(Healthy) Ageing: Focus on Iodothyronines




Abstract
:1. The Thyroid Gland and Thyroid Hormones: A Brief Introduction
2. Thyroid Gland Activity: Ageing vs. Healthy Ageing
2.1. Selective Age-Related Decline in Thyroid Hormone Levels, Evidence from Animal Studies
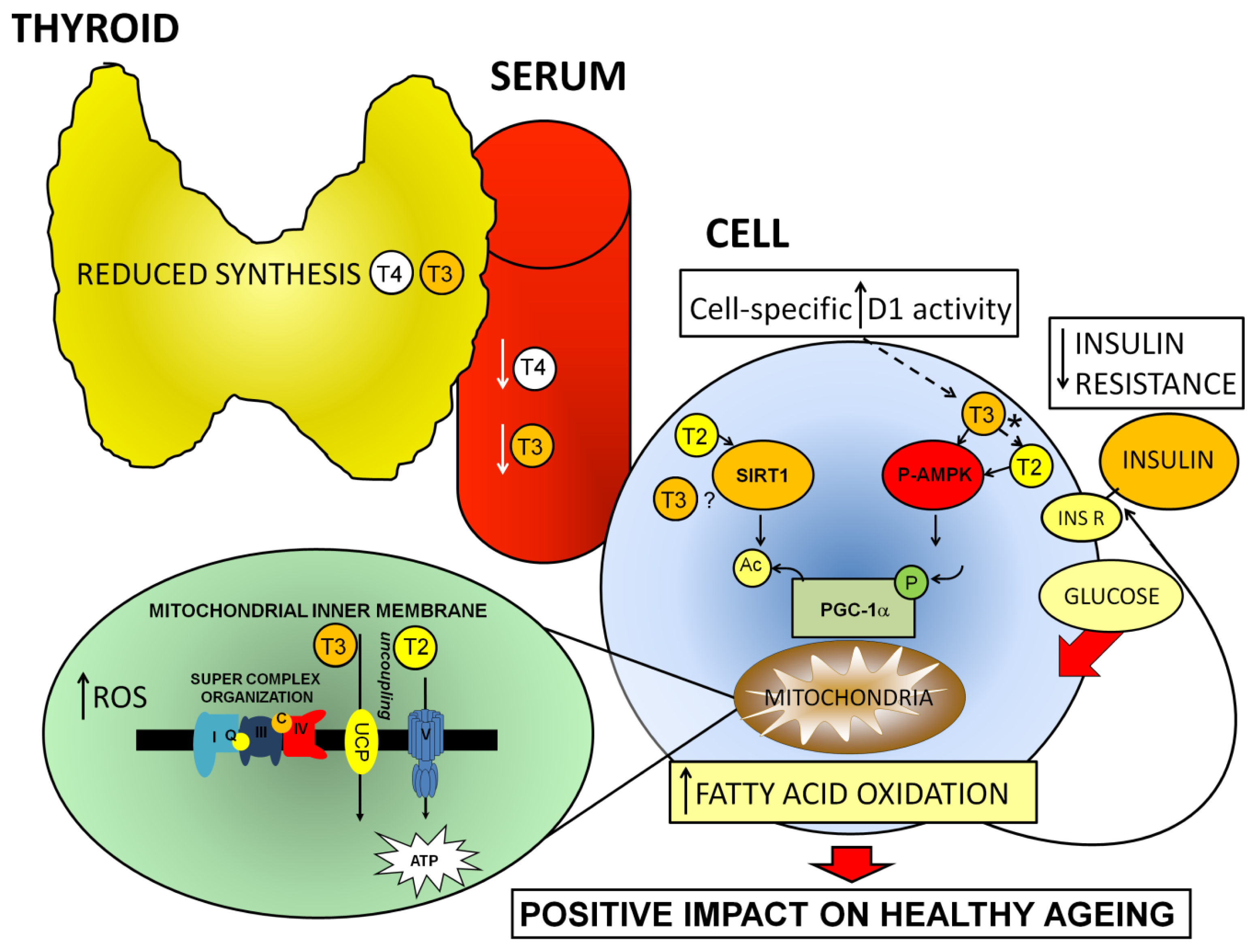
2.2. Toward an Understanding of the Age-Related Control of Peripheral T3 Levels
2.3. Thyroid Hormone Metabolism in Humans, and Maintenance of Type I Deiodinase Activity during Ageing
3. Healthy Ageing: Key Factors Involved, and Their Relationship with the Thyroid and Iodothyronines
3.1. Central Factors in Metabolic Control: Sirtuins and AMPK
3.2. Resveratrol: Providing a Link among Ageing, AMPK, SIRT1, and Iodothyronine Function
3.3. Oxidative Stress and Ageing
3.4. Glycemic Control and Ageing
3.5. Evidence for Contrasting Effects of T3 and T2 on SIRT1 Activity
4. Mitochondrial Organization during Ageing: Proteomic and Transgenic Approaches
5. Conclusions
Acknowledgments
Conflict of Interest
References
- Yen, P.M. Physiological and molecular basis of thyroid hormone action. Physiol. Rev 2001, 81, 1079–1142. [Google Scholar]
- Braverman, L.E.; Ingbar, S.H.; Sterling, K. Conversion of thyroxine (T4) to triiodothyronine (T3) in athyreotic human subjects. J. Clin. Invest 1970, 49, 855–864. [Google Scholar]
- Koehrle, J. The selenoenzyme family of deiodinase isozymes controls local thyroid hormone availability. Rev. Endocr. Metab. Dis 2000, 1, 49–58. [Google Scholar]
- Silvestri, E.; Lombardi, A.; de Lange, P.; Schiavo, L.; Lanni, A.; Goglia, F.; Visser, T.J.; Moreno, M. Age-related changes in renal and hepatic cellular mechanisms associated with variations in rat serum thyroid hormone levels. Am. J. Physiol. Endocrinol. Metab 2008, 294, E1160–E1168. [Google Scholar]
- Engler, D.; Burger, A.G. The deiodination of the iodothyronines and of their derivatives in man. Endocr. Rev 1984, 5, 151–184. [Google Scholar]
- Dentice, M.; Salvatore, D. Deiodinases: The balance of thyroid hormone: Local impact of thyroid hormone inactivation. J. Endocrinol 2011, 209, 273–282. [Google Scholar]
- Weinberger, C.; Thompson, C.C.; Onges, E.S.; Lebo, R.; Gruol, D.J.; Evans, R.M. The c-erbA gene encodes a thyroid hormone receptor. Nature 1986, 324, 641–646. [Google Scholar]
- Sap, J.; Munoz, A.; Damm, K.; Goldberg, Y.; Ghyadael, J.; Leutz, A.; Beug, H.; Vennstrom, B. The c-erbA protein is a high affinity receptor for thyroid hormone. Nature 1986, 324, 635–640. [Google Scholar]
- Lazar, M.A. Thyroid hormone receptors: Multiple forms, multiple possibilities. Endocr. Rev 1993, 14, 348–399. [Google Scholar]
- Hodin, R.A.; Lazar, M.A.; Chin, W.W. Differential and tissue-specific regulation of the multiple rat c-erbA mRNA species by thyroid hormone. J. Clin. Invest 1990, 85, 101–104. [Google Scholar]
- Strait, K.A.; Schwartz, H.L.; Perez-Castillo, A.; Oppenheimer, J.H. Relationship of c-erbA mRNA content to tissue triiodothyronine nuclear binding capacity and function in developing and adult rats. J. Biol. Chem. 1990, 265, 10514–10521. [Google Scholar]
- Moreno, M.; de Lange, P.; Lombardi, A.; Silvestri, E.; Lanni, A.; Goglia, F. Metabolic effects of thyroid hormone derivatives. Thyroid 2008, 18, 239–253. [Google Scholar]
- Davis, P.J.; Davis, F.B. Nongenomic actions of thyroid hormone. Thyroid 1996, 6, 497–504. [Google Scholar]
- Moeller, L.C.; Co, X.; Dumitrescu, A.M.; Seo, H.; Refetoff, S. Thyroid hormone mediated changes in gene expression can be initiated by cytosolic action of the thyroid hormone receptor β through the phosphatidylinositol 3-kinase pathway. Nucl. Recept. Signal 2006, 4, 1–4. [Google Scholar]
- Lanni, A.; Moreno, M.; Lombardi, A.; de Lange, P.; Silvestri, E.; Ragni, M.; Farina, P.; Baccari, G.C.; Fallahi, P.; Antonelli, A.; et al. 3,5-diiodo-l-thyronine powerfully reduces adiposity in rats by increasing the burning of fats. FASEB J 2005, 19, 1552–1554. [Google Scholar]
- Irrcher, I.; Walkinshaw, D.R.; Sheehan, T.E.; Hood, D.A. Thyroid hormone (T3) rapidly activates p38 and AMPK in skeletal muscle in vivo. J. Appl. Physiol 2008, 104, 178–185. [Google Scholar]
- De Lange, P.; Senese, R.; Cioffi, F.; Moreno, M.; Lombardi, A.; Silvestri, E.; Goglia, F.; Lanni, A. Rapid activation by 3,5,3′-l-triiodothyronine of adenosine 5′-monophosphate-activated protein kinase/acetyl-coenzyme a carboxylase and Akt/protein kinase B signaling pathways: Relation to changes in fuel metabolism and myosin heavy-chain protein content in rat gastrocnemius muscle in vivo. Endocrinology 2008, 149, 6462–6470. [Google Scholar]
- Lombardi, A.; de Lange, P.; Silvestri, E.; Busiello, R.A.; Lanni, A.; Goglia, F.; Moreno, M. 3,5-diiodo-l-thyronine rapidly enhances mitochondrial fatty acid oxidation rate and thermogenesis in rat skeletal muscle: AMP-activated protein kinase involvement. Am. J. Physiol. Endocrinol. Metab 2009, 296, E497–E502. [Google Scholar]
- Lanni, A.; Moreno, M.; Cioffi, M.; Goglia, F. Effect of 3,3′-diiodothyronine and 3,5-diiodothyronine on rat liver oxidative capacity. Mol. Cell. Endocrinol 1992, 86, 143–148. [Google Scholar]
- Lanni, A.; Moreno, M.; Lombardi, A.; Goglia, F. Rapid stimulation in vitro of rat liver cytochrome oxidase activity by 3,5-diiodo-l-thyronine and by 3,3′-diiodo-l-thyronine. Mol. Cell. Endocrinol 1994, 99, 89–94. [Google Scholar]
- De Lange, P.; Cioffi, F.; Senese, R.; Moreno, M.; Lombardi, A.; Silvestri, E.; de Matteis, R.; Lionetti, L.; Mollica, M.P.; Goglia, F.; et al. Nonthyrotoxic prevention of diet-induced insulin resistance by 3,5-diiodo-l-thyronine in rats. Diabetes 2011, 60, 2730–2739. [Google Scholar]
- Cordeiro, A.; Souza, L.L.; Oliveira, L.S.; Faustino, L.C.; Santiago, L.A.; Bloise, F.; Ortiga-Carvalho, T.M.; Almeida, N.A.; Pazos-Moura, C.C. Thyroid hormone regulation of sirtuin 1 (SIRT1) expression and implications to integrated responses in fasted mice. J. Endocrinol 2012, 216, 181–193. [Google Scholar]
- Cioffi, F.; Lanni, A.; Goglia, F. Thyroid hormones, mitochondrial bioenergetics and lipid handling. Curr. Opin. Endocrinol. Diabetes Obes 2010, 17, 402–407. [Google Scholar]
- Cordeiro, A.; Souza, L.L.; Einicker-Lamas, M.; Pazos-Moura, C.C. Non-classic thyroid hormone signaling involved in hepatic lipid metabolism. J. Endocrinol. 2013, 216, R47–R57. [Google Scholar]
- Mariotti, S.; Franceschi, C.; Cossarizza, A.; Pinchera, A. The ageing thyroid. Endocr. Rev 1995, 16, 686–715. [Google Scholar]
- Peeters, R.P. Thyroid hormones and ageing. Hormones 2008, 7, 28–35. [Google Scholar]
- Kemnitz, J.W. Caloric restriction and ageing in nonhuman primates. ILAR J 2011, 8, 66–77. [Google Scholar]
- Roth, G.S.; Handy, A.M.; Mattison, J.A.; Tilmont, E.M.; Ingram, D.K.; Lane, M.A. Effects of dietary caloric restriction and ageing on thyroid hormones of rhesus monkeys. Horm. Metab. Res 2002, 34, 378–382. [Google Scholar]
- Gussekloo, J.; van Exel, E.; de Craen, A.J.; Meinders, A.E.; Frohlich, M.; Westendorp, R.G. Thyroid status, disability and cognitive function, and survival in old age. JAMA 2004, 292, 2591–2599. [Google Scholar]
- Atzmon, G.; Barzilai, N.; Hollowell, J.G.; Surks, M.I.; Gabriely, I. Extreme longevity is associated with increased serum thyrotropin. J. Clin. Endocrinol. Metab 2009, 94, 1251–1254. [Google Scholar]
- Rozing, M.P.; Houwing-Duistermaat, J.J.; Slagboom, P.E.; Beekman, M.; Frohlich, M.; de Craen, A.J.M.; Westendorp, R.G.J.; van Heemst, D. Familial longevity is associated with decreased thyroid function. J. Clin. Endocrinol. Metab 2010, 95, 4979–4984. [Google Scholar]
- Brown-Borg, H.M.; Borg, K.E.; Meliska, C.J.; Bartke, A. Dwarf mice and the ageing process. Nature 1996, 384, 33. [Google Scholar]
- Tatar, M.; Bartke, A.; Antebi, A. The endocrine regulation of ageing by insulin-like signals. Science 2003, 299, 1346–1351. [Google Scholar]
- Vergara, M.; Smith-Wheelock, M.; Harper, J.M.; Sigler, R.; Miller, J.A. Hormone-treated snell dwarf mice regain fertility but remain long lived and disease resistant. J. Gerontol. A. Biol. Sci. Med. Sci 2004, 59, 1244–1250. [Google Scholar]
- Franklyn, J.A.; Sheppard, M.C.; Maisonneuve, P. Thyroid function and mortality in patients treated for hyperthyroidism. JAMA 2005, 294, 71–80. [Google Scholar]
- Cappola, A.R.; Fried, L.P.; Arnold, A.M.; Danese, M.D.; Kuller, L.H.; Burke, G.L.; Tracy, R.P.; Ladenson, P.W. Thyroid status, cardiovascular risk, and mortality in older adults. JAMA 2006, 295, 1033–1041. [Google Scholar]
- Mirza, F.S.; Prestwood, K.M. Bone health and ageing: Implications for menopause. Endocrinol. Metab. Clin. North Am 2004, 33, 741–759. [Google Scholar]
- Yeap, B.B.; Alfonso, H.; Chubb, S.A.P.; Walsh, J.P.; Hankey, G.J.; Almeida, O.P.; Flicker, L. Higher free thyroxine levels are associated with frailty in older men. Clin. Endocrinol 2012, 76, 741–748. [Google Scholar]
- Murphy, E.; Gluer, C.C.; Reid, D.M.; Felsenberg, D.; Roux, C.; Eastell, R.; Williams, G.R. Thyroid function within the upper normal range is associated with reduced menopause bone mineral density and an increased risk of nonvertebral fractures in healthy euthyroid postmenopausal women. J. Clin. Endocrinol. Metab 2010, 95, 3173–3181. [Google Scholar]
- Monfoulet, L.E.; Rabier, B.; Dacquin, R.; Anginot, A.; Photsavang, J.; Jurdic, P.; Vico, L.; Malaval, L.; Chassande, O. Thyroid hormone receptor β mediates thyroid hormone effects on bone remodeling and bone mass. J. Bone Miner. Res 2011, 26, 2036–2044. [Google Scholar]
- Schindhelm, R.K.; ten Boekel, E.; Heima, N.E.; van Schoor, N.M.; Simsek, S. Thyroid hormones and erythrocyte indices in a cohort of euthyroid older subjects. Eur. J. Int. Med 2013, 24, 241–244. [Google Scholar]
- Bremner, A.P.; Feddema, P.; Joska, D.J.; Leedman, P.J.; O’Leary, P.C.; Olynyk, J.K.; Walsh, J.P. Significant association between thyroid hormones and erythrocyte indices in euthyroid subjects. Clin. Endocrinol 2012, 76, 304–311. [Google Scholar]
- Donda, A.; Lemarchand-Beraud, T. Ageing alters the activity of 5′-deiodinase in the adenohypophysis, thyroid gland, and liver of the male rat. Endocrinology 1989, 124, 1305–1309. [Google Scholar]
- Feng, X.; Jiang, Y.; Meltzer, P.; Yen, P.M. Thyroid hormones regulation of hepatic genes in vivo detected by complementary cDNA microarray. Mol. Endocrinol 2000, 14, 947–955. [Google Scholar]
- Clark, P.J. Biology of renal ageing in humans. Adv. Ren. Repl. Ther 2000, 7, 11–21. [Google Scholar]
- Visser, W.E.; Friesema, E.C.; Jansen, J.; Visser, T.J. Thyroid hormone transport by monocarboxylate transporters. Best Pract. Res. Clin. Endocrinol. Metab 2007, 21, 223–236. [Google Scholar]
- Hollowell, J.G.; Staehling, N.W.; Flanders, W.D.; Hannon, W.H.; Gunter, E.W.; Spencer, C.A.; Braverman, L.E. Serum TSH, T4, and thyroid antibodies in the United States population (1988–1994): National health and nutrition examination survey (NHANES III). J. Clin. Endocrinol. Metab 2002, 87, 489–499. [Google Scholar]
- Surks, M.I.; Hollowell, J.G. Age-specific distribution of serum thyrotropin and antithyroid antibodies in the US population: Implications for the prevalence of subclinical hypothyroidism. J. Clin. Endocrinol. Metab 2007, 92, 4575–4582. [Google Scholar]
- Bremner, A.P.; Feddema, P.; Leedman, P.J.; Brown, S.J.; Beilby, J.P.; Lim, E.M.; Wilson, S.G.; O’Leary, P.C.; Walsh, J.P. Age-related changes in thyroid function: A longitudinal study of a community-based cohort. J. Clin. Endocrinol. Metab 2012, 97, 1554–1562. [Google Scholar]
- Waring, A.C.; Arnold, A.M.; Newman, A.B.; Buzkova, P.; Hirsch, C.; Cappola, A.R. Longitudinal changes in thyroid function in the oldest old and survival: The cardiovascular health study all-stars study. J. Clin. Endocrinol. Metab 2012, 97, 3944–3950. [Google Scholar]
- Hadlow, N.C.; Rothacker, K.M.; Wardrop, R.; Brown, S.J.; Lim, E.M.; Walsh, J.P. The relationship between TSH and free T4 in a large population is complex, non-linear and differs by age and gender. J. Clin. Endocrinol. Metab. 2013. [Google Scholar] [CrossRef]
- Nishikawa, M.; Inada, M.; Naito, K.; Ishii, H.; Tanaka, K.; Mashio, Y.; Imura, H. Age-related changes of serum 3,3′-diiodothyronine, 3,5′-diiodothyronine, and 3,5-diiodothyronine concentrations in man. J. Clin. Endocrinol. Metab 1991, 52, 517–522. [Google Scholar]
- Heima, N.E.; Eekhoff, E.M.; Oosterwerff, M.M.; Lips, P.T.; van Schoor, N.M.; Simsek, S. Thryroid function and the metabolic syndrome in older persons: A population-based study. Eur. J. Endocrinol 2012, 168, 59–65. [Google Scholar]
- Waring, A.C.; Rodondi, N.; Harrison, S.; Kanaya, A.M.; Simonsick, E.M.; Milijkovic, I.; Satterfield, S.; Newman, A.B.; Bauer, D.C. Thyroid function and prevalent and incident metabolic syndrome in older adults: The health, ageing and body composition study. Clin. Endocrinol 2012, 76, 911–918. [Google Scholar]
- Larsen, P.R.; Berry, M.J. Nutrition and hormonal regulation of thyroid hormone deiodinases. Ann. Rev. Nutr 1995, 15, 323–352. [Google Scholar]
- Bianco, A.C.; Salvatore, D.; Gereben, B.; Berry, M.; Larsen, P.R. Biochemistry, cellular and molecular biology, and physiological roles of the iodothyronine selenodeodinases. Endocr. Rev 2002, 23, 38–89. [Google Scholar]
- McCann, J.C.; Ames, B.N. Adaptive dysfunction of selenoproteins from the perspective of the triage theory: Why modest selenium deficiency may increase risk of diseases of ageing. FASEB J 2011, 25, 1793–1814. [Google Scholar]
- Frye, R.A. Phylogenetic classification of prokaryotic and eukaryotic sir2-like proteins. Biochem. Biophys. Res. Commun 2000, 273, 793–798. [Google Scholar]
- Sinclair, D.A.; Guarente, L. Extrachromosomal rDNA circles—A cause of ageing in yeast. Cell 1997, 91, 1033–1042. [Google Scholar]
- Tissenbaum, J.A.; Guarente, L. Increased dosage of a sir-2 gene extends lifespan in Caenorhabditis elegans. Nature 2001, 410, 227–230. [Google Scholar]
- Rogina, B.; Helfand, S.L. Sir2 mediates longevity in the fly through a pathway related to calorie restriction. Proc. Natl. Acad. Sci. USA 2004, 101, 15998–16003. [Google Scholar]
- Sasaki, T.; Maier, B.; Bartke, A.; Scrable, H. Progressive loss of SIRT1 with cell cycle withdrawal. Aging Cell 2006, 5, 413–422. [Google Scholar]
- Oberdoerffer, P.; Michan, S.; McVay, M.; Mostoslavsky, R.; Vann, J.; Park, S.K.; Hartlerode, A.; Stegmuller, J.; Hafner, A.; Loerch, P.; et al. SIRT1 redistribution on chromatin promotes genomic stability but alters gene expression during ageing. Cell 2008, 135, 907–918. [Google Scholar]
- Herranz, D.; Munoz-Martin, M.; Canamero, M.; Mulero, F.; Martinez-Pastor, B.; Fernandez-Capetillo, O.; Serrano, M. Sirt1 improves healthy ageing and protects from metabolic syndrome-assocoiated cancer. Nat. Commun. 2010. [Google Scholar] [CrossRef]
- Mostoslavsky, R.; Chua, K.F.; Lombard, D.B.; Pang, W.W.; Fischer, M.R.; Gellon, L.; Liu, P.; Mostoslavsky, G.; Franco, S.; Murphy, M.M.; et al. Genomic instability and ageing-like phenotype in the absence of mammalian SIRT6. Cell 2006, 124, 315–329. [Google Scholar]
- Sack, M.N. The role of SIRT3 in mitochondrial homeostasis and cardiac adaptation to hypertrophy and ageing. J. Mol. Cell. Cardiol 2012, 52, 520–525. [Google Scholar]
- Bordone, L.; Guarente, L. Calorie restriction, SIRT1 and metabolism: Understanding longevity. Nat. Rev. Mol. Cell. Biol 2005, 6, 298–305. [Google Scholar]
- Sinclair, D.A. Toward an unified theory of caloric restriction and longevity regulation. Mech. Ageing Dev 2005, 126, 987–1002. [Google Scholar]
- Zillikens, M.C.; van Meurs, J.B.J.; Rivadeneira, F.; Amin, N.; Hofman, A.; Oostra, B.A.; Sijbrands, E.J.G.; Witteman, J.C.M.; Pols, H.A.P.; van Duijn, C.M.; et al. SIRT1 genetic variation is related to BMI and risk of obesity. Diabetes 2009, 58, 2828–2834. [Google Scholar]
- Eigarska, S.M.; Vonk, J.M.; Boezen, H.M. SIRT1 polymorphism, long-term survival and glucose tolerance in the general population. PLoS One 2013, 8, e58636. [Google Scholar]
- Morris, B.J. Seven sirtuins for seven deadly diseases of ageing. Free Radic. Biol. Med 2013, 56, 133–171. [Google Scholar]
- Baur, J.A.; Ungvari, Z.; Minor, R.K.; Le Couteur, D.G.; de Cabo, R. Are sirtuins viable targets for improving healthspan and lifespan? Nat. Rev. Drug Disc 2012, 11, 443–561. [Google Scholar]
- De Lange, P.; Moreno, M.; Silvestri, E.; Lombardi, A.; Goglia, F.; Lanni, A. Fuel economy in food-deprived skeletal muscle: Signaling pathways and regulatory mechanisms. FASEB J 2007, 21, 3431–3441. [Google Scholar]
- Dasgupta, B.; Ju, J.S.; Sasaki, Y.; Liu, X.; Jung, S.-R.; Higashida, K.; Linquist, D.; Milbrandt, J. The AMPK β2 subunit is required for energy homeostasis during metabolic stress. Mol. Cell. Biol 2012, 32, 2837–2848. [Google Scholar]
- Apfeld, J.; O’Connor, G.; McDonagh, T.; DiStefano, P.S.; Curtis, R. The AMP-activated protein kinase AAK-2 links energy levels and insulin-like signals to lifespan in C. elegans. Genes Dev 2004, 18, 3004–3009. [Google Scholar]
- Greer, E.L.; Banko, M.R.; Brunet, A. AMP-activated protein kinase and FoxO transcription factors in dietary restriction-induced longevity. Ann. N.Y. Acad. Sci 2009, 1170, 688–692. [Google Scholar]
- Qjang, W.; Weiqiang, K.; Qing, Z.; Pengju, Z.; Yi, L. Ageing impairs insulin-stimulated glucose uptake in rat skeletal muscle via suppressing AMPKalpha. Exp. Mol. Med 2007, 39, 535–543. [Google Scholar]
- Reznick, R.M.; Zong, H.; Li, J.; Morino, K.; Moore, I.K.; Yu, H.J.; Liu, Z.X.; Dong, J.; Mustard, K.J.; Hawley, S.A.; et al. Ageing-associated reductions in AMPK-activated protein kinase activity and mitochondrial biogenesis. Cell Metab 2007, 5, 151–156. [Google Scholar]
- Mulligan, J.D.; Gonzalez, A.A.; Kumar, R.; Davis, A.J.; Saupe, K.W. Ageing elevates basal adenosine monophosphate-activated protein kinase (AMPK) activity and eliminates hypoxic activation of AMPK in mouse liver. J. Gerontol. A: Biol. Sci. Med. Sci 2005, 60, 21–27. [Google Scholar]
- McCarty, M.F. Chronic activation of AMP-activated kinase as a strategy for slowing ageing. Med. Hypotheses 2004, 63, 334–339. [Google Scholar]
- Knutson, M.D.; Leeuwenburgh, C. Resveratrol and novel potent activators of SIRT1: Effects on ageing and age-related diseases. Nutr. Rev 2008, 66, 591–596. [Google Scholar]
- Bass, T.M.; Weinkove, D.; Houthoofd, K.; Gems, D.; Partridge, L. Effects of resveratrol on lifespan in Drosophila melanogaster and Caenorhabditis elegans. Mech. Ageing Dev 2007, 128, 546–601. [Google Scholar]
- Barger, J.L.; Kayo, T.; Vann, J.M.; Arias, E.B.; Wang, J.; Hacker, T.A.; Wang, Y.; Raederstorff, D.; Morrow, J.D.; Leeuwenburgh, C.; et al. A low dose of dietary resveratrol partially mimics caloric restriction and retards ageing parameters in mice. PLoS One 2008, 3, e2264. [Google Scholar]
- Agarwal, B.; Baur, J.A. Resveratrol and life extension. Ann. N. Y. Acad. Sci. 2011, 1215, 138–143. [Google Scholar]
- Price, N.L.; Gomes, A.P.; Ling, A.J.Y.; Duarte, F.V.; Martin-Montalvo, A.; North, B.J.; Agarwal, B.; Ye, L.; Ramadori, G.; Teodoro, J.S.; et al. SIRT1 is required for AMPK activation and the beneficial effects of resveratrol on mitochondrial function. Cell Metab 2012, 15, 675–690. [Google Scholar]
- Lagouge, M.; Argmann, C.; Gerhart-Hines, Z.; Meziane, H.; Lerin, C.; Daussin, F.; Messadeq, N.; Milne, J.; Lambert, P.; Elliott, P.; et al. Resveratrol improves mitochondrial function and protects against metabolic disease by activating SIRT1 and PGC-1α. Cell 2006, 127, 1109–1122. [Google Scholar]
- Zang, M.; Xu, S.; Maitland-Toolan, K.A.; Zuccolo, A.; Hou, X.; Jiang, B.; Wierzbicki, M.; Verbeuren, T.J.; Cohen, R.A. Polyphenols stimulate AMP-activated kinase, lower lipids, and inhibit accelerated atherosclerosis in diabetic LDL-receptor deficient mice. Diabetes 2006, 55, 2180–2191. [Google Scholar]
- Park, C.E.; Kim, M.J.; Lee, J.H.; Min, B.I.; Bae, H.; Choe, W.; Kim, S.; Ha, J. Resveratrol stimulates glucose transport in C2C12 myotubes by activating AMP-activated protein kinase. Exp. Mol. Med 2007, 39, 222–229. [Google Scholar]
- Bottner, M.; Christoffel, J.; Rimoldi, G.; Wuttke, W. Effects of long-term treatment with resveratrol and subcutaneous and oral estradiol administration on the pituitary-thyroid axis. Exp. Clin. Endocrinol. Diabetes 2006, 114, 82–90. [Google Scholar]
- Sebai, H.; Hovsépian, S.; Ristorcelli, E.; Aouani, E.; Lombardo, D.; Fayet, G. Resveratrol increases iodide trapping in the rat thyroid cell line FRTL-5. Thyroid 2010, 20, 195–203. [Google Scholar]
- Akieda-Asai, S.; Zaima, N.; Ikegami, N.; Kahyo, T.; Yao, I.; Hatanaka, T.; Iemura, S.-I.; Sugiyama, R.; Yokozeki, T.; Eishi, Y.; et al. SIRT1 regulates thyroid-stimulating hormone release by enhancing PIP5Kγ activity through deacetylation of specific lysine residues in mammals. PLoS One 2010, 5, e11755. [Google Scholar]
- Lombardi, A.; Silvestri, E.; Cioffi, F.; Senese, R.; Lanni, A.; Goglia, F.; de Lange, P.; Moreno, M. Defining the transcriptomic and proteomic profiles of rat ageing skeletal muscle by the use of a cDNA array, 2D- and Blue native-PAGE approach. J. Proteomics 2009, 72, 708–721. [Google Scholar]
- Nabben, M.; Hoeks, J.; Briedé, J.J.; Glatz, J.E.; Moonen-Kornips, E.; Hesselink, M.K.; Schrauwen, P. The effect of UCP3 overexpression on mitochondrial ROS production in skeletal muscle of young versus aged mice. FEBS Lett 2008, 582, 4147–4152. [Google Scholar]
- Lombardi, A.; Busiello, R.A.; Napolitano, L.; Cioffi, F.; Moreno, M.; de Lange, P.; Silvestri, E.; Lanni, A.; Goglia, F. UCP3 translocates lipid hydroperoxide and mediates lipid hydroperoxide-dependent mitochondrial uncoupling. J. Biol. Chem 2010, 285, 16599–16605. [Google Scholar]
- Videla, L.A.; Fernandez, V.; Cornejo, P.; Vargas, R. Metabolic basis for thyroid hormone liver preconditioning: Upregulation of AMP-activated protein kinase signaling. Sci. World J 2012, 2012, 475675, :1–475675:10.. [Google Scholar]
- Fernandez, V.; Castillo, I.; Tapia, G.; Romanque, P.; Uribe-Echevarria, S.; Uribe, M.; Cartier-Ugarte, D.; Santander, G.; Vial, M.T.; Videla, L.A. Thyroid hormone preconditioning: Protection against ischemia-reperfusion liver injury in the rat. Hepatology 2007, 45, 170–177. [Google Scholar]
- Pantos, C.I.; Malliopoulou, V.A.; Mourouzis, I.S.; Karamanoli, E.P.; Paizis, I.A.; Steimberg, N.; Varonos, D.D.; Cokkinos, D.V. Long-term thyroxine administration protects the heart in a pattern similar to ischemic preconditioning. Thyroid 2002, 12, 325–329. [Google Scholar]
- Li, F.; Lu, S.; Zhu, R.; Zhou, Z.; Ma, L.; Cai, L.; Liu, Z. Heme-oxygenase-1 is induced by thyroid hormone and involved in thyroid hormone preconditioning-induced protection against renal warm ischemia in rat. Mol. Cell. Endocrinol 2011, 339, 54–62. [Google Scholar]
- Wojtovich, A.P.; Nadtochiy, S.M.; Brookes, P.S.; Nehrke, K. Ischemic preconditioning: The role of mitochondrial and ageing. Exp. Gerontol 2012, 47, 1–7. [Google Scholar]
- Avogaro, A.; Vigili de Kreutzenberg, A.; Fadini, G.P. Insulin signaling and life span. Pflug. Arch 2010, 459, 301–314. [Google Scholar]
- Brenta, G. Why can insulin resistance be a natural consequence of thyroid dysfunction? J. Thyroid Res 2011, 2011, 152850, :1–152850:9.. [Google Scholar]
- Peeters, R.P.; van der Deure, W.M.; van den Beld, A.W.; van Toor, H.; Lamberts, S.W.; Janssen, J.A.; Uitterlinden, A.G.; Visser, T.J. The Asp727Glu polymorphism in the TSH receptor is associated with insulin resistance in healthy elderly men. Clin. Endocrinol 2007, 66, 808–815. [Google Scholar]
- Peeters, R.P.; van den Beld, A.W.; van Toor, H.; Uitterlinden, A.G.; Janssen, J.A.; Lamberts, S.W.; Visser, T.J. A polymorphism in type I deiodinase is associated with circulating free insulin-like growth factor I levels and body composition in humans. J. Clin. Endocrinol. Metab 2005, 90, 256–263. [Google Scholar]
- Canto, C.; Gerhardt-Hines, Z.; Feige, J.N.; Lagouge, M.; Noriega, L.; Milne, J.C.; Elliott, P.J.; Puigserver, P.; Auwerx, J. AMPK regulates energy expenditure by modulating NAD+ metabolism and SIRT1 activity. Nature 2009, 458, 1056–1060. [Google Scholar]
- Hou, X.; Xu, S.; Maitland-Toolan, K.A.; Sato, K.; Jiang, B.; Ido, Y.; Lan, F.; Walsh, K.; Wierzbicki, M.; Verbeuren, T.J.; et al. SIRT1 regulates hepatocyte lipid metabolism through activating AMP-activating protein kinase. J. Biol. Chem 2008, 283, 20015–20026. [Google Scholar]
- Jager, S.; Handschin, C.; St. Pierre, J.; Spiegelman, B.M. AMP-activated protein kinase (AMPK) action in skeletal muscle via direct phosphorylaiton of PGC-1alpha. Proc. Natl. Acad. Sci. USA 2007, 104, 12017–12022. [Google Scholar]
- Ponugoti, B.; Kim, D.-H.; Xiao, Z.; Smith, Z.; Miao, J.; Zang, M.; Wu, S.-Y.; Chiang, C.-M.; Veenstra, T.D.; Kemper, J.K. SIRT1 deacetylates and inhibits SREBP-1c activity in regulation of hepatic lipid metabolism. J. Biol. Chem 2010, 285, 33959–33970. [Google Scholar]
- Greer, E.L.; Oskoui, P.R.; Banko, M.R.; Maniar, J.M.; Gygi, M.P.; Gygi, S.P.; Brunet, A. The energy sensor AMP-activated protein kinase directly regulates the mammalian FOXO3 transcription. J. Biol. Chem 2007, 282, 30107–30119. [Google Scholar]
- Brunet, A.; Sweeney, L.B.; Sturgill, J.F.; Chua, K.F.; Greer, P.L.; Lin, Y.; Tran, H.; Ross, S.E.; Mostoslavsky, R.; Cohen, H.Y.; et al. Stress-dependent regulation of FOXO transcription factors by the SIRT1 deacetylase. Science 2004, 303, 2011–2015. [Google Scholar]
- Canto, C.; Auwerx, J. Targeting sirtuin 1 to improve metabolism: All you need is NAD+? Pharm. Rev 2012, 64, 166–187. [Google Scholar]
- Moreno, M.; Lombardi, A.; Beneduce, L.; Silvestri, E.; Pinna, G.; Goglia, F.; Lanni, A. Are the effects of T3 on resting metabolic rate in euthyroid rats entirely caused by T3 itself? Endocrinology 2002, 143, 504–510. [Google Scholar]
- Li, Y.; Xu, S.; Giles, A.; Nakamura, K.; Lee, J.W.; Hou, X.; Donmez, G.; Li, J.; Luo, Z.; Walsh, K.; et al. Hepatic overexpression of SIT1 in mice attenuates endoplasmic reticulum stress and insulin resistance in the liver. FASEB J 2011, 25, 1664–1679. [Google Scholar]
- Feige, J.N.; Lagouge, M.; Canto, C.; Strehle, A.; Houten, S.M.; Milne, J.C.; Lambert, P.D.; Mataki, C.; Elliott, P.J.; Auwerx, J. Specific SIRT1 activation mimics low energy levels and protects against diet-induced metabolic disorders by enhancing fat oxidation. Cell Metab 2008, 8, 347–358. [Google Scholar]
- Purushotham, A.; Xu, Q.; Li, X. Systemic SIRT1 insufficiency results in disruption of energy homeostasis and steroid hormone metabolism upon high-fat-diet feeding. FASEB J 2012, 26, 656–667. [Google Scholar]
- Beck Jorgensen, S.; O’Neill, H.M.; Hewitt, K.; Kemp, B.E.; Steinberg, G.R. Reduced AMPK-activated protein kinase activity in mouse skeletal muscle does not exacerbate the development of insulin resistance with obesity. Diabetologia 2009, 52, 2395–2404. [Google Scholar]
- Jun, M.; Perkovic, I.; Cass, A. Intensive glycemic control and renal outcome. Contrib. Nephrol 2011, 170, 196–208. [Google Scholar]
- Maeda, S.; Koya, D.; Araki, S.; Babazono, T.; Umezono, T.; Toyoda, M.; Kawai, K.; Imanishi, M.; Uzu, T.; Suzuki, D.; et al. Association between single nucleotide polymorphisms within genes encoding sirtuin families and diabetic nephropathy in Japanese subjects with type 2 diabetes. Clin. Exp. Nephrol 2011, 15, 381–390. [Google Scholar]
- Shang, G.; Gao, P.; Zhao, Z.; Chen, Q.; Jiang, T.; Zhang, N.; Li, H. 3,5-diiodo-l-thyronine ameliorates diabetic nephropathy in streptozotocin-induced diabetic rats. Biochim. Biophys. Acta 2013, 1832, 674–684. [Google Scholar]
- Hashimoto, K.; Cohen, R.N.; Yamada, M.; Markan, K.R.; Monden, T.; Satoh, T.; Mori, M.; Wondisford, F.E. Cross-talk between thyroid hormone receptor and liver X receptor regulatory pathways is revealed in a thyroid hormone resistance mouse model. J. Biol. Chem 2006, 281, 295–302. [Google Scholar]
- Thakran, S.; Sharma, P.; Attia, R.R.; Hori, R.T.; Deng, X.; Elam, M.B.; Park, E.A. Role of sirtuin 1 in the regulation of hepatic gene expression by thyroid hormone. J. Biol. Chem 2013, 288, 807–818. [Google Scholar]
- Frenzel, M.; Rommelspacher, H.; Sugawa, M.D.; Dencher, N.A. Ageing alters the supramolecular architecture of OxPhos complexes in rat brain cortex. Exp. Gerontol 2010, 45, 563–572. [Google Scholar]
- Gomez, L.A.; Monette, J.S.; Chavez, J.D.; Maier, C.S.; Hagen, T.M. Supercomplexes of the mitochondrial electron transport chain decline in the ageing rat heart. Arch. Biochem. Biophys 2009, 490, 30–35. [Google Scholar]
- Gates, A.C.; Bernal-Mizrachi, C.; Chinault, S.L.; Feng, C.; Schneider, J.G.; Coleman, T.; Malone, J.P.; Townsend, R.R.; Chakravarty, M.V.; Semenkovich, C.F. Respiratory uncoupling in skeletal muscle delays death and diminishes age-related disease. Cell Metab 2007, 6, 497–505. [Google Scholar]
- Speakman, J.R.; Talbot, D.A.; Selman, C.; Snart, C.; McLaren, J.S.; Redman, P.; Krol, E.; Jackson, D.M.; Johnson, M.S.; Brand, M.D. Uncoupled and surviving: Individual mice with high metabolism have greater mitochondrial uncoupling and live longer. Aging Cell 2004, 3, 87–95. [Google Scholar]
- Amara, C.E.; Shankland, E.G.; Jubrias, S.A.; Marcinek, D.J.; Kushmerick, M.J.; Conley, K.E. Mild mitochondrial uncoupling impacts cellular aging in human muscles in vivo. Proc. Natl. Acad. Sci. USA 2007, 104, 1057–1062. [Google Scholar]
- Kerner, J.; Turkaly, P.J.; Minkler, P.E.; Hoppel, C.L. Ageing skeletal muscle mitochondria in the rat: Decreased uncoupling protein-3 content. Am. J. Physiol. Endocrinol. Metab 2001, 281, E1054–E1062. [Google Scholar]
- Lanni, A.; Beneduce, L.; Lombardi, A.; Moreno, M.; Boss, O.; Muzzin, P.; Giacobino, J.P.; Goglia, F. Expression of uncoupling protein-3 and mitochondrial activity in the transition from hypothyroid to hyperthyroid state in rat skeletal muscle. FEBS Lett 1999, 444, 250–254. [Google Scholar]
- De Lange, P.; Lanni, A.; Beneduce, L.; Moreno, M.; Lombardi, A.; Silvestri, E.; Goglia, F. Uncoupling protein-3 is a molecular determinant for the regulation of resting metabolic rate by thyroid hormone. Endocrinology 2001, 142, 3414–3420. [Google Scholar]


{kind=link}
| Hormone | Young | Adult | Old | Species and References |
|---|---|---|---|---|
| TSH | 100 | 100 | 150 | Human [47–51] |
| 100 | 100 | 50 | Rhesus monkey [28] | |
| 100 | 100 | 100 | Rat [4] | |
| T4 | 100 | 100 | 100 | Human [25,26] |
| 100 | 100 | 65 | Rhesus monkey [28] | |
| 100 | 69 | 47 | Rat [4] | |
| FT4 | 100 | 100 | 100 | Human [25,26,47–51] |
| 100 | 100 | 65 | Rhesus monkey [28] | |
| 100 | 52 | 21 | Rat [4] | |
| T3 | 100 | 100 | 50 | Human [25,26] |
| 100 | 100 | <100 (N.S.) | Rhesus monkey [28] | |
| 100 | 100 | 29 | Rat [4] | |
| FT3 | 100 | 100 | 50 | Human [25,26] |
| 100 | 100 | 50 | Rat [4] | |
| 3,3′-T2 | 100 | N.D. | 62 | Human [52] |
| 3,5′-T2 | 100 | N.D. | 42 | Human [52] |
| 3,5-T2 (T2) | 100 | N.D. | 53 | Human [52] |
© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
De Lange, P.; Cioffi, F.; Silvestri, E.; Moreno, M.; Goglia, F.; Lanni, A. (Healthy) Ageing: Focus on Iodothyronines. Int. J. Mol. Sci. 2013, 14, 13873-13892. https://doi.org/10.3390/ijms140713873
De Lange P, Cioffi F, Silvestri E, Moreno M, Goglia F, Lanni A. (Healthy) Ageing: Focus on Iodothyronines. International Journal of Molecular Sciences. 2013; 14(7):13873-13892. https://doi.org/10.3390/ijms140713873
Chicago/Turabian StyleDe Lange, Pieter, Federica Cioffi, Elena Silvestri, Maria Moreno, Fernando Goglia, and Antonia Lanni. 2013. "(Healthy) Ageing: Focus on Iodothyronines" International Journal of Molecular Sciences 14, no. 7: 13873-13892. https://doi.org/10.3390/ijms140713873
APA StyleDe Lange, P., Cioffi, F., Silvestri, E., Moreno, M., Goglia, F., & Lanni, A. (2013). (Healthy) Ageing: Focus on Iodothyronines. International Journal of Molecular Sciences, 14(7), 13873-13892. https://doi.org/10.3390/ijms140713873