MicroRNA-143 Downregulates Interleukin-13 Receptor Alpha1 in Human Mast Cells


Abstract
:1. Introduction
2. Results
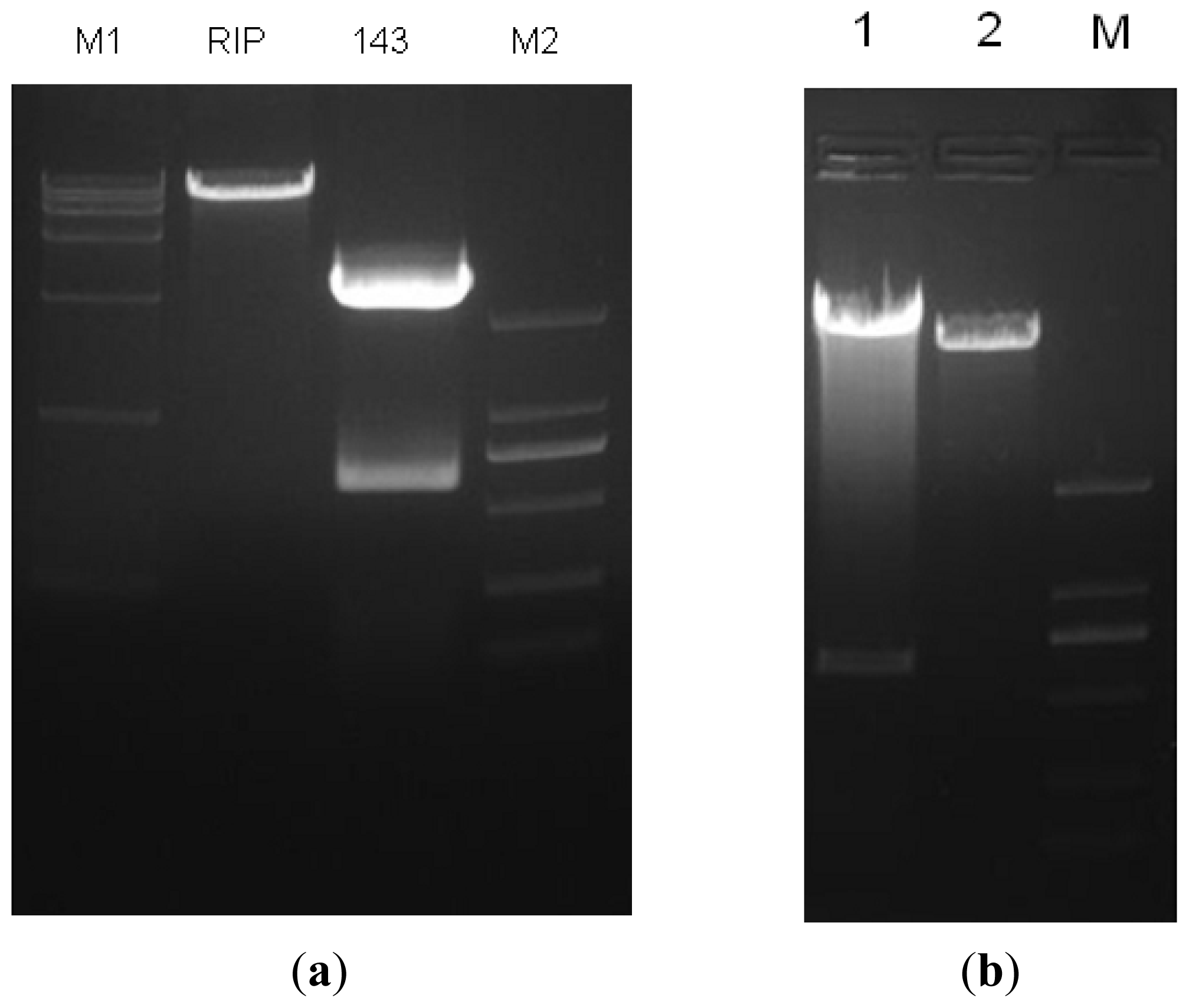
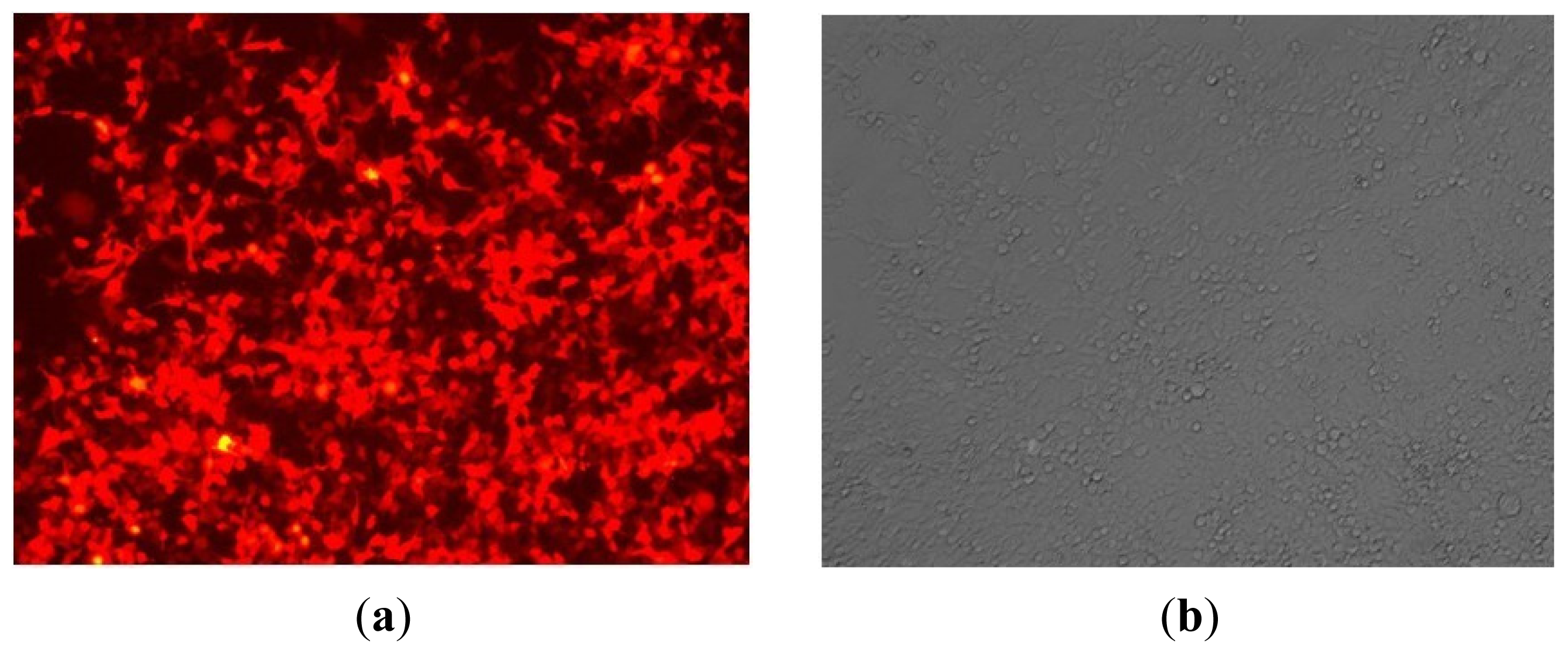
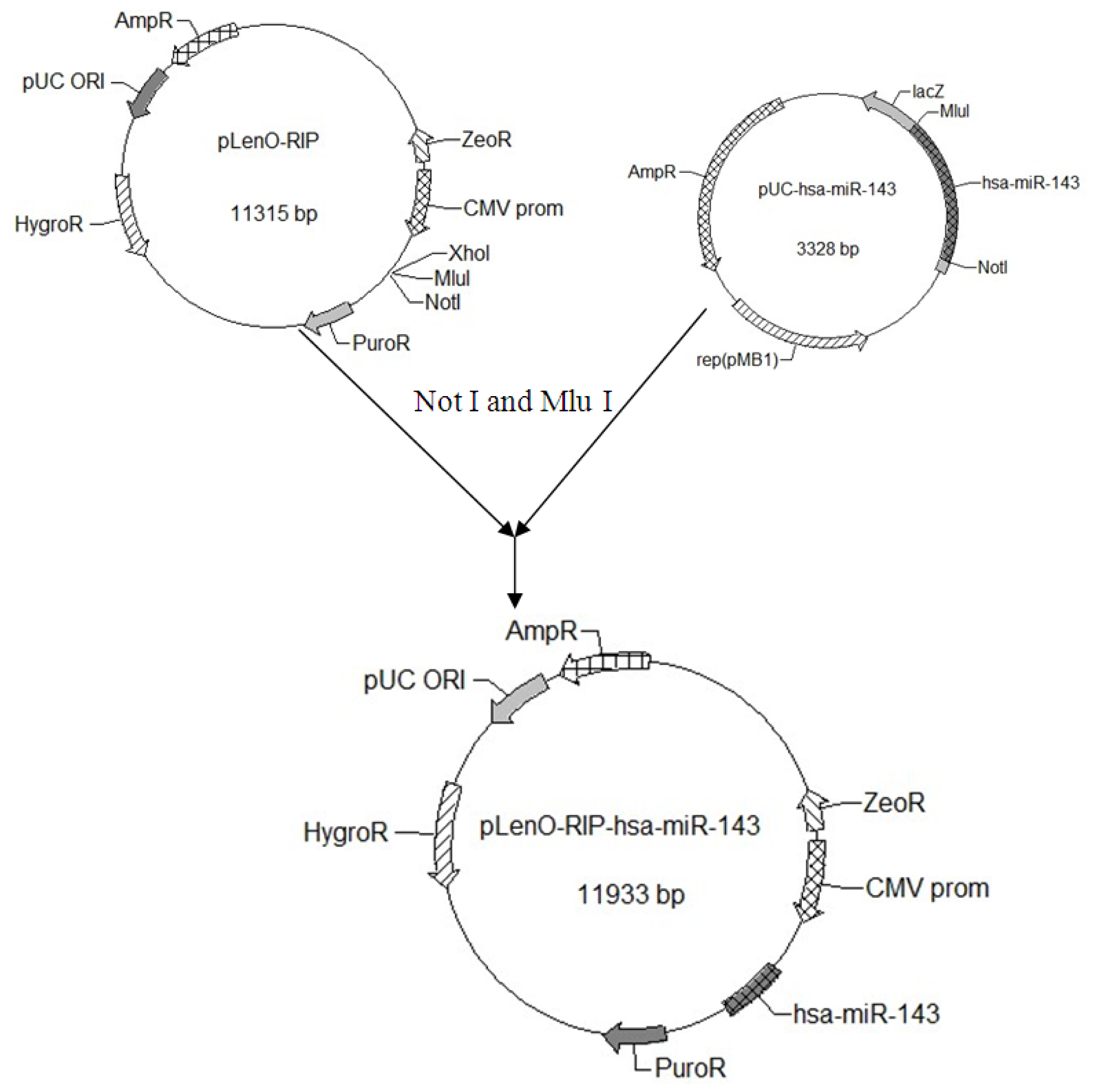
2.1. Construction and Identification of miR-143 pLenO-RIP Plasmid
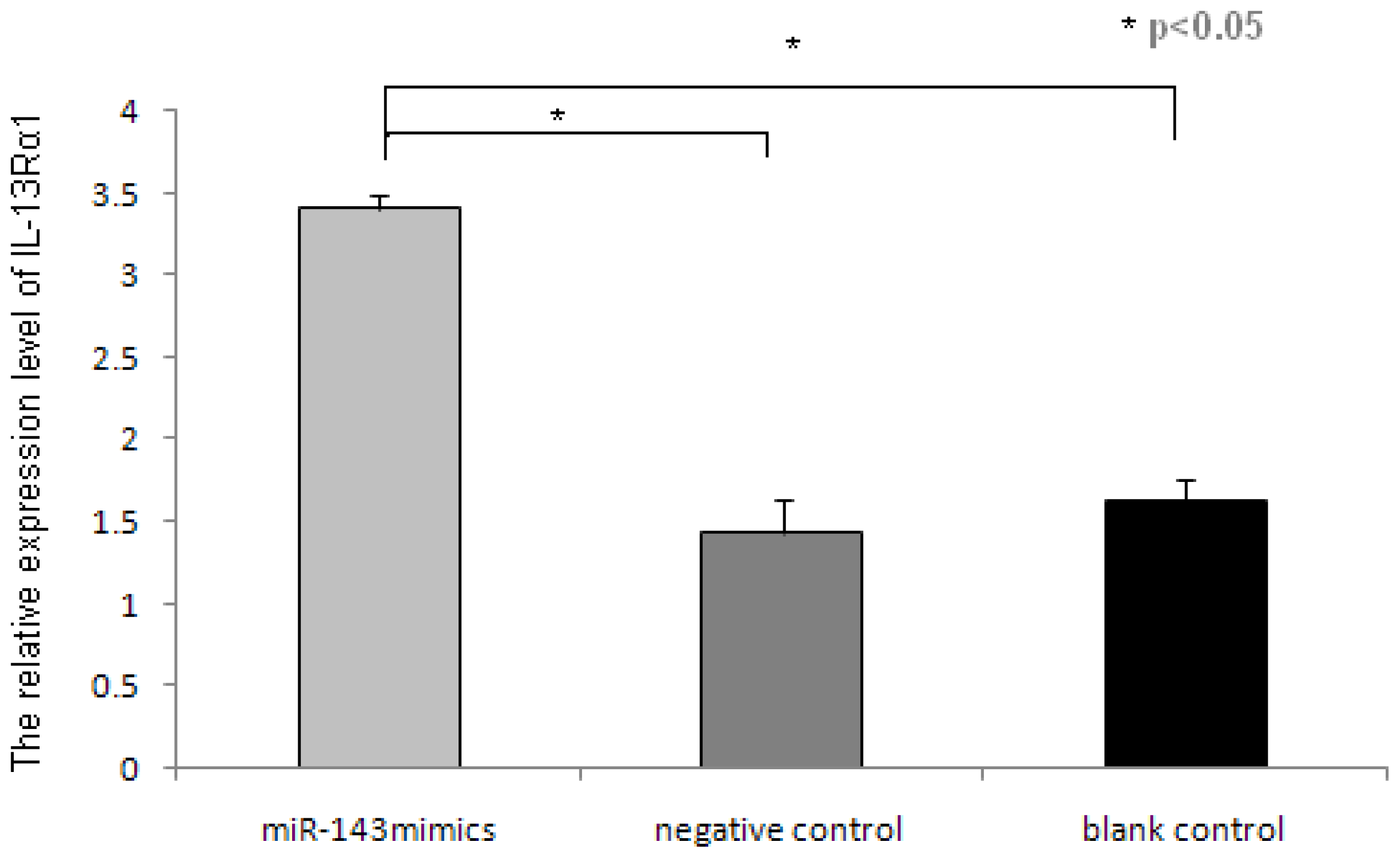
2.3. miR-143 Suppresses IL-13Rα1 Expression in HMC-1 Cells
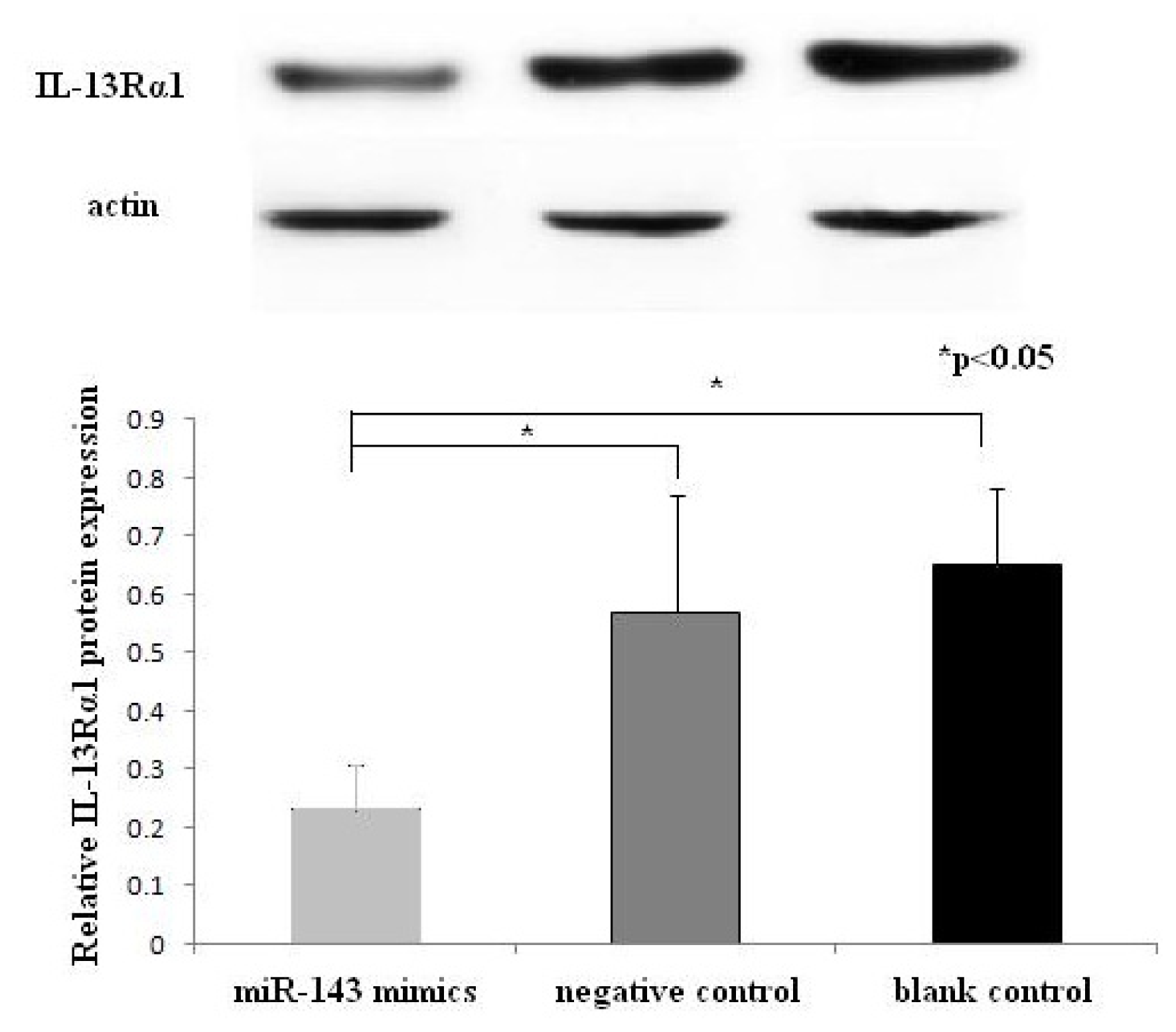
2.4. Western Blotting Results of IL-13Rα1 Downregulation in miR-143 Transfected HMC-1 Cells
3. Discussion
4. Experimental Section
4.1. Construction of miR-143 Target Sequence-Luciferase Reporter Plasmid
4.2. Lentivirus Production and Transfection
4.3. Cell Culture
4.4. Transfection and Luciferase Assays
4.5. RNA Isolation and Reverse Transcription and Polymerase Chain Reaction (RT-PCR) for miR-143-Target Gene Expression
4.6. Western Blotting Analysis
4.7. Statistical Analysis
5. Conclusions
Acknowledgments
Conflicts of Interest
References
- Meltzer, E.O.; Nathan, R.; Derebery, J.; Stang, P.E.; Campbell, U.B.; Yeh, W.S.; Corrao, M.; Stanford, R. Sleep, quality of life, and productivity impact of nasal symptoms in the United States: Findings from the burden of rhinitis in America survey. Allergy Asthma Proc 2009, 30, 244–254. [Google Scholar]
- Blaiss, M.S. Allergic rhinitis: Direct and indirect costs. Allergy Asthma Proc 2010, 31, 375–380. [Google Scholar]
- Lee, Y.; Ahn, C.; Han, J.; Choi, H.; Kim, J.; Yim, J.; Lee, J.; Provost, P.; Rådmark, O.; Kim, S.; et al. The nuclear RNase III Drosha initiates microRNA processing. Nature 2003, 425, 415–419. [Google Scholar]
- Ji, X.; Li, J.; Xian, X. microRNAs: Potential regulators of airway smooth muscle cell plasticity involved in asthma-induced airway remodeling. Asian Biomed 2013, 7, 3–14. [Google Scholar]
- Farh, K.K.; Grimson, A.; Jan, C.; Lewis, B.P.; Johnston, W.K.; Lim, L.P.; Burge, C.B.; Bartel, D.P. The widespread impact of mammalian microRNAs on mRNA repression and evolution. Science 2005, 310, 1817–1821. [Google Scholar]
- Yu, S.; Zhang, R.; Liu, G.; Yan, Z.; Hu, H.; Yu, S.; Zhang, J. Microarray analysis of differentially expressed microRNAs in allergic rhinitis. Am. J. Rhinol. Allergy 2011, 25, e242–e246. [Google Scholar]
- Oh, C.K.; Geba, G.P.; Molfino, N. Investigational therapeutics targeting the IL-4/IL-13/STAT-6 pathway for the treatment of asthma. Eur. Respir. Rev 2010, 19, 46–54. [Google Scholar]
- Zhang, X.; Graves, P.R.; Zeng, Y. Stable Argonaute2 overexpression differentially regulates microRNA production. Biochim. Biophys. Acta 2009, 1789, 153–159. [Google Scholar]
- Pfeifer, A.; Hofmann, A. Lentiviral transgenesis. Methods Mol. Biol 2009, 530, 391–405. [Google Scholar]
- Loewen, N.; Poeschla, E.M. Lentiviral vectors. Adv. Biochem. Eng. Biotechnol 2005, 99, 169–191. [Google Scholar]
- Scherr, M.; Battmer, K.; Ganser, A.; Eder, M. Modulation of gene expression by lentiviral-mediated delivery of small interfering RNA. Cell Cycle 2003, 2, 251–257. [Google Scholar]
- Elia, L.; Quintavalle, M.; Zhang, J.; Contu, R.; Cossu, L.; Latronico, M.V.; Peterson, K.L.; Indolfi, C.; Catalucci, D.; Chen, J.; et al. The knockout of miR-143 and -145 alters smooth muscle cell maintenance and vascular homeostasis in mice: correlates with human disease. Cell Death Differ 2009, 16, 1590–1598. [Google Scholar]
- Boettger, T.; Beetz, N.; Kostin, S.; Schneider, J.; Krüger, M.; Hein, L.; Braun, T. Acquisition of the contractile phenotype by murine arterial smooth muscle cells depends on the miR143/145 gene cluster. J. Clin. Invest 2009, 119, 2634–2647. [Google Scholar]
- Norata, G.D.; Pinna, C.; Zappella, F.; Elia, L.; Sala, A.; Condorelli, G.; Catapano, A.L. microRNA 143–145 deficiency impairs vascular function. Int. J. Immunopathol. Pharmacol 2012, 25, 467–474. [Google Scholar]
- Lai, V.K.; Ashraf, M.; Jiang, S.; Haider, K. microRNA-143 is a critical regulator of cell cycle activity in stem cells with co-overexpression of Akt and angiopoietin-1 via transcriptional regulation of Erk5/cyclin D1 signaling. Cell Cycle 2012, 11, 767–777. [Google Scholar]
- Ugras, S.; Brill, E.; Jacobsen, A.; Hafner, M.; Socci, N.D.; Decarolis, P.L.; Khanin, R.; O’Connor, R.; Mihailovic, A.; Taylor, B.S.; et al. Small RNA sequencing and functional characterization reveals microRNA-143 tumor suppressor activity in liposarcoma. Cancer Res 2011, 71, 5659–5669. [Google Scholar]
- Kelly-Welch, A.; Hanson, E.M.; Keegan, A.D. Interleukin-13 (IL-13) pathway. Sci. STKE 2005, 2005, cm8. [Google Scholar]
- Munitz, A.; Brandt, E.B.; Mingler, M.; Finkelman, F.D.; Rothenberg, M.E. Distinct roles for IL-13 and IL-4 via IL-13 receptor alpha1 and the type II IL-4 receptor in asthma pathogenesis. Proc. Natl. Acad. Sci. USA 2008, 105, 7240–7245. [Google Scholar]
- Guo, J.; Apiou, F.; Mellerin, M.P.; Lebeau, B.; Jacques, Y.; Minvielle, S. Chromosome mapping and expression of the human interleukin-13 receptor. Genomics 1997, 42, 141–145. [Google Scholar]
- Kaur, D.; Hollins, F.; Woodman, L.; Yang, W.; Monk, P.; May, R.; Bradding, P.; Brightling, C.E. Mast cells express IL-13R alpha 1: IL-13 promotes human lung mast cell proliferation and Fc epsilon RI expression. Allergy 2006, 61, 1047–1053. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Nuclei mapped to alignments | Nuclei mapped to sequence | Structure of predicted duplex | Probabilities | Free energies kcal/mol |
|---|---|---|---|---|
| 1397 | 1338 | _GAG__GCAG__G________CAUCUCA_: _CUC__UGUC__C________GUAGAGU_ | 0.30 | −24.1 |
© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Yu, S.; Zhang, R.; Zhu, C.; Cheng, J.; Wang, H.; Wu, J. MicroRNA-143 Downregulates Interleukin-13 Receptor Alpha1 in Human Mast Cells. Int. J. Mol. Sci. 2013, 14, 16958-16969. https://doi.org/10.3390/ijms140816958
Yu S, Zhang R, Zhu C, Cheng J, Wang H, Wu J. MicroRNA-143 Downregulates Interleukin-13 Receptor Alpha1 in Human Mast Cells. International Journal of Molecular Sciences. 2013; 14(8):16958-16969. https://doi.org/10.3390/ijms140816958
Chicago/Turabian StyleYu, Shaoqing, Ruxin Zhang, Chunshen Zhu, Jianqiu Cheng, Hong Wang, and Jing Wu. 2013. "MicroRNA-143 Downregulates Interleukin-13 Receptor Alpha1 in Human Mast Cells" International Journal of Molecular Sciences 14, no. 8: 16958-16969. https://doi.org/10.3390/ijms140816958
APA StyleYu, S., Zhang, R., Zhu, C., Cheng, J., Wang, H., & Wu, J. (2013). MicroRNA-143 Downregulates Interleukin-13 Receptor Alpha1 in Human Mast Cells. International Journal of Molecular Sciences, 14(8), 16958-16969. https://doi.org/10.3390/ijms140816958