Enhanced Development of Azoxymethane-Induced Colonic Preneoplastic Lesions in Hypertensive Rats

Abstract
:1. Introduction
2. Results and Discussion
2.1. General Observations
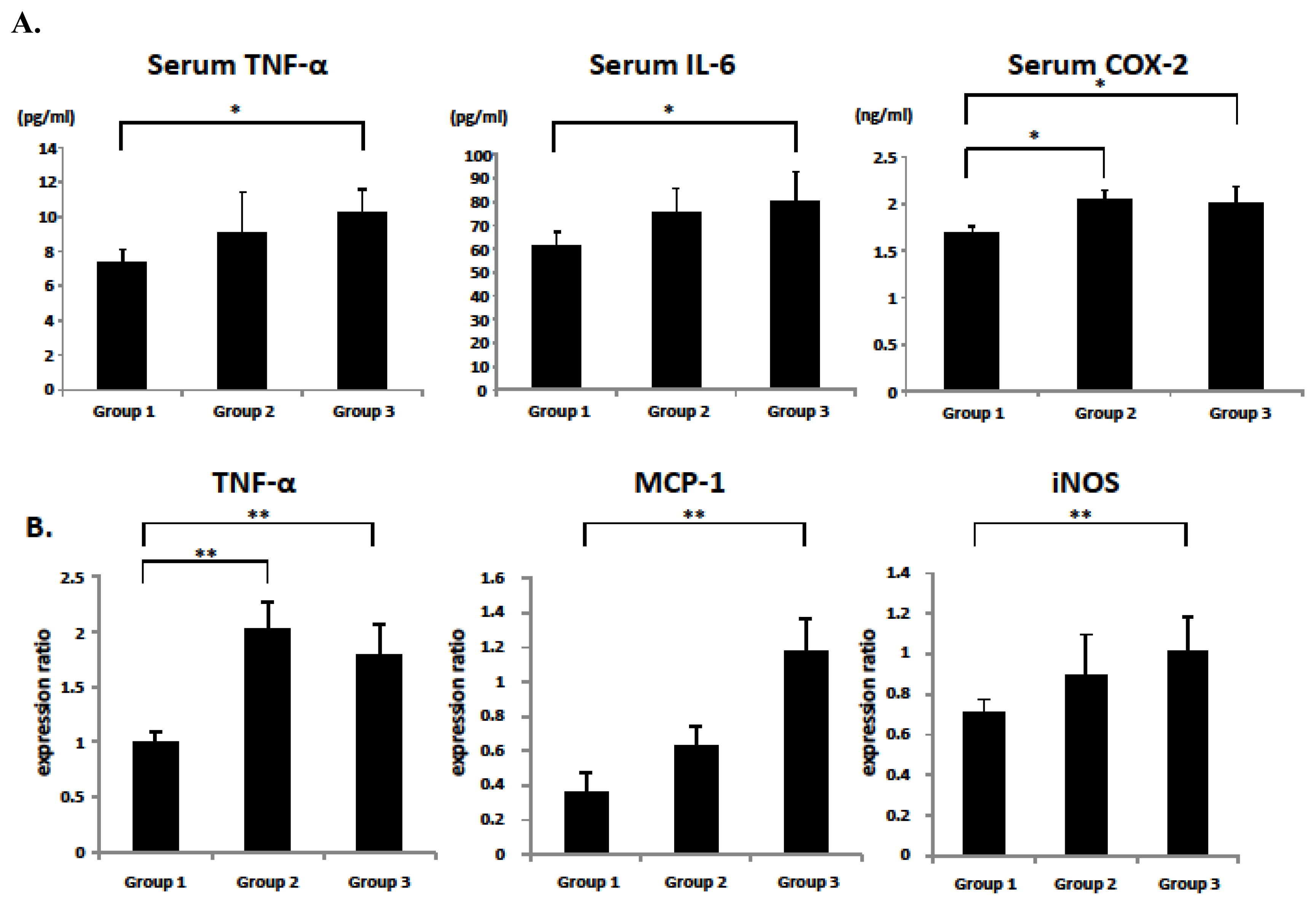
2.2. Serum Parameters of the Experimental Rats
2.3. Development of Colonic Preneoplastic Lesions
2.4. Systemic Oxidative Stress and Colonic Epithelial Expression of GPx and CAT mRNA
2.5. Serum and Colonic Epithelial Expression of Inflammatory Markers
2.6. Discussion
3. Experimental Section
3.1. Animals and Chemicals
3.2. Experimental Procedure
3.3. Enumeration of ACF
3.4. RNA Extraction and Quantitative Real-Time Reverse Transcription-Polymerase Chain Reaction Analysis
3.5. Clinical Biochemistry
3.6. Oxidative Stress Analysis
3.7. Statistical Analysis
4. Conclusions
Supplementary Information
ijms-14-14700-s001.pdfConflict of Interest
References
- Alberti, K.G.; Zimmet, P.; Shaw, J. IDF Epidemiology Task Force Consensus Group. The metabolic syndrome—A new worldwide definition. Lancet 2005, 366, 1059–1062. [Google Scholar]
- Grundy, S.M.; Cleeman, J.I.; Daniels, S.R.; Donato, K.A.; Eckel, R.H.; Franklin, B.A.; Gordon, D.J.; Krauss, R.M.; Savage, P.J.; Smith, S.C., Jr; et al. Diagnosis and management of the metabolic syndrome: An American Heart Association/National Heart, Lung, and Blood Institute Scientific Statement. Circulation 2005, 112, 2735–2752. [Google Scholar]
- Ahmed, R.L.; Schmitz, K.H.; Anderson, K.E.; Rosamond, W.D.; Folsom, A.R. The metabolic syndrome and risk of incident colorectal cancer. Cancer 2006, 107, 28–36. [Google Scholar]
- Stocks, T.; van Hemelrijck, M.; Manjer, J.; Bjorge, T.; Ulmer, H.; Hallmans, G.; Lindkvist, B.; Selmer, R.; Nagel, G.; Tretli, S.; et al. Blood pressure and risk of cancer incidence and mortality in the Metabolic Syndrome and Cancer Project. Hypertension 2012, 59, 802–810. [Google Scholar]
- Ishino, K.; Mutoh, M.; Totsuka, Y.; Nakagama, H. Metabolic syndrome: A novel high-risk state for colorectal cancer. Cancer Lett. 2012. [Google Scholar] [CrossRef]
- Shimizu, M.; Kubota, M.; Tanaka, T.; Moriwaki, H. Nutraceutical approach for preventing obesity-related colorectal and liver carcinogenesis. Int. J. Mol. Sci 2012, 13, 579–595. [Google Scholar]
- Yuhara, H.; Steinmaus, C.; Cohen, S.E.; Corley, D.A.; Tei, Y.; Buffler, P.A. Is diabetes mellitus an independent risk factor for colon cancer and rectal cancer? Am. J. Gastroenterol 2011, 106, 1911–1921. [Google Scholar]
- Donohoe, C.L.; Pidgeon, G.P.; Lysaght, J.; Reynolds, J.V. Obesity and gastrointestinal cancer. Br. J. Surg 2010, 97, 628–642. [Google Scholar]
- De Kloet, A.D.; Krause, E.G.; Woods, S.C. The renin angiotensin system and the metabolic syndrome. Physiol. Behav 2010, 100, 525–534. [Google Scholar]
- Ager, E.I.; Neo, J.; Christophi, C. The renin-angiotensin system and malignancy. Carcinogenesis 2008, 29, 1675–1684. [Google Scholar]
- Deshayes, F.; Nahmias, C. Angiotensin receptors: A new role in cancer? Trends Endocrinol. Metab 2005, 16, 293–299. [Google Scholar]
- Paul, M.; Poyan Mehr, A.; Kreutz, R. Physiology of local renin-angiotensin systems. Physiol. Rev 2006, 86, 747–803. [Google Scholar]
- Fyhrquist, F.; Saijonmaa, O. Renin-angiotensin system revisited. J. Intern. Med 2008, 264, 224–236. [Google Scholar]
- George, A.J.; Thomas, W.G.; Hannan, R.D. The renin-angiotensin system and cancer: Old dog, new tricks. Nat. Rev. Cancer 2010, 10, 745–759. [Google Scholar]
- Hiraoka-Yamamoto, J.; Nara, Y.; Yasui, N.; Onobayashi, Y.; Tsuchikura, S.; Ikeda, K. Establishment of a new animal model of metabolic syndrome: SHRSP fatty (fa/fa) rats. Clin. Exp. Pharmacol. Physiol 2004, 31, 107–109. [Google Scholar]
- Ueno, T.; Takagi, H.; Fukuda, N.; Takahashi, A.; Yao, E.H.; Mitsumata, M.; Hiraoka-Yamamoto, J.; Ikeda, K.; Matsumoto, K.; Yamori, Y. Cardiovascular remodeling and metabolic abnormalities in SHRSP.Z-Lepr(fa)/IzmDmcr rats as a new model of metabolic syndrome. Hypertens. Res. Off. J. Jpn. Soc. Hypertens 2008, 31, 1021–1031. [Google Scholar]
- Chen, H.; Sullivan, G.; Yue, L.Q.; Katz, A.; Quon, M.J. QUICKI is a useful index of insulin sensitivity in subjects with hypertension. Am. J. Physiol. Endocrinol. Metab 2003, 284, E804–E812. [Google Scholar]
- Chrysant, S.G.; Chrysant, G.S.; Chrysant, C.; Shiraz, M. The treatment of cardiovascular disease continuum: Focus on prevention and RAS blockade. Curr. Clin. Pharmacol 2010, 5, 89–95. [Google Scholar]
- Kochi, K.; Shimizu, M.; Ohno, T.; Baba, A.; Sumi, T.; Kubota, M.; Shirakami, Y.; Tsurumi, H.; Tanaka, T.; Moriwaki, H. Department of Internal Medicine, Gifu University Graduate School of Medicine, 2013; Unpublished data.
- Tudek, B.; Speina, E. Oxidatively damaged DNA and its repair in colon carcinogenesis. Mutat. Res 2012, 736, 82–92. [Google Scholar]
- Suzuki, Y.; Imai, K.; Takai, K.; Hanai, T.; Hayashi, H.; Naiki, T.; Nishigaki, Y.; Tomita, E.; Shimizu, M.; Moriwaki, H. Hepatocellular carcinoma patients with increased oxidative stress levels are prone to recurrence after curative treatment: A prospective case series study using the d-ROM test. J. Cancer Res. Clin. Oncol. 2013, in press. [Google Scholar]
- Hotamisligil, G.S.; Shargill, N.S.; Spiegelman, B.M. Adipose expression of tumor necrosis factor-alpha: Direct role in obesity-linked insulin resistance. Science 1993, 259, 87–91. [Google Scholar]
- Hotamisligil, G.S. Inflammation and metabolic disorders. Nature 2006, 444, 860–867. [Google Scholar]
- Szlosarek, P.; Charles, K.A.; Balkwill, F.R. Tumour necrosis factor-alpha as a tumour promoter. Eur. J. Cancer 2006, 42, 745–750. [Google Scholar]
- Gupta, R.A.; Dubois, R.N. Colorectal cancer prevention and treatment by inhibition of cyclooxygenase-2. Nat. Rev. Cancer 2001, 1, 11–21. [Google Scholar]
- Cassis, P.; Conti, S.; Remuzzi, G.; Benigni, A. Angiotensin receptors as determinants of life span. Pflugers Arch 2010, 459, 325–332. [Google Scholar]
- Smith, G.R.; Missailidis, S. Cancer, inflammation and the AT1 and AT2 receptors. J. Inflam 2004, 1, 3. [Google Scholar]
- Massiera, F.; Bloch-Faure, M.; Ceiler, D.; Murakami, K.; Fukamizu, A.; Gasc, J.M.; Quignard-Boulange, A.; Negrel, R.; Ailhaud, G.; Seydoux, J.; et al. Adipose angiotensinogen is involved in adipose tissue growth and blood pressure regulation. FASEB 2001, 15, 2727–2729. [Google Scholar]
- Yvan-Charvet, L.; Massiera, F.; Lamande, N.; Ailhaud, G.; Teboul, M.; Moustaid-Moussa, N.; Gasc, J.M.; Quignard-Boulange, A. Deficiency of angiotensin type 2 receptor rescues obesity but not hypertension induced by overexpression of angiotensinogen in adipose tissue. Endocrinology 2009, 150, 1421–1428. [Google Scholar]
- Uemura, H.; Ishiguro, H.; Nagashima, Y.; Sasaki, T.; Nakaigawa, N.; Hasumi, H.; Kato, S.; Kubota, Y. Antiproliferative activity of angiotensin II receptor blocker through cross-talk between stromal and epithelial prostate cancer cells. Mol. Cancer Ther 2005, 4, 1699–1709. [Google Scholar]
- Uemura, H.; Ishiguro, H.; Ishiguro, Y.; Hoshino, K.; Takahashi, S.; Kubota, Y. Angiotensin II induces oxidative stress in prostate cancer. Mol. Cancer Res. MCR 2008, 6, 250–258. [Google Scholar]
- Lever, A.F.; Hole, D.J.; Gillis, C.R.; McCallum, I.R.; McInnes, G.T.; MacKinnon, P.L.; Meredith, P.A.; Murray, L.S.; Reid, J.L.; Robertson, J.W. Do inhibitors of angiotensin-I-converting enzyme protect against risk of cancer? Lancet 1998, 352, 179–184. [Google Scholar]
- Lang, L. ACE inhibitors may reduce esophageal cancer incidence. Gastroenterology 2006, 131, 343–344. [Google Scholar]
- Sipahi, I.; Chou, J.; Mishra, P.; Debanne, S.M.; Simon, D.I.; Fang, J.C. Meta-analysis of randomized controlled trials on effect of angiotensin-converting enzyme inhibitors on cancer risk. Am. J. Cardiol 2011, 108, 294–301. [Google Scholar]
- Hallas, J.; Christensen, R.; Andersen, M.; Friis, S.; Bjerrum, L. Long term use of drugs affecting the renin-angiotensin system and the risk of cancer: A population-based case-control study. Br. J. Clin. Pharmacol 2012, 74, 180–188. [Google Scholar]
- Shimomoto, T.; Ohmori, H.; Luo, Y.; Chihara, Y.; Denda, A.; Sasahira, T.; Tatsumoto, N.; Fujii, K.; Kuniyasu, H. Diabetes-associated angiotensin activation enhances liver metastasis of colon cancer. Clin. Exp. Metast 2012, 29, 915–925. [Google Scholar]
- Kubota, M.; Shimizu, M.; Sakai, H.; Yasuda, Y.; Ohno, T.; Kochi, T.; Tsurumi, H.; Tanaka, T.; Moriwaki, H. Renin-angiotensin system inhibitors suppress azoxymethane-induced colonic preneoplastic lesions in C57BL/KsJ-db/db obese mice. Biochem. Biophys. Res. Commun 2011, 410, 108–113. [Google Scholar]
- Takahashi, M.; Wakabayashi, K. Gene mutations and altered gene expression in azoxymethane-induced colon carcinogenesis in rodents. Cancer Sci 2004, 95, 475–480. [Google Scholar]
- Chen, J.; Huang, X.F. The signal pathways in azoxymethane-induced colon cancer and preventive implications. Cancer Biol. Ther 2009, 8, 1313–1317. [Google Scholar]
- Reddy, B.S. Studies with the azoxymethane-rat preclinical model for assessing colon tumor development and chemoprevention. Environ. Mol. Mutagenesis 2004, 44, 26–35. [Google Scholar]
- Raju, J. Azoxymethane-induced rat aberrant crypt foci: Relevance in studying chemoprevention of colon cancer. World J. Gastroenterol 2008, 14, 6632–6635. [Google Scholar]
- Ogawa, K.; Hara, T.; Shimizu, M.; Ninomiya, S.; Nagano, J.; Sakai, H.; Hoshi, M.; Ito, H.; Tsurumi, H.; Saito, K.; et al. Suppression of azoxymethane-induced colonic preneoplastic lesions in rats by 1-methyltryptophan, an inhibitor of indoleamine 2,3-dioxygenase. Cancer Sci 2012, 103, 951–918. [Google Scholar]
- Primer Blast. Available online: http://www.ncbi.nlm.nih.gov/tools/primer-blast/ (on accessed 12 July 2012).




{kind=link}
{kind=link}
{kind=link}
| Group NO. | Strain | No. | Body weight (g) | Relative adipose tissue weight (g/100g body weight) a | Blood pressure (mmHg) | |
|---|---|---|---|---|---|---|
| Systolic | Diastolic | |||||
| Group 1 | WKY b | 8 | 256.5 ± 11.7 e | 0.72 ± 0.16 | 127 ± 12.8 | 92 ± 4.9 |
| Group 2 | SHRSP c | 8 | 218.9 ± 8.0 f | 0.77 ± 0.16 | 188 ± 12.5 f | 141 ± 10.6 f |
| Group 3 | SHRSP-ZF d | 8 | 270.1 ± 23.4 g | 1.64 ± 0.17 f,g | 169 ± 13.7 f,g | 129 ± 9.0 f,g |
| Glucose (mg/dL) | Insulin (μIU/mL) | Quicki | Leptin (pg/mL) | NEFA (mEq/L) | Triglyceride (mg/dL) | Angiotensin II (ng/mL) | |
|---|---|---|---|---|---|---|---|
| Group 1 | 85.4 ± 11.7 | 15.81 ± 0.35 | 0.313 ± 0.010 | 11.2 ± 3.6 | 0.459 ± 0.03 | 27.1 ± 7.4 | 352.6 ± 38.1 |
| Group 2 | 83.5 ± 12.3 | 17.00 ± 1.39 | 0.320 ± 0.008 | 12.2 ± 3.4 | 0.419 ± 0.05 | 39.6 ± 14.1 | 494.4 ± 75.6 b |
| Group 3 | 120.0 ± 14.2 b,c | 25.60 ± 8.98 b,c | 0.291 ± 0.010 b,c | 102.7 ± 30.6 b,c | 0.538 ± 0.03 b,c | 257.1 ± 79.4 b,c | 500.9 ± 42.5 b |
© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Kochi, T.; Shimizu, M.; Ohno, T.; Baba, A.; Sumi, T.; Kubota, M.; Shirakami, Y.; Tsurumi, H.; Tanaka, T.; Moriwaki, H. Enhanced Development of Azoxymethane-Induced Colonic Preneoplastic Lesions in Hypertensive Rats. Int. J. Mol. Sci. 2013, 14, 14700-14711. https://doi.org/10.3390/ijms140714700
Kochi T, Shimizu M, Ohno T, Baba A, Sumi T, Kubota M, Shirakami Y, Tsurumi H, Tanaka T, Moriwaki H. Enhanced Development of Azoxymethane-Induced Colonic Preneoplastic Lesions in Hypertensive Rats. International Journal of Molecular Sciences. 2013; 14(7):14700-14711. https://doi.org/10.3390/ijms140714700
Chicago/Turabian StyleKochi, Takahiro, Masahito Shimizu, Tomohiko Ohno, Atsushi Baba, Takafumi Sumi, Masaya Kubota, Yohei Shirakami, Hisashi Tsurumi, Takuji Tanaka, and Hisataka Moriwaki. 2013. "Enhanced Development of Azoxymethane-Induced Colonic Preneoplastic Lesions in Hypertensive Rats" International Journal of Molecular Sciences 14, no. 7: 14700-14711. https://doi.org/10.3390/ijms140714700
APA StyleKochi, T., Shimizu, M., Ohno, T., Baba, A., Sumi, T., Kubota, M., Shirakami, Y., Tsurumi, H., Tanaka, T., & Moriwaki, H. (2013). Enhanced Development of Azoxymethane-Induced Colonic Preneoplastic Lesions in Hypertensive Rats. International Journal of Molecular Sciences, 14(7), 14700-14711. https://doi.org/10.3390/ijms140714700