp. 25–87. [Google Scholar]
- Hasanuzzaman, M.; Gill, S.S.; Fujita, M. Physiological role of nitric oxide in plants grown under adverse environmental conditions. In Plant Acclimation to Environmental Stress; Tuteja, N., Gill, S.S., Eds.; Springer: New York, NY, USA, 2013; pp. 269–322. [Google Scholar]
- Waraich, E.A.; Ahmad, R.; Halim, A.; Aziz, T. Alleviation of temperature stress by nutrient management in crop plants: A review. J. Soil Sci. Plant Nutr 2012, 12, 221–244. [Google Scholar]
- Barnabás, B.; Jäger, K.; Fehér, A. The effect of drought and heat stress on reproductive processes in cereals. Plant Cell Environ 2008, 31, 11–38. [Google Scholar]
- Zhang, Y.; Mian, M.A.R.; Bouton, J.H. Recent molecular and genomic studies on stress tolerance of forage and turf grasses. Crop Sci 2006, 46, 497–511. [Google Scholar]
- Wahid, A.; Gelani, S.; Ashraf, M.; Foolad, M.R. Heat tolerance in plants: An overview. Environ. Exp. Bot 2007, 61, 199–223. [Google Scholar]
- Rodríguez, M.; Canales, E.; Borrás-Hidalgo, O. Molecular aspects of abiotic stress in plants. Biotechnol. Appl 2005, 22, 1–10. [Google Scholar]
- Kosová, K.; Vítámvás, P.; Prášil, I.T.; Renaut, J. Plant proteome changes under abiotic stress-Contribution of proteomics studies to understanding plant stress response. J. Proteom 2011, 74, 1301–1322. [Google Scholar]
- Duque, A.S.; de Almeida, A.M.; da Silva, A.B.; da Silva, J.M.; Farinha, A.P.; Santos, D.; Fevereiro, P.; de Sousa Araújo, S. Abiotic stress responses in plants: Unraveling the complexity of genes and networks to survive. In Abiotic Stress—Plant Responses and Applications in Agriculture; Vahdati, K., Leslie, C., Eds.; InTech: Rijeka, Croatia, 2013; pp. 3–23. [Google Scholar]
- Schöffl, F.; Prandl, R.; Reindl, A. Molecular responses to heat stress. In Molecular Responses to Cold, Drought, Heat and Salt Stress in Higher Plants; Shinozaki, K., Yamaguchi-Shinozaki, K., Eds.; R.G. Landes Co.: Austin, TX, USA, 1999; pp. 81–98. [Google Scholar]
- Ahuja, I.; de Vos, R.C.H.; Bones, A.M.; Hall, R.D. Plant molecular stress responses face climate change. Trends Plant Sci 2010, 15, 664–674. [Google Scholar]
- Mittler, R.; Blumwald, E. Genetic engineering for modern agriculture: Challenges and perspectives. Ann. Rev. Plant Biol 2010, 61, 443–462. [Google Scholar]
- Lobell, D.B.; Schlenker, W.; Costa-Roberts, J. Climate trends and global crop production since 1980. Science 2011, 333, 616–620. [Google Scholar]
- McClung, C.R.; Davis, S.J. Ambient thermometers in plants: From physiological outputs towards mechanisms of thermal sensing. Curr. Biol 2010, 20, 1086–1092. [Google Scholar]
- Ruelland, E.; Zachowski, A. How plants sense temperature. Environ. Exp. Bot 2010, 69, 225–232. [Google Scholar]
- Suzuki, N.; Miller, G.; Morales, J.; Shulaev, V.; Torres, M.A.; Mittler, R. Respiratory burst oxidases: The engines of ROS signaling. Curr. Opin. Plant Biol 2011, 14, 691–699. [Google Scholar]
- Suzuki, N.; Koussevitzky, S.; Mittler, R.; Miller, G. ROS and redox signalling in the response of plants to abiotic stress. Plant Cell Environ 2012, 35, 259–270. [Google Scholar]
- Pagamas, P.; Nawata, E. Sensitive stages of fruit and seed development of chili pepper (Capsicum annuum L. var. Shishito) exposed to high-temperature stress. Sci. Hort 2008, 117, 21–25. [Google Scholar]
- Cao, Y.Y.; Duan, H.; Yang, L.N.; Wang, Z.Q.; Liu, L.J.; Yang, J.C. Effect of high temperature during heading and early filling on grain yield and physiological characteristics in indica rice. Acta Agron. Sin 2009, 35, 512–521. [Google Scholar]
- Hurkman, W.J.; Vensel, W.H.; Tanaka, C.K.; Whitehand, L.; Altenbach, S.B. Effect of high temperature on albumin and globulin accumulation in the endosperm proteome of the developing wheat grain. J. Cereal Sci 2009, 49, 12–23. [Google Scholar]
- Rahman, M.A.; Chikushi, J.; Yoshida, S.; Karim, A.J.M.S. Growth and yield components of wheat genotypes exposed to high temperature stress under control environment. Bangladesh J. Agric. Res 2009, 34, 361–372. [Google Scholar]
- Djanaguiraman, M.; Prasad, P.V.V.; Seppanen, M. Selenium protects sorghum leaves from oxidative damage under high temperature stress by enhancing antioxidant defense system. Plant Physiol. Biochem 2010, 48, 999–1007. [Google Scholar]
- Mohammed, A.R.; Tarpley, L. Effects of high night temperature and spikelet position on yield-related parameters of rice (Oryza sativa L.) plants. Eur. J. Agron 2010, 33, 117–123. [Google Scholar]
- Suwa, R.; Hakata, H.; Hara, H.; El-Shemy, H.A.; Adu-Gyamfi, J.J.; Nguyen, N.T.; Kanai, S.; Lightfoot, D.A.; Mohapatra, P.K.; Fujita, K. High temperature effects on photosynthetic partitioning and sugar metabolism during ear expansion in maize (Zea mays L.) genotypes. Plant Physiol. Biochem 2010, 48, 124–130. [Google Scholar]
- Yin, Y.; Li, S.; Liao, W.; Lu, Q.; Wen, X.; Lu, C. Photosystem II photochemistry, photoinhibition, and the xanthophyll cycle in heat-stressed rice leaves. J. Plant Physiol 2010, 167, 959–966. [Google Scholar]
- Djanaguiraman, M.; Prasad, P.V.V.; Al-Khatib, K. Ethylene perception inhibitor 1-MCP decreases oxidative damage of leaves through enhanced antioxidant defense mechanisms in soybean plants grown under high temperature stress. Environ. Exp. Bot 2011, 71, 215–223. [Google Scholar]
- Tan, W.; Meng, Q.W.; Brestic, M.; Olsovska, K.; Yang, X. Photosynthesis is improved by exogenous calcium in heat-stressed tobacco plants. J. Plant Physiol 2011, 168, 2063–2071. [Google Scholar]
- Gunawardhana, M.D.M.; de Silva, C.S. Impact of temperature and water stress on growth yield and related biochemical parameters of okra. Trop. Agric. Res 2011, 23, 77–83. [Google Scholar]
- Edreira, J.I.R.; Otegui, M.E. Heat stress in temperate and tropical maize hybrids: Differences in crop growth, biomass partitioning and reserves use. Field Crops Res 2012, 130, 87–98. [Google Scholar]
- Zhang, X.; Cai, J.; Wollenweber, B.; Liu, F.; Dai, T.; Cao, W.; Jiang, D. Multiple heat and drought events affect grain yield and accumulations of high molecular weight glutenin subunits and glutenin macropolymers in wheat. J. Cereal Sci 2013, 57, 134–140. [Google Scholar]
- Saitoh, H. Ecological and Physiology of Vegetable; Nousangyoson Bunka Kyoukai: Tokyo, Japan, 2008. [Google Scholar]
- Johkan, M.; Oda, M.; Maruo, T.; Shinohara, Y. Crop production and global warming. In Global Warming Impacts—Case Studies on the Economy, Human Health, and on Urban and Natural Environments; Casalegno, S., Ed.; InTech: Rijeka, Croatia, 2011; pp. 139–152. [Google Scholar]
- Kumar, S.; Kaur, R.; Kaur, N.; Bhandhari, K.; Kaushal, N.; Gupta, K.; Bains, T.S.; Nayyar, H. Heat-stress induced inhibition in growth and chlorosis in mungbean (Phaseolus aureus Roxb.) is partly mitigated by ascorbic acid application and is related to reduction in oxidative stress. Acta Physiol. Plant 2011, 33, 2091–2101. [Google Scholar]
- Piramila, B.H.M.; Prabha, A.L.; Nandagopalan, V.; Stanley, A.L. Effect of heat treatment on germination, seedling growth and some biochemical parameters of dry seeds of black gram. Int. J. Pharm. Phytopharmacol. Res 2012, 1, 194–202. [Google Scholar]
- Toh, S.; Imamura, A.; Watanabe, A.; Okamot, M.; Jikumaru, Y.; Hanada, A.; Aso, Y.; Ishiyama, K.; Tamura, N.; Iuchi, S.; et al. High temperature-induced abscisic acid biosynthesis and its role in the inhibition of gibberellin action in Arabidopsis seeds. Plant Physiol 2008, 146, 1368–1385. [Google Scholar]
- Essemine, J.; Ammar, S.; Bouzid, S. Impact of heat stress on germination and growth in higher plants: Physiological, biochemical and molecular repercussions and mechanisms of defence. J. Biol. Sci 2010, 10, 565–572. [Google Scholar]
- Cheng, L.; Zou, Y.; Ding, S.; Zhang, J.; Yu, X.; Cao, J.; Lu, G. Polyamine accumulation in transgenic tomato enhances the tolerance to high temperature stress. J. Integr. Plant Biol 2009, 51, 489–499. [Google Scholar]
- Mitra, R.; Bhatia, C.R. Bioenergetic cost of heat tolerance in wheat crop. Curr. Sci 2008, 94, 1049–1053. [Google Scholar]
- Ashraf, M.; Hafeez, M. Thermotolerance of pearl millet and maize at early growth stages: Growth and nutrient relations. Biol. Plant 2004, 48, 81–86. [Google Scholar]
- Wahid, A. Physiological implications of metabolites biosynthesis in net assimilation and heat stress tolerance of sugarcane (Saccharum officinarum) sprouts. J. Plant Res 2007, 120, 219–228. [Google Scholar]
- Srivastava, S.; Pathak, A.D.; Gupta, P.S.; Shrivastava, A.K.; Srivastava, A.K. Hydrogen peroxide-scavenging enzymes impart tolerance to high temperature induced oxidative stress in sugarcane. J. Environ. Biol 2012, 33, 657–661. [Google Scholar]
- Omae, H.; Kumar, A.; Shono, M. Adaptation to high temperature and water deficit in the common bean (Phaseolus vulgarisL.) during the reproductive period. J. Bot. 2012. [Google Scholar] [CrossRef]
- Koini, M.A.; Alvey, L.; Allen, T.; Tilley, C.A.; Harberd, N.P.; Whitelam, G.C.; Franklin, K.A. High temperature-mediated adaptations in plant architecture require the bHLH transcription factor PIF4. Curr. Biol 2009, 19, 408–413. [Google Scholar]
- Patel, D.; Franklin, K.A. Temperature-regulation of plant architecture. Plant Signal. Behav 2009, 4, 577–579. [Google Scholar]
- Savin, R.; Stone, P.J.; Nicolas, M.E.; Wardlaw, I.F. Effects of heat stress and moderately high temperature on grain growth and malting quality of barley. Aust. J. Agric. Res 1997, 48, 615–624. [Google Scholar]
- Nahar, K.; Ahamed, K.U.; Fujita, M. Phenological variation and its relation with yield in several wheat (Triticum aestivum L.) cultivars under normal and late sowing mediated heat stress condition. Not. Sci. Biol 2010, 2, 51–56. [Google Scholar]
- Yamamoto, Y.; Aminaka, R.; Yoshioka, M.; Khatoon, M.; Komayama, K.; Takenaka, D.; Yamashita, A.; Nijo, N.; Inagawa, K.; Morita, N.; et al. Quality control of photosystem II: Impact of light and heat stresses. Photosynth. Res 2008, 98, 589–608. [Google Scholar]
- Crafts-Brandner, S.J.; Salvucci, M.E. Sensitivity of photosynthesis in a C4 plant, maize, to heat stress. Plant Physiol 2002, 129, 1773–1780. [Google Scholar]
- Yang, X.; Chen, X.; Ge, Q.; Li, B.; Tong, Y.; Zhang, A.; Li, Z.; Kuang, T.; Lu, C. Tolerance of photosynthesis to photoinhibition, high temperature and drought stress in flag leaves of wheat: A comparison between a hybridization line and its parents grown under field conditions. Plant Sci 2006, 171, 389–397. [Google Scholar]
- Wang, J.Z.; Cui, L.J.; Wang, Y.; Li, J.L. Growth, lipid peroxidation and photosynthesis in two tall fescue cultivars differing in heat tolerance. Biol. Plant 2009, 53, 247–242. [Google Scholar]
- Marchand, F.L.; Mertens, S.; Kockelbergh, F.; Beyens, L.; Nijs, I. Performance of high arctic tundra plants improved during but deteriorated after exposure to a simulated extreme temperature event. Glob. Change Biol 2005, 11, 2078–2089. [Google Scholar]
- Morales, D.; Rodriguez, P.; Dellamico, J.; Nicolas, E.; Torrecillas, A.; Sanchez-Blanco, M.J. High-temperature preconditioning and thermal shock imposition affects water relations, gas exchange and root hydraulic conductivity in tomato. Biol. Plant 2003, 47, 203–208. [Google Scholar]
- Kumar, A.; Omae, H.; Egawa, Y.; Kashiwaba, K.; Shono, M. Some Physiological Responses of Snap Bean (Phseolus vulgarisL.) to Water Stress during Reproductive Period. In Proceedings of the International Conference on Sustainable Crop Production in Stress Environment: Management and Genetic Option; JNKVV: Jabarpur, India, 2005; pp. 226–227. [Google Scholar]
- Greer, D.H.; Weedon, M.M. Modelling photosynthetic responses to temperature of grapevine (Vitis vinifera cv. Semillon) leaves on vines grown in a hot climate. Plant Cell Environ 2012, 35, 1050–1064. [Google Scholar]
- Kepova, K.D.; Holzer, R.; Stoilova, L.S.; Feller, U. Heat stress effects on ribulose-1,5-bisphosphate carboxylase/oxygenase, rubisco binding protein and rubisco activase in wheat leaves. Biol. Plant 2005, 49, 521–525. [Google Scholar]
- Sumesh, K.V.; Sharma-Natu, P.; Ghildiyal, M.C. Starch synthase activity and heat shock protein in relation to thermal tolerance of developing wheat grains. Biol. Plant 2008, 52, 749–753. [Google Scholar]
- Djanaguiraman, M.; Sheeba, J.A.; Devi, D.D.; Bangarusamy, U. Cotton leaf senescence can be delayed by nitrophenolate spray through enhanced antioxidant defense system. J. Agron. Crop Sci 2009, 195, 213–224. [Google Scholar]
- Young, L.W.; Wilen, R.W.; Bonham-Smith, P.C. High temperature stress of Brassica napus during flowering reduces micro- and megagametophyte fertility, induces fruit abortion, and disrupts seed production. J. Exp. Bot 2004, 55, 485–495. [Google Scholar]
- Sato, S.; Kamiyama, M.; Iwata, T.; Makita, N.; Furukawa, H.; Ikeda, H. Moderate increase of mean daily temperature adversely affects fruit set of Lycopersicon esculentum by disrupting specific physiological processes in male reproductive development. Ann. Bot 2006, 97, 731–738. [Google Scholar]
- Maheswari, M.; Yadav, S.K.; Shanker, A.K.; Kumar, M.A.; Venkateswarlu, B. Overview of plant stresses: Mechanisms, adaptations and research pursuit. In Crop Stress and Its Management: Perspectives and Strategies; Venkateswarlu, B., Shanker, A.K., Shanker, C., Maheswari, M., Eds.; Springer: Dordrecht, The Netherlands, 2012; pp. 1–18. [Google Scholar]
- Foolad, M.R. Breeding for abiotic stress tolerances in tomato. In Abiotic Stresses: Plant Resistance through Breeding and Molecular Approaches; Ashraf, M., Harris, P.J.C., Eds.; The Haworth Press Inc: New York, NY, USA, 2005; pp. 613–684. [Google Scholar]
- Cao, Y.Y.; Duan, H.; Yang, L.N.; Wang, Z.Q.; Zhou, S.C.; Yang, J.C. Effect of heat stress during meiosis on grain yield of rice cultivars differing in heat tolerance and its physiological mechanism. Acta Agron. Sin 2008, 34, 2134–2142. [Google Scholar]
- Ahamed, K.U.; Nahar, K.; Fujita, M.; Hasanuzzaman, M. Variation in plant growth, tiller dynamics and yield components of wheat (Triticum aestivum L.) due to high temperature stress. Adv. Agric. Bot 2010, 2, 213–224. [Google Scholar]
- Prasad, P.V.V.; Staggenborg, S.A.; Ristic, Z. Impacts of drought and/or heat stress on physiological, developmental, growth, and yield processes of crop plants. In Response of Crops to Limited Water: Understanding and Modeling Water Stress Effects on Plant Growth Processes; Ahuja, L.H., Saseendran, S.A., Eds.; Advances in Agricultural Systems Modeling Series 1; ASA-CSSA: Madison, WI, USA, 2008; pp. 301–355. [Google Scholar]
- Tubiello, F.N.; Soussana, J.F.; Howden, S.M. Crop and pasture response to climate change. Proc. Nat. Acad. Sci. USA 2007, 104, 19686–19690. [Google Scholar]
- Warland, J.S.; McDonald, M.R.; McKeown, A.M. Annual yields of five crops in the family Brassicacae in southern Ontario in relation to weather and climate. Can. J. Plant Sci 2006, 86, 1209–1215. [Google Scholar]
- Kalra, N.; Chakraborty, D.; Sharma, A.; Rai, H.K.; Jolly, M.; Chander, S.; Kumar, P.R.; Bhadraray, S.; Barman, D.; Mittal, R.B.; et al. Effect of increasing temperature on yield of some winter crops in northwest India. Curr. Sci 2008, 94, 82–88. [Google Scholar]
- Hatfield, J.L.; Boote, K.J.; Kimball, B.A.; Ziska, L.H.; Izaurralde, R.C.; Ort, D.; Thomson, A.; Wolfe, D. Climate impacts on agriculture: Implications for crop production. Agron. J 2011, 103, 351–370. [Google Scholar]
- Wang, X.; Cai, J.; Liu, F.; Jin, M.; Yu, H.; Jiang, D.; Wollenweber, B.; Dai, T.; Cao, W. Pre-anthesis high temperature acclimation alleviates the negative effects of post-anthesis heat stress on stem stored carbohydrates remobilization and grain starch accumulation in wheat. J. Cereal Sci 2012, 55, 331–336. [Google Scholar]
- Kutcher, H.R.; Warland, J.S.; Brandt, S.A. Temperature and precipitation effects on canola yields in Saskatchewan. Can. Agric. Forest Meteorol 2010, 150, 161–165. [Google Scholar]
- Sinsawat, V.; Leipner, J.; Stamp, P.; Fracheboud, Y. Effect of heat stress on the photosynthetic apparatus in maize (Zea mays L.) grown at control or high temperature. Environ. Exp. Bot 2004, 52, 123–129. [Google Scholar]
- Hay, R.K.M.; Porter, J.R. The Physiology of Crop Yield; Blackwell Publishing Ltd: Oxford, UK, 2006. [Google Scholar]
- Reynolds, M.P.; Pierre, C.S.; Saad, A.S.I.; Vargas, M.; Condon, A.G. Evaluating potential genetic gains in wheat associated with stress-adaptive trait expression in elite genetic resources under drought and heat stress. Crop Sci 2007, 47, 172–189. [Google Scholar]
- Cicchino, M.; Edreira, J.I.R; Uribelarrea, M.; Otegui, M.E. Heat stress in field grown maize: response of physiological determinants of grain yield. Crop Sci 2010, 50, 1438–1448. [Google Scholar]
- Högy, P.; Poll, C.; Marhan, S.; Kandeler, E.; Fangmeier, A. Impacts of temperature increase and change in precipitation pattern on crop yield and yield quality of barley. Food Chem 2013, 136, 1470–1477. [Google Scholar]
- Vasseur, F.; Pantin, F.; Vile, D. Changes in light intensity reveal a major role for carbon balance in Arabidopsis responses to high temperature. Plant Cell Environ 2011, 34, 1563–1576. [Google Scholar]
- Asada, K. Production and scavenging of reactive oxygen species in chloroplasts and their functions. Plant Physiol 2006, 141, 391–396. [Google Scholar]
- Soliman, W.S.; Fujimori, M.; Tase, K.; Sugiyama, S.I. Oxidative stress and physiological damage under prolonged heat stress in C3 grass Lolium perenne. Grassland Sci 2011, 57, 101–106. [Google Scholar]
- Halliwell, B. Oxidative stress and neurodegeneration: where are we now? J. Neurochem 2006, 97, 1634–1658. [Google Scholar]
- Moller, I.M.; Jensen, P.E.; Hansson, A. Oxidative modifications to cellular components in plants. Ann. Rev. Plant Biol 2007, 58, 459–481. [Google Scholar]
- Karuppanapandian, T.; Wang, H.W.; Prabakaran, N.; Jeyalakshmi, K.; Kwon, M.; Manoharan, K.; Kim, W. 2,4-dichlorophenoxyacetic acid-induced leaf senescence in mung bean (Vigna radiata L. Wilczek) and senescence inhibition by co-treatment with silver nanoparticles. Plant Physiol. Biochem 2011, 49, 168–177. [Google Scholar]
- Karuppanapandian, T.; Moon, J.C.; Kim, C.; Manoharan, K.; Kim, W. Reactive oxygen species in plants: their generation, signal transduction, and scavenging mechanisms. Aust. J. Crop Sci 2011b, 5, 709–725. [Google Scholar]
- Huang, B.; Xu, C. Identification and characterization of proteins associated with plant tolerance to heat stress. J. Integr. Plant Biol 2008, 50, 1230–1237. [Google Scholar]
- Camejo, D.; Jiménez, A.; Alarcón, J.J.; Torres, W.; Gómez, J.M.; Sevilla, F. Changes in photosynthetic parameters and antioxidant activities following heat–shock treatment in tomato plants. Func. Plant Biol 2006, 33, 177–187. [Google Scholar]
- Bavita, A.; Shashi, B.; Navtej, S.B. Nitric oxide alleviates oxidative damage induced by high temperature stress in wheat. Indian J. Exp. Biol 2012, 50, 372–378. [Google Scholar]
- Savicka, M.; Škute, N. Effects of high temperature on malondialdehyde content, superoxide production and growth changes in wheat seedlings (Triticum aestivum L.). Ekologija 2010, 56, 26–33. [Google Scholar]
- Miller, G.; Schlauch, K.; Tam, R.; Cortes, D.; Torres, M.A.; Shulaev, V.; Dangl, J.L.; Mittler, R. The plant NADPH oxidase RbohD mediates rapid, systemic signaling in response to diverse stimuli. Sci. Signal. 2.
- Qi, Y.; Wang, H.; Zou, Y.; Liu, C.; Liu, Y.; Wang, Y.; Zhang, W. Over-expression of mitochondrial heat shock protein 70 suppresses programmed cell death in rice. FEBS Lett 2010, 585, 231–239. [Google Scholar]
- Źróbek-Sokolnik, A. Temperature stress and responses of plants. In Environmental Adaptations and Stress Tolerance of Plants in the Era of Climate Change; Ahmad, P., Prasad, M.N.V., Eds.; Springer: New York, NY, USA, 2012; pp. 113–134. [Google Scholar]
- Larcher, W. Physiological Plant Ecology; Springer: Berlin, Germany, 1995. [Google Scholar]
- Fitter, A.H.; Hay, R.K.M. Environmental Physiology of Plants, 3rd ed; Academic Press: London, UK, 2002. [Google Scholar]
- Adams, S.R.; Cockshull, K.E.; Cave, C.R.J. Effect of temperature on the growth and development of tomato fruits. Ann. Bot 2001, 88, 869–877. [Google Scholar]
- Sarieva, G.E.; Kenzhebaeva, S.S.; Lichtenthaler, H.K. Adaptation potential of photosynthesis in wheat cultivars with a capability of leaf rolling under high temperature conditions. Russ. J. Plant Physiol 2010, 57, 28–36. [Google Scholar]
- Hall, A.E. The mitigation of heat stress. Available online: http://www.planstress.com (accessed on 3 June 2011).
- Wang, W.; Vinocur, B.; Shoseyov, O.; Altman, A. Role of plant heat-shock proteins and molecular chaperones in the abiotic stress response. Trends Plant Sci 2004, 9, 244–252. [Google Scholar]
- Radin, J.W.; Lu, Z.; Percy, R.G.; Zeiger, E. Genetic variability for stomatal conductance in Pima cotton and its relation to improvements of heat adaptation. Proc. Natl. Acad. Sci. USA 1994, 91, 7217–7221. [Google Scholar]
- Queitsch, C.; Hong, S.W.; Vierling, E.; Lindquist, S. Hsp101 plays a crucial role in thermotolerance in Arabidopsis. Plant Cell 2000, 12, 479–492. [Google Scholar]
- Vinocur, B.; Altman, A. Recent advances in engineering plant tolerance to abiotic stress: achievements and limitations. Curr. Opin. Biotechnol 2005, 16, 123–132. [Google Scholar]
- Almeselmani, M.; Deshmukh, P.S.; Sairam, R.K.; Kushwaha, S.R.; Singh, T.P. Protective role of antioxidant enzymes under high temperature stress. Plant Sci 2006, 171, 382–388. [Google Scholar]
- Babu, N.R.; Devraj, V.R. High temperature and salt stress response in French bean (Phaseolus vulgaris). Aust. J. Crop Sci 2008, 2, 40–48. [Google Scholar]
- Xu, S.; Li, J.; Zhang, X.; Wei, H.; Cui, L. Effects of heat acclimation pretreatment on changes of membrane lipid peroxidation, antioxidant metabolites, and ultrastructure of chloroplasts in two cool-season turfgrass species under heat stress. Environ. Exp. Bot 2006, 56, 274–285. [Google Scholar]
- Gong, M.; Chen, S.N.; Song, Y.Q.; Li, Z.G. Effect of calcium and calmodulin on intrinsic heat tolerance in relation to antioxidant systems in maize seedlings. Aust. J. Plant Physiol 1997, 24, 371–379. [Google Scholar]
- Dat, J.; Foyer, C.; Scott, I. Change in salicylic acid and antioxidants during induced thermo tolerance in mustard seedlings. Plant Physiol 1998, 118, 1455–1461. [Google Scholar]
- Apel, K.; Hirt, H. Reactive oxygen species: metabolism, oxidative stress and signal transduction. Ann. Rev. Plant Biol 2004, 55, 373–399. [Google Scholar]
- Chakraborty, U.; Pradhan, D. High temperature-induced oxidative stress in Lens culinaris, role of antioxidants and amelioration ofstress by chemical pre-treatments. J. Plant Interact 2011, 6, 43–52. [Google Scholar]
- Sairam, R.K.; Srivastava, G.C.; Saxena, D.C. Increased antioxidant activity under elevated temperature: A mechanism of heat stress tolerance in wheat genotypes. Biol. Plant 2000, 43, 245–251. [Google Scholar]
- Kocsy, G.; Szalai, G.; Galiba, G. Effect of heat stress on glutathione biosynthesis in wheat. Acta Biol. Szeged 2002, 46, 71–72. [Google Scholar]
- Chauhan, S. Physiological and Molecular Basis of Heat Tolerance with Emphasis on Oxidative Stress Metabolism in Wheat. PhD Thesis, HNB Garhwal University, Srinagar, Uttaranchal, India, June 2005. [Google Scholar]
- Balla, K.; Bencze, S.; Janda, T.; Veisz, O. Analysis of heat stress tolerance in winter wheat. Acta Agron. Hung 2009, 57, 437–444. [Google Scholar]
- Almeselmani, M.; Deshmukh, P.S.; Sairam, R.K. High temperature stress tolerance in wheat genotypes: role of antioxidant defence enzymes. Acta Agron. Hung 2009, 57, 1–14. [Google Scholar]
- Rani, B.; Dhawan, K.; Jain, V.; Chhabra, M.L.; Singh, D. High temperature induced changes in antioxidative enzymes in Brassica juncea(L) Czern & Coss. Available online: http://www.australianoilseeds.com/__data/assets/pdf_file/0003/6861/46_High_temperature_induced_changes_in_antioxidative_enzymes_in_Brassica_juncea.pdf (accessed on 24 April 2013).
- Kumar, S.; Gupta, D.; Nayyar, H. Comparative response of maize and rice genotypes to heat stress: status of oxidative stress and antioxidants. Acta Physiol. Plant 2012, 34, 75–86. [Google Scholar]
- Tuteja, N. Integrated calcium signaling in plants. In Signaling in Plants I; Baluska, F., Mancuso, S., Eds.; Springer: Heidelberg, Germany, 2009; pp. 29–49. [Google Scholar]
- Kaur, N.; Gupta, A.K. Signal transduction pathways under abiotic stresses in plants. Curr. Sci 2005, 88, 1771–1780. [Google Scholar]
- Ahmad, P.; Bhardwaj, R.; Tuteja, N. Plant signaling under abiotic stress environment. In Environmental Adaptations and Stress Tolerance of Plants in the Era of Climate Change; Ahmad, P., Prasad, M.N.V., Eds.; Springer: New York, NY, USA, 2012; pp. 297–324. [Google Scholar]
- Ciarmiello, L.F.; Woodrow, P.; Fuggi, A.; Pontecorvo, G.; Carillo, P. Plant Genes for Abiotic Stress. In Abiotic Stress in Plants—Mechanisms and Adaptations; Shanker, A.K., Venkateswarlu, B., Eds.; InTech: Rijeka, Croatia, 2011; pp. 283–308. [Google Scholar]
- Proveniers, M.C.G.; van Zanten, M. High temperature acclimation through PIF4 signaling. Trends Plant Sci 2013, 18, 59–64. [Google Scholar]
- Hasanuzzaman, M.; Hossain, M.A.; Fujita, M. Nitric oxide modulates antioxidant defense and the methylglyoxal detoxification system and reduces salinity-induced damage of wheat seedlings. Plant Biotechnol. Rep 2011, 5, 353–365. [Google Scholar]
- Hasanuzzaman, M.; Hossain, M.A.; Fujita, M. Selenium-induced up-regulation of the antioxidant defense and methylglyoxal detoxification system reduces salinity-induced damage in rapeseed seedlings. Biol. Trace Elem. Res 2011, 143, 1704–1721. [Google Scholar]
- Hasanuzzaman, M.; Fujita, M. Selenium and Plants’ Health: The Physiological Role of Selenium. In Selenium: Sources, Functions and Health Effects; Aomori, C., Hokkaido, M., Eds.; Nova Science Publishers: New York, NY, USA, 2012; pp. 101–122. [Google Scholar]
- Rasheed, R.; Wahid, A.; Farooq, M.; Hussain, I.; Basra, S.M.A. Role of proline and glycinebetaine pretreatments in improving heat tolerance of sprouting sugarcane (Saccharum sp.) buds. Plant Growth Regul 2011, 65, 35–45. [Google Scholar]
- Kaushal, N.; Gupta, K.; Bhandhari, K.; Kumar, S.; Thakur, P.; Nayyar, H. Proline induces heat tolerance in chickpea (Cicer arietinum L.) plants by protecting vital enzymes of carbon and antioxidative metabolism. Physiol. Mol. Biol. Plants 2011, 17, 203–213. [Google Scholar]
- Kumar, S.; Kaushal, N.; Nayyar, H.; Gaur, P. Abscisic acid induces heat tolerance in chickpea (Cicer arietinum L.) seedlings by facilitated accumulation of osmoprotectants. Acta Physiol. Plant 2012, 34, 1651–1658. [Google Scholar]
- Lu, J.; Zhang, R.; Zong, X.F.; Wang, S.-G.; He, G.H. Effect of salicylic acid on heat resistance of rice seedling under heat stress. Chin. J. Eco-Agric 2009, 17, 1168–1171. [Google Scholar]
- Wang, L.J.; Fan, L.; Loescher, W.; Duan, W.; Liu, G.J.; Cheng, J.S.; Luo, H.B.; Li, S.H. Salicylic acid alleviates decreases in photosynthesis under heat stress and accelerates recovery in grapevine leaves. BMC Plant Biol 2010, 10, 34. [Google Scholar]
- Chhabra, M.L.; Dhawan, A.; Sangwan, N.; Dhawan, K.; Singh, D. Phtohormones induced amelioration of high temperature stress in Brassica juncea(L.). Proceedings of 16th Australian Research Assembly on Brassicas, Ballarat, Australia, 10–14 September 2009.
- El-Bassiony, A.M.; Ghoname, A.A.; El-Awadi, M.E.; Fawzy, Z.F.; Gruda, N. Ameliorative effects of brassinosteroids on growth and productivity of snap beans grown under high temperature. Gesunde Pflanzen 2012, 64, 175–182. [Google Scholar]
- Kumar, S.; Sirhindi, G.; Bhardwaj, R.; Kumar, M.; Arora, P. Role of 24-epibrassinolide in amelioration of high temperature stress through antioxidant defense system in Brassica juncea L. Plant Stress 2012, 6, 55–58. [Google Scholar]
- Chen, P.; Yu, S.; Zhan, Y.; Kang, X.L. Effects of jasmonate acid on thermotolerance of grape seedlings. J. Shihezi Univ. (Nat. Sci.) 2006, 24, 87–91. [Google Scholar]
- Song, L.; Ding, W.; Zhao, M.; Sun, B.; Zhang, L. Nitric oxide protects against oxidative stress under heat stress in the calluses from two ecotypes of reed. Plant Sci 2006, 171, 449–458. [Google Scholar]
- Yang, J.D.; Yun, J.Y.; Zhang, T.H.; Zhao, H.L. Presoaking with nitric oxide donor SNP alleviates heat shock damages in mung bean leaf discs. Bot. Stud 2006, 47, 129–136. [Google Scholar]
- Khalil, S.I.; El-Bassiouny, H.M.S.; Hassanein, R.A.; Mostafa, H.A. Antioxidant defense system in heat shocked wheat plants previously treated with arginine or putrescine. Aust. J. Basic Appl. Sci 2009, 3, 1517–1526. [Google Scholar]
- Cheng, L.; Sun, R.; Wang, F.; Peng, Z.; Kon, F. Spermidine affects the transcriptome responses to high temperature stress in ripening tomato fruit. J. Zhejiang Univ. Sci. 2012, 283–297. [Google Scholar]
- Bibi, A.C.; Oosterhuis, D.M.; Gonias, E.D. Exogenous application of putrescine ameliorates the effect of high temperature in Gossypium hirsutum L. flowers and fruit development. J. Agron. Crop Sci 2010, 196, 205–211. [Google Scholar]
- Asthir, B.; Koundal, A.; Bains, N.S. Putrescine modulates antioxidant defense response in wheat under high temperature stress. Biol. Plant 2012, 56, 757–761. [Google Scholar]
- Sakamoto, A.; Murata, N. The role of glycine betaine in the protection of plants from stress: Clues from transgenic plants. Plant Cell Environ 2002, 25, 163–171. [Google Scholar]
- Jain, R.; Solomon, S.; Shrivastava, A.K.; Lal, P. Nutrient application improves stubble bud sprouting under low temperature conditions in sugarcane. Sug. Tech 2009, 11, 83–85. [Google Scholar]
- Rasheed, R.; Wahid, A.; Ashraf, M.; Basra, S.M.A. Role of proline and glycinebetaine in improving chilling stress tolerance in sugarcane buds at sprouting. Int. J. Agric. Biol 2010, 12, 1–8. [Google Scholar]
- Wang, L.J.; Li, S.H. Salicylic acid-induced heat or cold tolerance in relation to Ca2+ homeostasis and antioxidant systems in young grape plants. Plant Sci 2006, 170, 685–694. [Google Scholar]
- Ding, W.; Song, L.; Wang, X.; Bi, Y. Effect of abscisic acid on heat stress tolerance in the calli from two ecotypes of Phragmites communis. Biol. Plant 2010, 54, 607–613. [Google Scholar]
- Hossain, K.K.; Itoh, R.D.; Yoshimura, G.; Tokuda, G.; Oku, H.; Cohen, M.F.; Yamasaki, H. Effects of nitric oxide scavengers on thermoinhibition of seed germination in Arabidopsis thaliana. Russ. J. Plant Physiol 2010, 57, 222–232. [Google Scholar]
- Alcázar, R.; Cuevas, J.C.; Patrón, M.; Altabella, T.; Tiburcio, A.F. Abscisic acid modulates polyamine metabolism under water stress in Arabidopsis thaliana. Plant Physiol 2006, 128, 448–455. [Google Scholar]
- Hussain, S.S.; Ali, M.; Ahmad, M.; Siddique, K.H.M. Polyamines: Natural and engineered abiotic and biotic stress tolerance in plants. Biotechnol. Adv 2011, 29, 300–311. [Google Scholar]
- Kusano, T.; Yamaguchi, K.; Berberich, T.; Takahashi, Y. Advances in polyamine research in 2007. J. Plant Res 2007, 120, 345–350. [Google Scholar]
- Mostafa, H.A.M.; Hassanein, R.A.; Khalil, S.I.; El-Khawas, S.A.; El-Bassiouny, H.M.S.; El-Monem, A.A.A. Effect of arginine or putrescine on growth, yield and yield components of late sowing wheat. J. Appl. Sci. Res 2010, 6, 177–183. [Google Scholar]
- Königshofer, H.; Lechner, S. Are polyamines involved in the synthesis of heatshock proteins in cell suspension cultures of tobacco and alfalfa in response to high-temperature stress? Plant Physiol. Biochem 2002, 40, 51–59. [Google Scholar]
- Konstantinos, P.A.; Imene, T.; Panagiotis, M.N.; Roubelakis-Angelakis, K.A. ABA-dependent amine oxidases-derived H2O2 affects stomata conductance. Plant Signal. Behav 2010, 5, 1153–1156. [Google Scholar]
- Chang, H.C.; Tang, Y.C.; Hayer-Hartl, M.; Hartl, F.U. SnapShot: Molecular chaperones, Part I. Cell 2007, 128. [Google Scholar] [CrossRef]
- Baniwal, S.K.; Bharti, K.; Chan, K.Y.; Fauth, M.; Ganguli, A.; Kotak, S.; Mishra, S.K.; Nover, L.; Port, M.; Scharf, K.D.; et al. Heat stress response in plants: a complex game with chaperones and more than twenty heat stress transcription factors. J. Biosci 2004, 29, 471–487. [Google Scholar]
- Prasinos, C.; Krampis, K.; Samakovli, D.; Hatzopoulos, P. Tight regulation of expression of two Arabidopsis cytosolic Hsp90 genes during embryo development. J. Exp. Bot 2005, 56, 633–644. [Google Scholar]
- Swindell, W.R.; Huebner, M.; Weber, A.P. Transcriptional profiling of Arabidopsis heat shock proteins and transcription factors reveals extensive overlap between heat and non-heat stress response pathways. BMC Genom 2007, 8, 125. [Google Scholar]
- Kulz, D. Evolution of the cellular stress proteome: From monophyletic origin to ubiquitous function. J. Exp. Biol 2003, 206, 3119–3124. [Google Scholar]
- Hong, S.W.; Lee, U.; Vierling, E. Arabidopsis hot mutants define multiple functions required for acclimation to high temperatures. Plant Physiol 2003, 132, 757–767. [Google Scholar]
- Morrow, G.; Tanguay, R.M. Small heat shock protein expression and functions during development. Int. J. Biochem. Cell Biol 2012, 44, 1613–1621. [Google Scholar]
- Nover, L.; Bharti, K.; Doring, P.; Mishra, S.K.; Ganguli, A.; Scharf, K.D. Arabidopsis and the heat stress transcription factor world: how many heat stress transcription factors do we need. Cell Stress Chaper 2001, 6, 177–189. [Google Scholar]
- Lee, J.H.; Hubel, A.; Schoffl, F. Derepression of the activity of genetically engineered heat shock factor causes constitutive synthesis of heat shock proteins and increased thermotolerance in transgenic Arabidopsis. Plant J 1995, 8, 603–612. [Google Scholar]
- Mishra, S.K.; Tripp, J.; Winkelhaus, S.; Tschiersch, B.; Theres, K.; Nover, L.; Scharf, K.D. In the complex family of heat stress transcription factors, HsfA1 has a unique role as master regulator of thermotolerance in tomato. Genes Dev 2002, 16, 1555–1567. [Google Scholar]
- Sun, W.; Bernard, C.; van de Cotte, B.; Montagu, M.V.; Verbruggen, N. At-HSP17.6A, encoding a small heat-shock protein in Arabidopsis, can enhance osmotolerance upon overexpression. Plant J 2001, 27, 407–415. [Google Scholar]
- Wang, Y.; Ying, J.; Kuzma, M.; Chalifoux, M.; Sample, A.; McArthur, C.; Uchacz, T.; Sarvas, C.; Wan, J.; Dennis, D.T.; et al. Molecular tailoring of farnesylation for plant drought tolerance and yield protection. Plant J 2005, 43, 413–424. [Google Scholar]
- Miroshnichenko, S.; Tripp, J.; Nieden, U.Z.; Neumann, D.; Conrad, U.; Manteuffel, R. Immunomodulation of function of small heat shock proteins prevents their assembly into heat stress granules and results in cell death at sub-lethal temperatures. Plant J 2005, 41, 269–281. [Google Scholar]
- Torok, Z.; Goloubinoff, P.; Horvath, I.; Tsvetkova, N.M.; Glatz, A.; Balogh, G.; Varvasovszki, V.; Los, D.A.; Vierling, E.; et al. Synechocystis HSP17 is an amphitropic protein that stabilizes heat-stressed membranes and binds denatured proteins for subsequent chaperone mediated refolding. Proc. Natl. Acad. Sci. USA 2001, 98, 3098–3103. [Google Scholar]
- Reis, S.P.; Lima, A.M.; de Souza, C.R.B. Recent molecular advances on downstream plant responses to abiotic stress. Int. J. Mol Sci 2012, 13, 8628–8647. [Google Scholar]
- Cho, E.K.; Hong, C.B. Over-expression of tobacco NtHSP70–1 contributes to drought-stress tolerance in plants. Plant Cell Rep 2006, 25, 349–358. [Google Scholar]
- Liming, Y.; Qian, Y.; Pigang, L.; Sen, L. Expression of the HSP24 gene from Trichoderma harzianum in Saccharomyces cerevisiae. J. Therm. Biol 2008, 33, 1–6. [Google Scholar]
- Bohnert, H.J.; Gong, Q.; Li, P.; Ma, S. Unraveling abiotic stress tolerance mechanisms—Getting genomics going. Curr. Opin. Plant Biol 2006, 9, 180–188. [Google Scholar]
- Maestri, E.; Klueva, N.; Perrotta, C.; Gulli, M.; Nguyen, H.T.; Marmiroli, N. Molecular genetics of heat tolerance and heat shock proteins in cereals. Plant Mol. Biol 2002, 48, 667–681. [Google Scholar]
- Malik, M.K.; Slovin, J.P.; Hwang, C.H.; Zimmerman, J.L. Modified expression of a carrot small heat shock protein gene, Hsp17.7, results in increased or decreased thermotolerance. Plant J 1999, 20, 89–99. [Google Scholar]
- Liu, J.; Shono, M. Characterization of mitochondria-located small heat shock protein from tomato (Lycopersicon esculentum). Plant Cell Physiol 1999, 40, 1297–1304. [Google Scholar]
- Sanmiya, K.; Suzuki, K.; Egawa, Y.; Shono, M. Mitochondrial small heat shock protein enhances thermotolerance in tobacco plants. FEBS Lett 2004, 557, 265–268. [Google Scholar]
- Katiyar-Agarwal, S.; Agarwal, M.; Grover, A. Heat tolerant basmati rice engineered by over-expression of hsp101. Plant Mol. Biol 2003, 51, 677–686. [Google Scholar]
- Lee, B.H.; Won, S.H.; Lee, H.S.; Miyao, M.; Chung, W.I.; Kim, I.J.; Jo, J. Expression of the chloroplast-localized small heat shock protein by oxidative stress in rice. Gene 2000, 245, 283–290. [Google Scholar]
- Murakami, T.; Matsuba, S.; Funatsuki, H.; Kawaguchi, K.; Saruyama, H.; Tanida, M.; Sato, Y. Overexpression of a small heat shock protein, sHSP17.7, confers both heat tolerance and UV-B resistance to rice plants. Mol. Breed 2004, 13, 165–175. [Google Scholar]
- Ono, K.; Hibino, T.; Kohinata, T.; Suzuki, S.; Tanaka, Y.; Nakamura, T.; Takabe, T.; Takabe, T. Overexpression of DnaK from a halotolerant cyanobacterium Aphanothece halophytica enhances the high-temperature tolerance of tobacco during germination and early growth. Plant Sci 2001, 160, 455–461. [Google Scholar]
- Alia, H.H.; Sakamoto, A.; Murata, N. Enhancement of the tolerance of Arabidopsis to high temperatures by genetic engineering of the synthesis of glycinebetaine. Plant J 1998, 16, 155–161. [Google Scholar]
- Yang, X.; Liang, Z.; Lu, C. Genetic engineering of the biosynthesis of glycine betaine enhances photosynthesis against high temperature stress in transgenic tobacco plants. Plant Physiol 2005, 138, 2299–2309. [Google Scholar]
- Salvucci, M.E.; Crafts-Brandner, S.J. Inhibition of photosynthesis by heat stress: the activation state of Rubisco as a limiting factor in photosynthesis. Physiol. Plant 2004, 120, 179–186. [Google Scholar]
- Sharkey, T.D.; Badger, M.R.; Von-Caemmerer, S.; Andrews, T.J. Increased heat sensitivity of photosynthesis in tobacco plants with reduced Rubisco activase. Photosyn. Res 2001, 67, 147–156. [Google Scholar]
- Murakami, Y.; Tsuyama, M.; Kobayashi, Y.; Kodama, H.; Iba, K. Trienoic fatty acids and plant tolerance of high temperature. Science 2000, 287, 476–479. [Google Scholar]
- Kovtun, Y.; Chiu, W.L.; Tena, G.; Sheen, J. Functional analysis of oxidative stress-activated mitogen-activated protein kinase cascade in plants. Proc. Natl. Acad. Sci. USA 2000, 97, 2940–2945. [Google Scholar]
- Rizhsky, R.; Liang, H.; Shuman, J.; Shulaev, V.; Davletova, S.; Mittler, R. When defense pathways collide. The response of Arabidopsis to a combination of drought and heat stress. Plant Physiol 2004, 134, 1683–1696. [Google Scholar]
- Shi, W.M.; Muramoto, Y.; Ueda, A.; Takabe, T. Cloning of peroxisomal ascorbate peroxidase gene from barley and enhanced thermotolerance by overexpressing in Arabidopsis thaliana. Gene 2001, 273, 23–27. [Google Scholar]
- Davison, P.A.; Hunter, C.N.; Horton, P. Overexpression of beta-carotene hydroxylase enhances stress tolerance in Arabidopsis. Nature 2002, 418, 203–206. [Google Scholar]
- Grover, A.; Mittal, D.; Negi, M.; Lavania, D. Generating high temperature tolerant transgenic plants: Achievements and challenges. Plant Sci 2013, 205–206, 38–47. [Google Scholar]
- Montero-Barrientos, M.; Hermosa, R.; Cardoza, R.E.; Gutiérrez, S.; Nicolás, C.; Monte, E. Transgenic expression of the Trichoderma harzianum hsp70 gene increases Arabidopsis resistance to heat and other abiotic stresses. J. Plant Physiol 2010, 167, 659–665. [Google Scholar]
- Sohn, S.O.; Back, K. Transgenic rice tolerant to high temperature with elevated contents of dienoic fatty acids. Biol. Plant 2007, 51, 340–342. [Google Scholar]
- Park, S.M.; Hong, C.B. Class I small heat shock protein gives thermotolerance in tobacco. J. Plant Physiol 2002, 159, 25–30. [Google Scholar]
- Aprile, A.; Mastrangelo, A.M.; De Leonardis, A.M.; Galiba, G.; Roncaglia, E.; Ferrari, F.; De Bellis, L.; Turchi, L.; Giuliano, G.; Cattivelli, L. Transcriptional profiling in response to terminal drought stress reveals differential responses along the wheat genome. BMC Genom 2009, 10, 279. [Google Scholar]
- Hasanuzzaman, M.; Nahar, K.; Fujita, M.; Ahmad, P.; Chandna, R.; Prasad, M.N.V.; Öztürk, M. Enhancing Plant Productivity under Salt Stress—Relevance of Poly-Omics. In Salt Stress in Plants: Omics, Signaling and Responses; Ahmad, P., Azooz, M.M., Prasad, M.N.V., Eds.; Springer: Berlin, Germany, 2013; pp. 113–156. [Google Scholar]
- Yeh, C.H.; Kaplinsky, N.J.; Hu, C.; Charng, Y.Y. Some like it hot, some like it warm: Phenotyping to explore thermotolerance diversity. Plant Sci 2012, 195, 10–23. [Google Scholar]
- Wienkoop, S.; Morgenthal, K.; Wolschin, F.; Scholz, M.; Selbig, J.; Weckwerth, W. Integration of metabolomic and proteomic phenotypes: analysis of data covariance dissects starch and RFO metabolism from low and high temperature compensation response in Arabidopsis thaliana. Mol. Cell Proteom 2008, 7, 1725–1736. [Google Scholar]
- Caldana, C.; Degenkolbe, T.; Cuadros-Inostroza, A.; Klie, S.; Sulpice, R.; Leisse, A.; Steinhauser, D.; Fernie, A.R.; Willmitzer, L.; Hannah, M.A. High-density kinetic analysis of the metabolomic and transcriptomic response of Arabidopsis to eight environmental conditions. Plant J 2011, 67, 869–884. [Google Scholar]
- De Block, M.; Verduyn, C.; De Brouwer, D.; Cornelissen, M. Poly (ADP-ribose) polymerase in plants affects energy homeostasis, cell death and stress tolerance. Plant J 2005, 41, 95–106. [Google Scholar]
- Maruyama, K.; Takeda, M.; Kidokoro, S.; Yamada, K.; Sakuma, Y.; Urano, K.; Fujita, M.; Yoshiwara, K.; Matsukura, S.; Morishita, Y.; et al. Metabolic pathways involved in cold acclimation identified by integrated analysis of metabolites and transcripts regulated by DREB1A and DREB2A. Plant Physiol 2009, 150, 1972–1980. [Google Scholar]
- Hirai, M.Y.; Klein, M.; Fujikawa, Y.; Yano, M.; Goodenowe, D.B.; Yamazaki, Y.; Kanaya, S.; Nakamura, Y.; Kitayama, M.; Suzuki, H.; et al. Elucidation of gene-to-gene and metabolite-to-gene networks in Arabidopsis by integration of metabolomics and transcriptomics. J. Biol. Chem 2005, 280, 25590–25595. [Google Scholar]
- Deuschle, K.; Okumoto, S.; Fehr, M.; Looger, L.L.; Kozhukh, L.; Frommer, W.B. Construction and optimization of a family of genetically encoded metabolite sensors by semirational protein engineering. Protein Sci 2005, 14, 2304–2314. [Google Scholar]
- Liu, H.C.; Liao, H.Y.; Charng, Y.Y. The role of class A1 heat shock factors (HSFA1s) in response to heat and other stresses in Arabidopsis. Plant Cell Environ 2011, 34, 738–751. [Google Scholar]
- Rizhsky, L.; Hongjian, L.; Mittler, R. The combined effect of drought stress and heat shock on gene expression in tobacco. Plant Physiol 2002, 130, 1143–1151. [Google Scholar]
- Penueli, L.; Liang, H.; Rozenberg, M.; Mittler, R. Growth suppression, altered stomatal responses, and augmented induction of heat shock proteins in cytosolic ascorbate peroxidase (Apx1)-deficient Arabidopsis plants. Plant J 2003, 34, 187–203. [Google Scholar]
- Meehl, G.A.; Stocker, T.F.; Collins, W.D.; Friedlingstein, P.; Gaye, A.T.; Gregory, J.M.; Kitoh, A.; Knutti, R.; Murphy, J.M.; Noda, A.; et al. Global Climate Projections. In Climate Change 2007; Solomon, S., Qin, D., Manning, M., Chen, Z., Marquis, M., Averyt, K.B., Tignor, M., Miller, H.L., Eds.; Cambridge University Press: Cambridge, UK, 2007; pp. 749–845. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Crops | Heat treatment | Growth stage | Major effects | References |
|---|---|---|---|---|
| Chili pepper (Capsicum annuum) | 38/30 °C (day/night) | Reproductive, maturity and harvesting stage | Reduced fruit width and fruit weight, increased the proportion of abnormal seeds per fruit. | [36] |
| Rice (Oryza sativa) | Above 33 °C, 10 days | Heading stage | Reduced the rates of pollen and spikelet fertility. | [37] |
| Wheat (Triticum aestivum) | 37/28 °C (day/night), 20 days | Grain filling and maturity stage | Shortened duration of grain filling and maturity, decreases in kernel weight and yield. | [38] |
| Wheat (Triticum aestivum) | 30/25 °C day/night | From 60 DAS to maturity stage | Reduced leaf size, shortened period for days to booting, heading, anthesis, and maturity, drastic reduction of number of grains/spike and smaller grain size and reduced yield. | [39] |
| Sorghum (Hordeum vulgare) | 40/30 °C (day/night) | 65 DAS to maturity stage | Decreased chlorophyll (chl) content, chl a fluorescence, decreased photosystem II (PSII) photochemistry, Pn and antioxidant enzyme activity and increased ROS content, and thylakoid membrane damage, reduced yield. | [40] |
| Rice (Oryza sativa) | 32 °C (night temperature) | Reproductive stage | Decreased yield, increased spikelet sterility, decreased grain length, width and weight. | [41] |
| Maize (Zea mays) | 35/27 °C (day/night), 14 days | Reproductive stage | Reduced ear expansion, particularly suppression of cob extensibility by impairing hemicellulose and cellulose synthesis through reduction of photosynthate supply. | [42] |
| Rice (Oryza sativa) | 25–42.5 °C | Vegetative growth stage | Decrease in the CO2 assimilation rate. | [43] |
| Soybean (Glycine max) | 38/28 °C (day/night), 14 days | Flowering stage | Decreased the leaf Pn and stomatal conductance (gs), increased thicknesses of the palisade and spongy layers, damaged plasma membrane, chloroplast membrane, and thylakoid membranes, distorted mitochondrial membranes, cristae and matrix. | [44] |
| Tobacco (Nicotiana tabacum) | 43 °C, 2 h | Early growth stage | Decrease in net photosynthetic rate (Pn), stomatal conductance as well as the apparent quantum yield (AQY) and carboxylation efficiency (CE) of photosynthesis. Reduced the activities of antioxidant enzymes. | [45] |
| Okra (Abelmoschus esculentus) | 32 and 34 °C | Throughout the growing period | Reduced yield, damages in pod quality parameters such as fibre content and break down of the Ca-pectate. | [46] |
| Maize (Zea mays) | 33–40 °C, 15 days | During Pre-anthesis and silking onwards | Severe effect on plant and ear growth rates. | [47] |
| Wheat (Triticum aestivum) | 38 °C, 24 and 48 h | Seedling stage | Decreased chl and relative water content (RWC); diminished antioxidative capacity. | [18] |
| Wheat (Triticum aestivum) | 32/24 °C (day/night), 24 h | At the end of spikelet initiation stage | Spikelet sterility, reduced grain yield. | [48] |
| Crops | Heat treatments | Protectants | Protective effects | References |
|---|---|---|---|---|
| Saccharum officinarum | 42 °C, 48 h | 20 mM Pro or GB, 8 h | Restricted the H2O2 generation, improved K+ and Ca2+ contents, and increased the concentrations of free Pro | [138] |
| Cicer arietinum | 45/40 °C, 10 days | 10 μM Pro, 10 days | Reduced membrane injury Improved water and chl content Enhanced activities of antioxidants Reduced oxidative stress Enhance activities of enzymes of carbon metabolism | [139] |
| Cicer arietinum | 35/30, 40/35 and 45/40 °C as day/night | 10 μM Pro, GB and Tre | Increased growth Less oxidative damages Decreased MDA and H2O2 contents | [140] |
| Oryza sativa | 35 °C, 48 h | 0.5 mM SA, 24 h | Decreased electrolyte osmosis Reduced MDA content and O2·− production rate | [141] |
| Vitis vinifera | 43 °C, 24 h | 100 μM SA, 24 h | Higher Rubisco activity Increased PSII function Increased photosynthesis | [142] |
| Brassica juncea | 47 ± 5 °C | 0.5 & 1 μM ABA, 4 h | Decreased seedling mortality Increased growth | [143] |
| Cicer arietinum | 35/30, 40/35 and 45/40 °C as day/night | 2.5 μM ABA | Increased growth Less oxidative damages Decreased MDA and H2O2 contents | [140] |
| Phaseolus vulgaris | 34.7 to 35.2 °C | 25, 50 mg L−1 BRs spray | Increased vegetative growth, total yield and quality of pods Increased the total phenolic acids in the pod | [144] |
| Brassica juncea | 40 °C, 5 h × 3 days | 1 μM 24-EBL, 8 h | Better growth Increased protein content Enhanced antioxidant defense | [145] |
| Brassica juncea | 47 ± 5 °C | 100 μM IAA, 4 h | Decreased seedling mortality Increased growth | [143] |
| Brassica juncea | 47 ± 5 °C | 100 μM GA, 4 h | Decreased seedling mortality Increased growth | [143] |
| Vitis vinifera | 42 °C, 12 & 18 h | 50 μM JA, 6 h | Upregulation of the activities of antioxidant enzymes | [146] |
| Brassica juncea | 47 ± 5 °C | 50 and 100 μM kinetin | Decreased seedling mortality Increased growth | [143] |
| Phragmites communis | 45 °C, 2 h | 100 μM SNP and SNAP, 24 h | Decreased H2O2 and MDA contents. Increased activities of SOD, CAT, APX and POD | [147] |
| Phaseolus radiatus | 45 °C, 90 min | 150 μM SNP, 60 min | Increased the activities of CAT, SOD and POD | [148] |
| Triticum aestivum | 35 ± 2 °C, 4 or 8 h | Arginine or Put (0.0, 1.25 and 2.5 mM), 4 or 8 h | Increased SOD and CAT activities, increased DNA and RNA contents, reduced MDA level | [149] |
| Solanum lycopersicum | 33/27 °C, 16/8 h (light/dark) | Spd, 1 mM as pretreatment | Increase in the expression of Eth-related genes, PA biosynthesis genes, hormone pathways genes, and oxidation reduction genes | [150] |
| Gossypium hirsutum | 38 °C up to flowering stage | 10 mM Put, 24 h prior to anthesis | Increased endogenous Put content and seeds/cotton boll | [151] |
| Triticum aestivum | 45 °C in germinated seeds, 2 h | Put, 10 μM | Elevated activities of enzymatic and non-enzymatic antioxidants and DAO and PAO, reduced lipid peroxides in root and shoot | [152] |
| Sorghum bicolor | 40/30 °C, 45 days | 75 mg L−1 Na2SeO4 foliar spray | Decreased membrane damage Enhanced antioxidant defense Increased grain yield | [40] |
| Major classes of heat shock protein | Functions |
|---|---|
| HSP100 | ATP-dependent dissociation and degradation of aggregate protein |
| HSP90 | Co-regulator of heat stress linked signal transduction complexes and manages protein folding. It requires ATP for its function |
| HSP70, HSP40 | Primary stabilization of newly formed proteins, ATP-dependent binding and release |
| HSP60, HSP10 | ATP-dependent specialized folding machinery |
| HSP20 or small HSP (sHSP) | Formation of high molecular weight oligomeric complexes which serve as cellular matrix for stabilization of unfolded proteins. HSP100, HSP70 and HSP40 are needed for its release |
| Transgenic plants | Transgenes | Function of transgenes | References |
|---|---|---|---|
| Z. mays and O. sativa | Hsp100, Hsp101 from A. thaliana | HSP synthesis for HT tolerance | [114,187] |
| A. thaliana | Hsp70 | HSP synthesis for thermotolerance | [201] |
| N. tabacum | Fad 7 from N. tabacum and O. sativa | Desaturation of fatty acids (trienoic fatty acids and hexa-decatrienoic acid) that increased the level of unsaturated fatty acids and provide HT tolerance | [195,202] |
| Daucus carota | Hsp17.7 from D. carota | Synthesis of sHsp | [184,189] |
| N. tabacum | TLHS1 | Synthesis of sHSP (Class I) | [203] |
| A. thaliana | AtHSF1 | Heat shock transcription factor HSF1::GUS (β-glucuronidase) fusion and such modification will increase HSP production in large scale with small investment of HSFs | [173] |
| A. thaliana | gusA | β-glucuronidase synthesis and bind with HSFs to form active trimer | [173] |
| N. tabacum | MT-sHSP from L. esculentum | Molecular chaperone function in vitro | [185,186] |
| N. tabacum | Dnak1 from Aphanothece halophytica | High temperature tolerance | [190] |
| N. tabacum | BADH (betain aldehyde dehydrogenase) from Spinacia oleracea | Over production of GB osmolyte that will enhance the heat tolerance | [192] |
| A. thaliana | Cod A (choline oxidase A) from A. globiformis | Glycine betaine systhesis for tolerance to HT during imbibition and seedling germination | [191] |
| N. tabacum | ANP1/NPK1 | H2O2 responsive MAPK kinase kinase (MAPKKK) production to protect against the lethality in HT | [196] |
| A. thaliana | Ascorbate peroxidase (APX1 from P. sativum and HvAPX1 from H. vulgare) | H2O2 detoxification and conferred heat tolerance | [198] |
© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Hasanuzzaman, M.; Nahar, K.; Alam, M.M.; Roychowdhury, R.; Fujita, M. Physiological, Biochemical, and Molecular Mechanisms of Heat Stress Tolerance in Plants. Int. J. Mol. Sci. 2013, 14, 9643-9684. https://doi.org/10.3390/ijms14059643
Hasanuzzaman M, Nahar K, Alam MM, Roychowdhury R, Fujita M. Physiological, Biochemical, and Molecular Mechanisms of Heat Stress Tolerance in Plants. International Journal of Molecular Sciences. 2013; 14(5):9643-9684. https://doi.org/10.3390/ijms14059643
Chicago/Turabian StyleHasanuzzaman, Mirza, Kamrun Nahar, Md. Mahabub Alam, Rajib Roychowdhury, and Masayuki Fujita. 2013. "Physiological, Biochemical, and Molecular Mechanisms of Heat Stress Tolerance in Plants" International Journal of Molecular Sciences 14, no. 5: 9643-9684. https://doi.org/10.3390/ijms14059643
APA StyleHasanuzzaman, M., Nahar, K., Alam, M. M., Roychowdhury, R., & Fujita, M. (2013). Physiological, Biochemical, and Molecular Mechanisms of Heat Stress Tolerance in Plants. International Journal of Molecular Sciences, 14(5), 9643-9684. https://doi.org/10.3390/ijms14059643