Anti-Inflammatory Effect of Ginsenoside Rg5 in Lipopolysaccharide-Stimulated BV2 Microglial Cells


Abstract
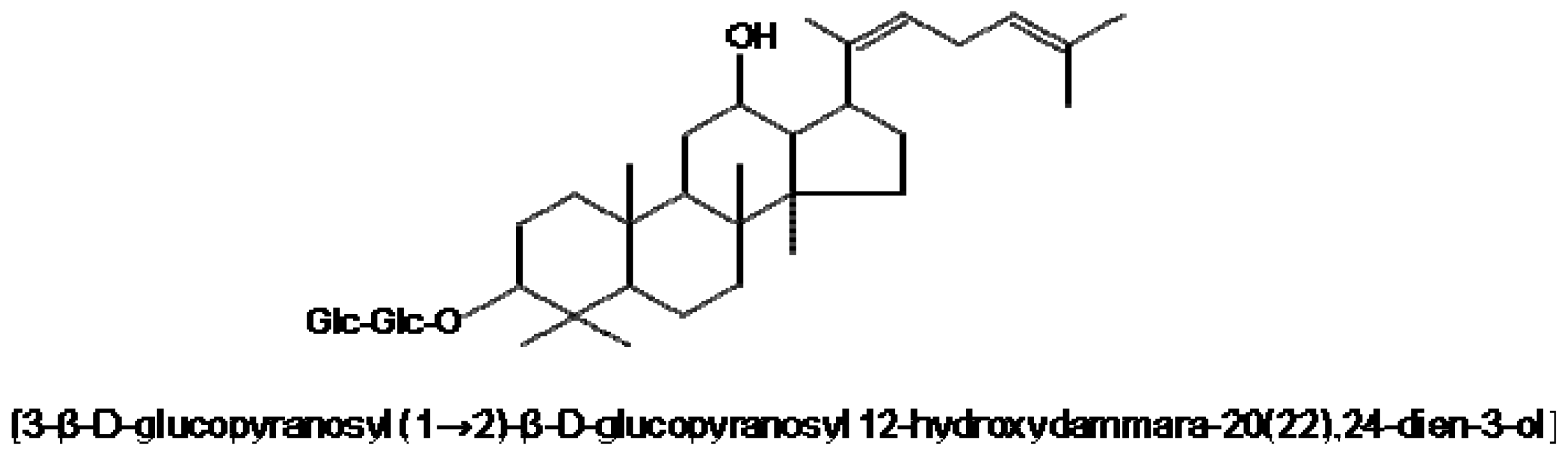
:1. Introduction
2. Results
2.1. Ginsenoside Rg5 Inhibited LPS-Induced NO Production and iNOS Expression in BV2 Microglial Cells
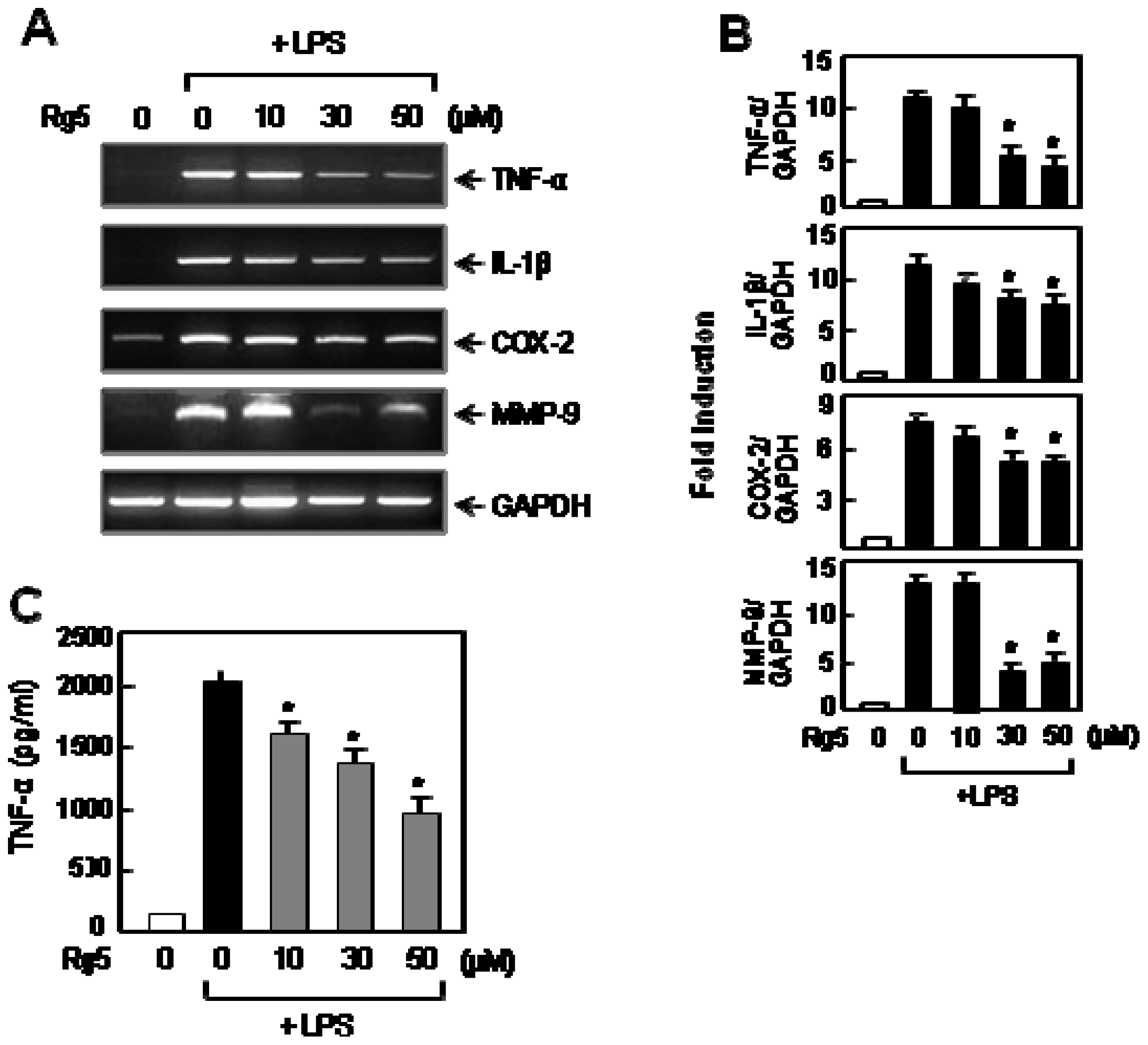
2.2. Rg5 Inhibited LPS-Induced Expression of TNF-α and Other Pro-Inflammatory Molecules
2.3. Rg5 Inhibited LPS-Induced NO and TNF-α Production in Rat Primary Microglia
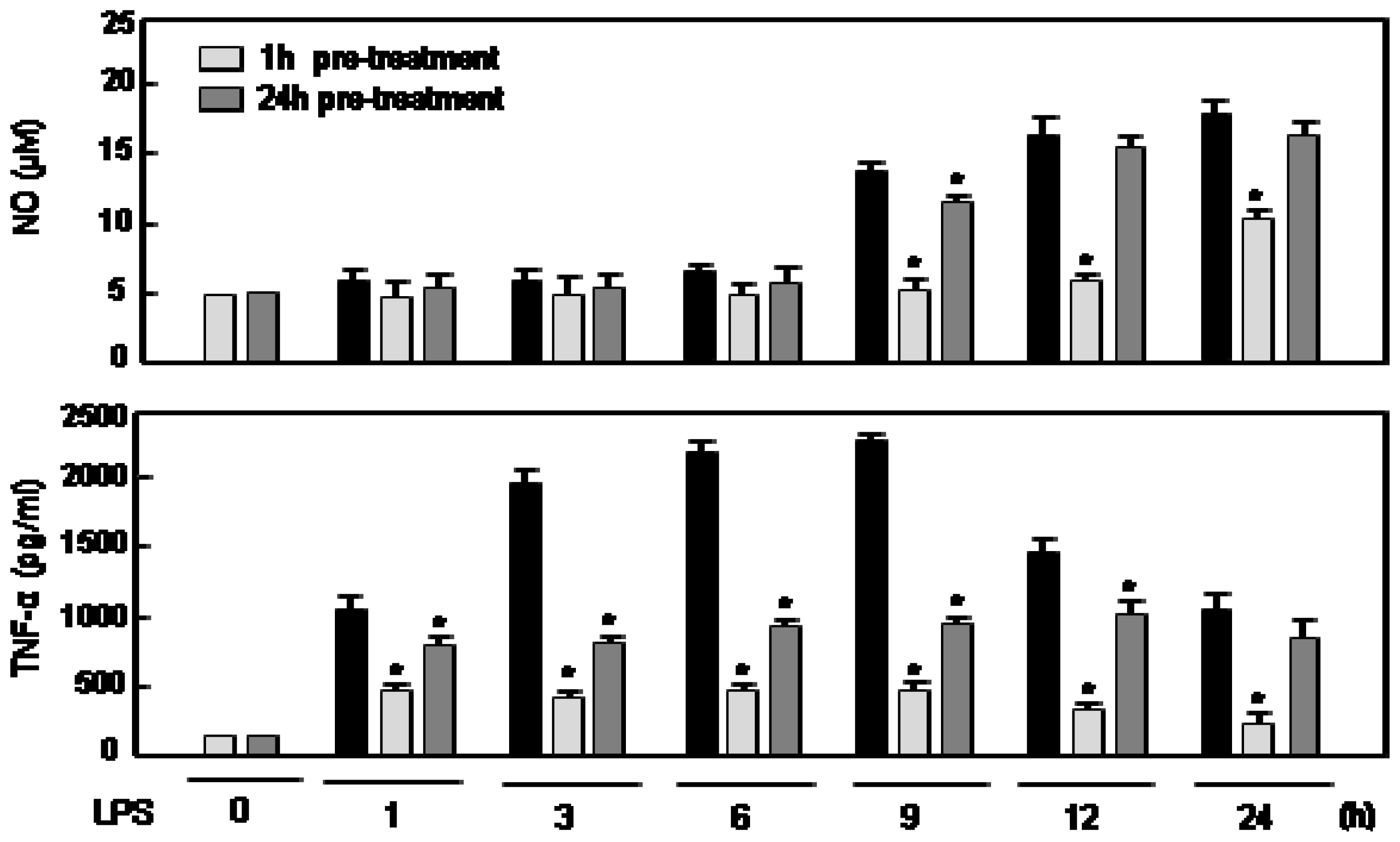
2.4. The Effects of Rg5 on NO and TNF Production Are Dependent on Pre-Treatment Time of Rg5
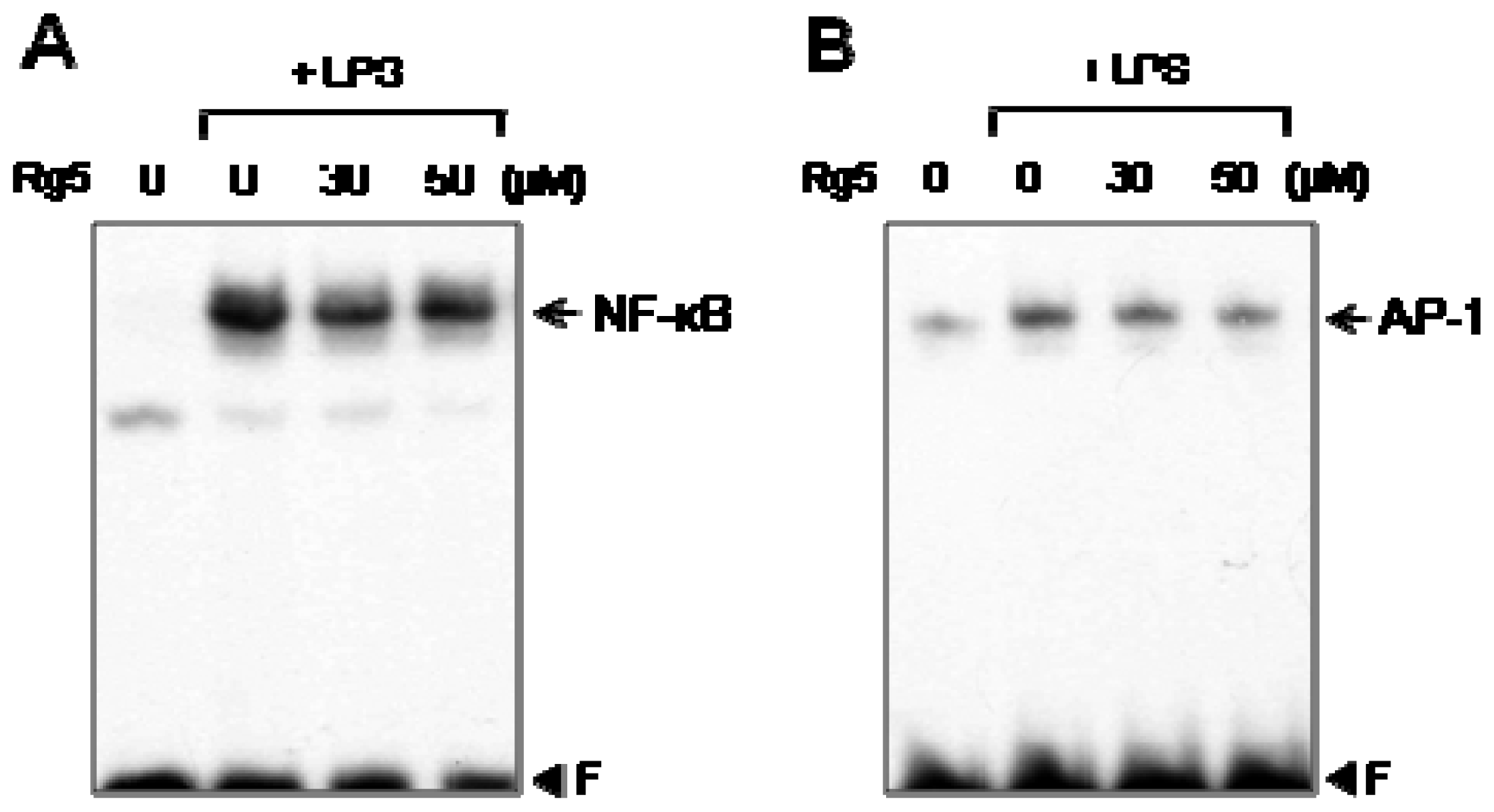
2.5. Rg5 Suppressed LPS-Induced DNA Binding Activities of NF-κB and AP-1
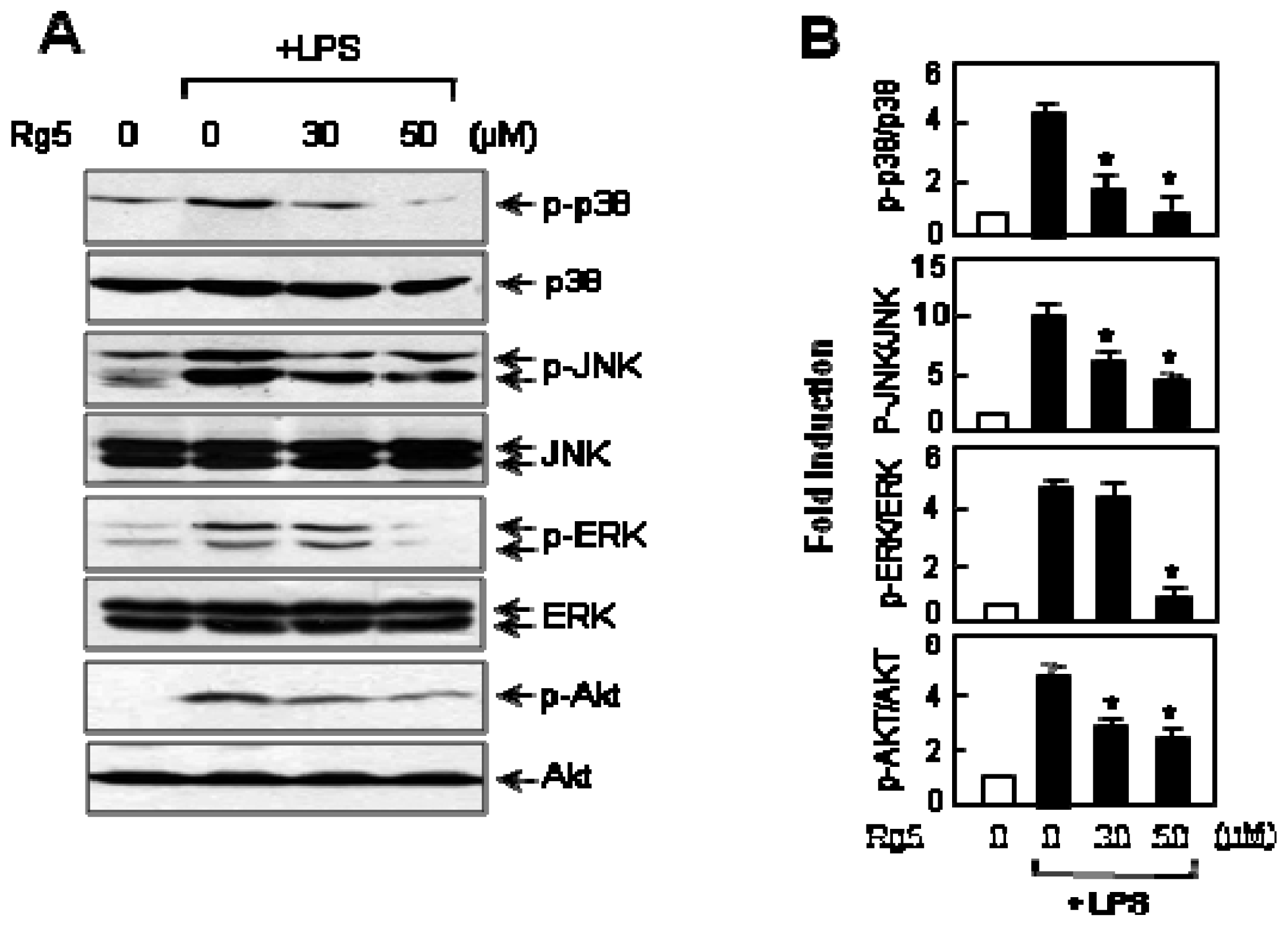
2.6. Rg5 Inhibited LPS-Induced Phosphorylation of MAPKs and Akt in BV2 Cells
2.7. Rg5 Inhibited ROS Production with Increase of ARE-Mediated of HO-1 Expression
3. Discussion
4. Experimental Section
4.1. Reagents
4.2. Microglial Cell Culture
4.3. Measurement of TNF-α, Nitrite and Intracellular ROS Levels
4.4. RT-PCR
4.5. Electrophoretic Mobility Shift Assay (EMSA)
4.6. Western Blot Analysis
4.7. Statistical Analysis
5. Conclusions
Acknowledgments
Conflict of Interest
References
- Tremblay, M.È.; Stevens, B.; Sierra, A.; Wake, H.; Bessis, A.; Nimmerjahn, A. The role of microglia in the healthy brain. J. Neurosci 2011, 31, 16064–16069. [Google Scholar]
- Block, M.L.; Hong, J.S. Microglia and inflammation-mediated neurodegeneration: Multiple triggers with a common mechanism. Progr. Neurobiol 2005, 76, 77–98. [Google Scholar]
- Kim, S.U.; de Vellis, J. Microglia in health and disease. J. Neurosci. Res 2005, 81, 302–313. [Google Scholar]
- Block, M.L.; Zecca, L.; Hong, J.S. Microglia-mediated neurotoxicity: Uncovering the molecular mechanisms. Nat. Rev. Neurosci 2007, 8, 57–69. [Google Scholar]
- Wyss-Coray, T.; Mucke, L. Inflammation in neurodegenerative disease: A double-edged sword. Neuron 2002, 35, 419–432. [Google Scholar]
- Chow, J.C.; Young, D.W.; Golenbock, D.T.; Christ, W.J.; Gusovsky, F. Toll-like receptor-4 mediates lipopolysaccharide-induced signal transduction. J. Biol. Chem 1999, 274, 10689–10692. [Google Scholar]
- Oh, Y.T.; Lee, J.Y.; Lee, J.; Kim, H.; Yoon, K.S.; Choe, W.; Kang, I. Oleic acid reduces lipopolysaccharide-induced expression of iNOS and COX-2 in BV2 murine microglial cells: Possible involvement of reactive oxygen species, p38 MAPK, and IKK/NF-κB signaling pathways. Neurosci. Lett 2009, 464, 93–97. [Google Scholar]
- Skaper, S.D. The brain as a target for inflammatory processes and neuroprotective strategies. Ann. N. Y. Acad. Sci 2007, 1122, 23–24. [Google Scholar]
- Jana, M.; Jana, A.; Liu, X.; Ghosh, S.; Pahan, K. Involvement of phosphatidylinositol 3-kinase-mediated up-regulation of IκBα in anti-inflammatory effect of gemfibrozil in microglia. J. Immunol 2007, 179, 4142–4152. [Google Scholar]
- Syapin, P.J. Regulation of Heme Oxygenase-1 for treatment of neuroinflammation and brain disorders. Br. J. Pharmacol 2008, 115, 623–640. [Google Scholar]
- Keum, Y.S. Regulation of Nrf2-mediated phase II detoxification and anti-oxidant genes. Biomol. Ther 2012, 20, 144–151. [Google Scholar]
- Martin, D.; Rojo, A.I.; Salinas, M.; Diaz, R.; Gallardo, G.; Alam, J.; De Galarreta, C.M.; Cuadrado, A. Regulation of hemeoxygenase-1 expression through the phosphatidylinositol 3-kinase/Akt pathway and the Nrf2 transcription factor in response to the antioxidant phytochemical carnosol. J. Biol. Chem 2004, 279, 8919–8929. [Google Scholar]
- Kim, S.N.; Ha, Y.W.; Shine, H.; Son, S.H.; Wu, S.J.; Kim, Y.S. Simultaneous quantification of 12 ginsenosides in Panax ginseng C.A. Meyer (Korean red ginseng) by HPLC-ELSD and its application to quality control. J. Pharm. Biomed. Anal 2007, 45, 164–170. [Google Scholar]
- Yun, T.K.; Lee, Y.S.; Lee, Y.H.; Kim, S.I.; Yun, H.Y. Anticarcinogenic effect of Panax ginseng C.A. Meyer and identification of active compounds. J. Korean Med. Sci 2001, 16, S6–S18. [Google Scholar]
- Blumenthal, M. Asian ginseng: Potential therapeutic uses. Adv. Nurse Pract 2001, 2, 26–28. [Google Scholar]
- Choi, I.H. Effects of Panax ginseng in neurodegenerative diseases. J. Ginseng Res 2012, 36, 342–353. [Google Scholar]
- Lee, K.Y.; Lee, Y.H.; Kim, S.I.; Park, J.H.; Lee, S.K. Ginsenoside-Rg5 suppresses cyclin E-dependent protein kinase activity via upregulating p21Cip/WAF1 and down-regulating cyclin E in SK-HEP-1 cells. Anticancer Res 1997, 17, 1067–1072. [Google Scholar]
- Kang, K.S.; Kim, H.Y.; Baek, S.H.; Yoo, H.H.; Park, J.H.; Yokozawa, T. Study on the hydroxyl radical scavenging activity changes of ginseng and ginsenoside-Rb2 by heating processing. Biol. Pharm. Bull 2007, 30, 724–728. [Google Scholar]
- Shin, Y.W.; Bae, E.A.; Kim, D.H. Inhibitory effect of ginsenoside Rg5 and its metabolite ginsenoside Rh3 in an oxazolone-induced mouse chronic dermatitis model. Arch. Pharm. Res 2006, 29, 685–690. [Google Scholar]
- Kim, T.W.; Joh, E.H.; Kim, B.; Kim, D.H. Ginsenoside Rg5 ameliorates lung inflammation in mice by inhibiting the binding of LPS to toll-like teceptor-4 on macrophages. Int. Immunopharmacol 2012, 12, 110–116. [Google Scholar]
- Kim, E.J.; Jung, I.H.; Lee, T.K.V.; Jeong, J.J.; Kim, N.J.; Kim, D.H. Ginsenoside Rg5 and Rh3 protect scopolamine-induced memory deficits in mice. J. Ethnopharmacol 2013, 146, 294–299. [Google Scholar]
- Smale, S.T. Selective transcription in response to an inflammatory stimulus. Cell 2010, 140, 833–844. [Google Scholar]
- Munoz, L.; Amit, A.J. Targeting p38 MAPK pathway for the treatment of Alzheimer’s disease. Neuropharmacology 2010, 58, 561–568. [Google Scholar]
- Sayre, L.M.; Perry, G.; Smith, M.A. Oxidative stress and neurotoxicity. Chem. Res. Toxicol 2008, 2, 172–188. [Google Scholar]
- Lee, J.Y.; Jhun, B.S.; Oh, Y.T.; Lee, J.H.; Choe, W.; Baik, H.H.; Ha, J.; Yoon, K.S.; Kim, S.S.; Kang, I. Activation of adenosine A3 receptor suppresses lipopolysaccharide-induced TNF-α production through inhibition of PI3-kinase/Akt and NF-κB activation in murine BV2 microglial cells. Neurosci. Lett 2006, 396, 1–6. [Google Scholar]
- Smolinska, M.J.; Page, T.H.; Urbaniak, A.M.; Mutch, B.E.; Horwood, N.J. Hck tyrosine kinase regulates TLR4-induced TNF and IL-6 production via AP-1. J. Immunol 2011, 187, 6043–6051. [Google Scholar]
- Woo, M.S.; Park, J.S.; Choi, I.Y.; Kim, W.K.; Kim, H.S. Inhibition of MMP-3 or -9 suppresses lipopolysaccharide-induced expression of pro-inflammatory cytokines and iNOS in microglia. J. Neurochem 2008, 106, 770–780. [Google Scholar]
- Lin, C.C.; Hsieh, H.L.; Shih, R.H.; Chi, P.L.; Cheng, S.E.; Chen, J.C.; Yang, C.M. NADPH oxidase 2-derived reactive oxygen species signal contributes to bradykinin-induced matrix metalloproteinase-9 expression and cell migration in brain astrocytes. Cell Commun. Signal 2012, 10, 35. [Google Scholar]
- Jung, J.S.; Shin, J.A.; Park, E.M.; Lee, J.E.; Kang, Y.S.; Min, S.W.; Kim, D.H.; Hyun, J.W.; Shin, C.Y.; Kim, H.S. Anti-inflammatory mechanism of ginsenoside Rh1 in lipopolysaccharide-stimmulated microglia: Critical role of the protein kinase A and hemeoxygenase-1 expression. J. Neurochem 2010, 115, 1668–1680. [Google Scholar]
- Attele, A.S.; Wu, J.A.; Yuan, C.S. Ginseng pharmacology: Multiple constituents and multiple actions. Biochem. Pharmacol 1999, 58, 1685–1693. [Google Scholar]
- Wu, J.; Pan, Z.; Wang, Z.; Zhu, W.; Shen, Y.; Cui, R.; Lin, J.; Yu, H.; Wang, Q.; Qian, J.; et al. Ginsenoside Rg1 protection against β-amyloid peptide-induced neuronal apoptosis via estrogen receptor α and glucocorticoid receptor-dependent anti-protein nitration pathway. Neuropharmacology 2012, 63, 349–361. [Google Scholar]
- Yang, C.S.; Ko, S.R.; Cho, B.S.; Shin, D.M.; Yuk, J.M.; Li, S.; Kim, J.M.; Evans, R.M.; Jung, J.S.; Song, D.K.; et al. The ginsenoside metabolite compound K, a novel agonist of glucocorticoid receptor, induces tolerance to endotoxin-induced lethal shock. J. Cell. Mol. Med 2008, 12, 1739–1753. [Google Scholar]
- Wu, J.; Jeong, H.K.; Bulin, S.E.; Kwon, S.W.; Park, J.H.; Bezprozvanny, I. Ginsenosides protect striatal neurons in cellular model of Huntington’s disease. J. Neurosci. Res 2009, 87, 1904–1912. [Google Scholar]
- Park, J.S.; Shin, J.A.; Jung, J.S.; Hyun, J.W.; Le, T.K.V.; Kim, D.H.; Park, E.M.; Kim, H.S. Anti-inflammatory mechanism of compound K in activated microglia and its neuroprotective effect on experimental stroke in mice. J. Pharmacol. Exp. Ther 2012, 341, 59–67. [Google Scholar]
- Kwon, S.W.; Han, S.B.; Park, I.H.; Kim, J.M.; Park, M.K.; Park, J.H. Liquid chromatographic determination of less polar ginsenosides in processed ginseng. J. Chromatogr. A 2001, 921, 335–339. [Google Scholar]
- Bocchini, V.; Mazzolla, R.; Barluzzi, R.; Blasi, E.; Sick, P.; Kettenmann, H. An immortalized cell line expresses properties of activated microglial cells. J. Neurosci. Res 1992, 31, 616–621. [Google Scholar]
- Qin, L.; Li, G.; Qian, X.; Liu, Y.; Wu, X.; Liu, B.; Hong, J.S.; Block, M.L. Interactive role of the toll-like receptor 4 and reactive oxygen species in LPS-induced microglia activation. Glia 2005, 52, 78–84. [Google Scholar]
- Woo, M.S.; Jang, P.G.; Park, J.S.; Kim, W.K.; Joh, T.H.; Kim, H.S. Selective modulation of lipoopolysaccharide-stimulated cytokine expression and mitogen-activated protein kinase pathways by dibutyryl-cAMP in BV2 microglial cells. Mol. Brain Res 2003, 113, 86–96. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer (5′→3′) | Reverse Primer (5′ →3′) | Size |
|---|---|---|---|
| iNOS | CAAGAGTTTGACCAGAGGACC | TGGAACCACTCGTACTTGGGA | 450 bp |
| TNF-α | CCTATGTCTCAGCCTCTTCT | CCTGGTATGAGATAGCAAAT | 354 bp |
| IL-1β | GGCAACTGTTCCTGAACTCAACTG | CCATTGAGGTGGAGAGCTTTCAGC | 447 bp |
| COX-2 | TTCAAAAGAAGTGCTGGAAAAGGT | GATCATCTCTACCTGAGTGTCTTT | 304 bp |
| MMP-9 | GTGATCCCCACTTACTATGGAAAC | GAAGCCATACAGTTTATCCTGGTC | 352 bp |
| GAPDH | ATGTACGTAGCCATCCAGGC | AGGAAGGAAGGCTGGAAGAG | 420 bp |
© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Lee, Y.Y.; Park, J.-S.; Jung, J.-S.; Kim, D.-H.; Kim, H.-S. Anti-Inflammatory Effect of Ginsenoside Rg5 in Lipopolysaccharide-Stimulated BV2 Microglial Cells. Int. J. Mol. Sci. 2013, 14, 9820-9833. https://doi.org/10.3390/ijms14059820
Lee YY, Park J-S, Jung J-S, Kim D-H, Kim H-S. Anti-Inflammatory Effect of Ginsenoside Rg5 in Lipopolysaccharide-Stimulated BV2 Microglial Cells. International Journal of Molecular Sciences. 2013; 14(5):9820-9833. https://doi.org/10.3390/ijms14059820
Chicago/Turabian StyleLee, Yu Young, Jin-Sun Park, Ji-Sun Jung, Dong-Hyun Kim, and Hee-Sun Kim. 2013. "Anti-Inflammatory Effect of Ginsenoside Rg5 in Lipopolysaccharide-Stimulated BV2 Microglial Cells" International Journal of Molecular Sciences 14, no. 5: 9820-9833. https://doi.org/10.3390/ijms14059820
APA StyleLee, Y. Y., Park, J. -S., Jung, J. -S., Kim, D. -H., & Kim, H. -S. (2013). Anti-Inflammatory Effect of Ginsenoside Rg5 in Lipopolysaccharide-Stimulated BV2 Microglial Cells. International Journal of Molecular Sciences, 14(5), 9820-9833. https://doi.org/10.3390/ijms14059820