The Characterisation of Pluripotent and Multipotent Stem Cells Using Fourier Transform Infrared Microspectroscopy

Abstract
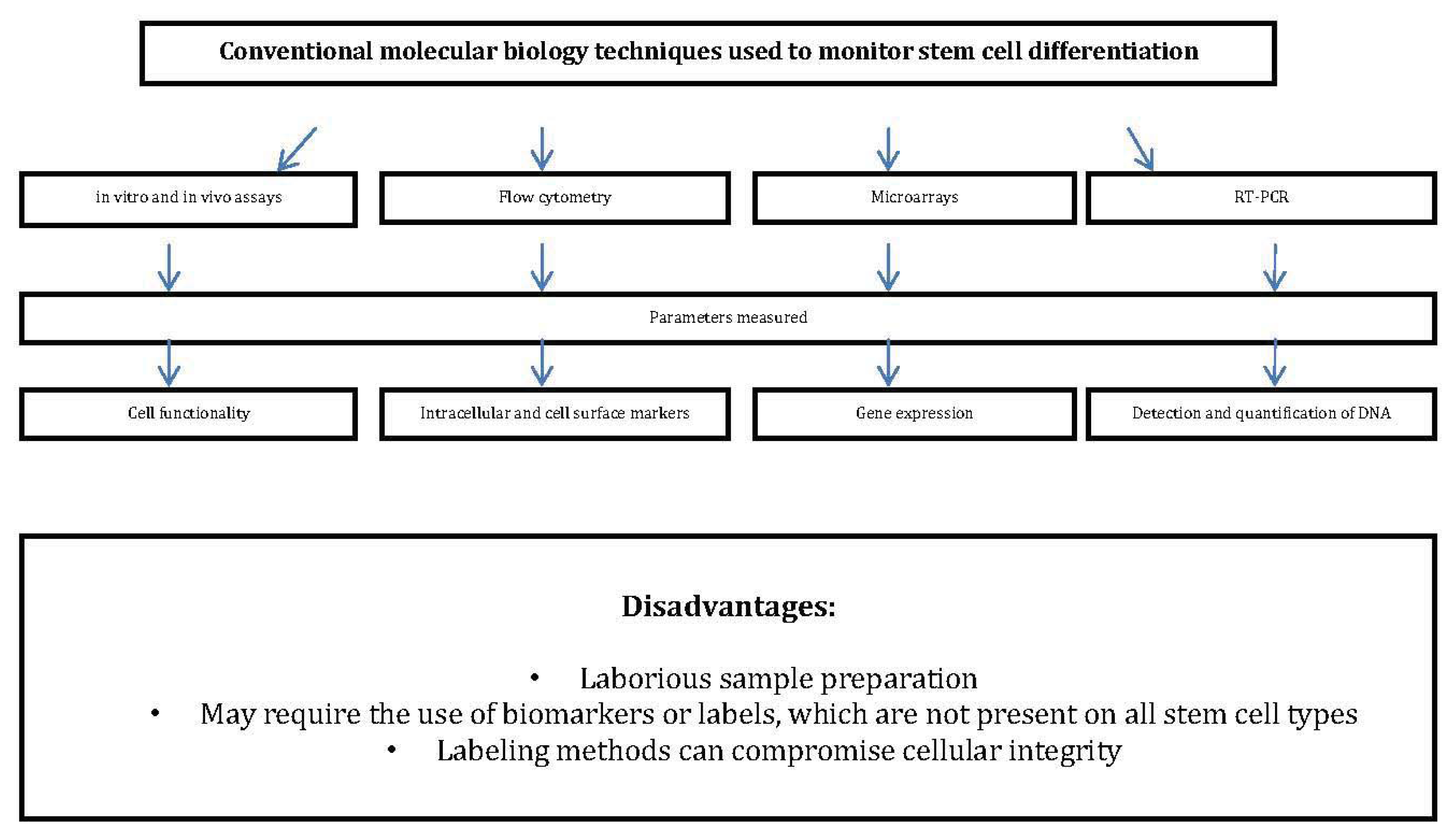
:1. Introduction
2. FTIR Microspectroscopy—A Concise Background
FTIR Microspectroscopy Instrumentation
3. Interpreting Features of the FTIR Spectrum
3.1. Principal Component Analysis (PCA)
3.2. Linear Discriminant Analysis (LDA)
3.3. Partial Least Squares Discriminant Analysis (PLS-DA)
3.4. Unsupervised Hierarchical Cluster Analysis (UHCA)
3.5. Artificial Neural Networks
4. FTIR Microspectroscopy Discriminates between Stem Cells of Different Potencies
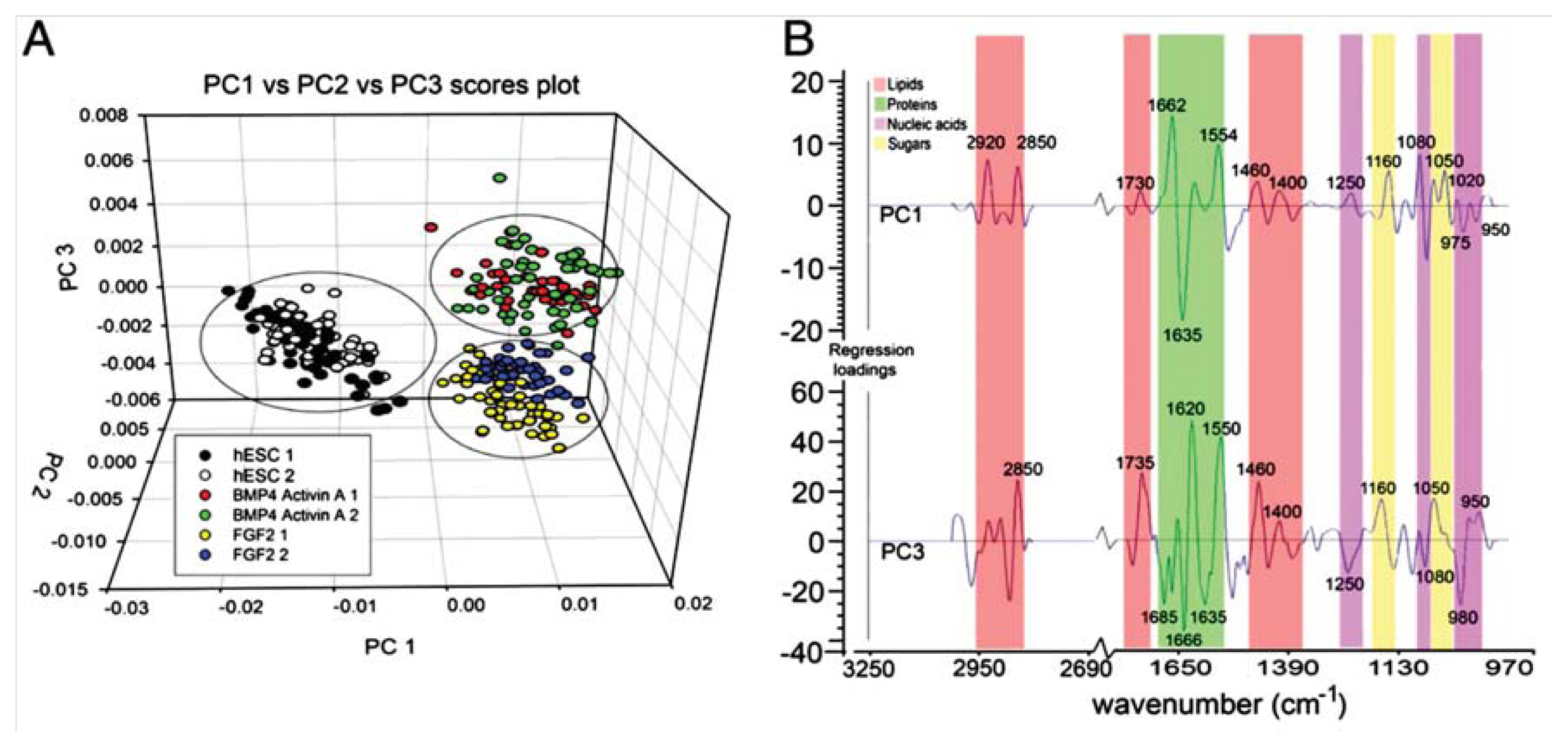
5. Pluripotent Stem Cell Applications—Embryonic Stem Cells
6. Pluripotent Stem Cell Applications—Induced Pluripotent Stem Cells
7. Multipotent Stem Cell Applications
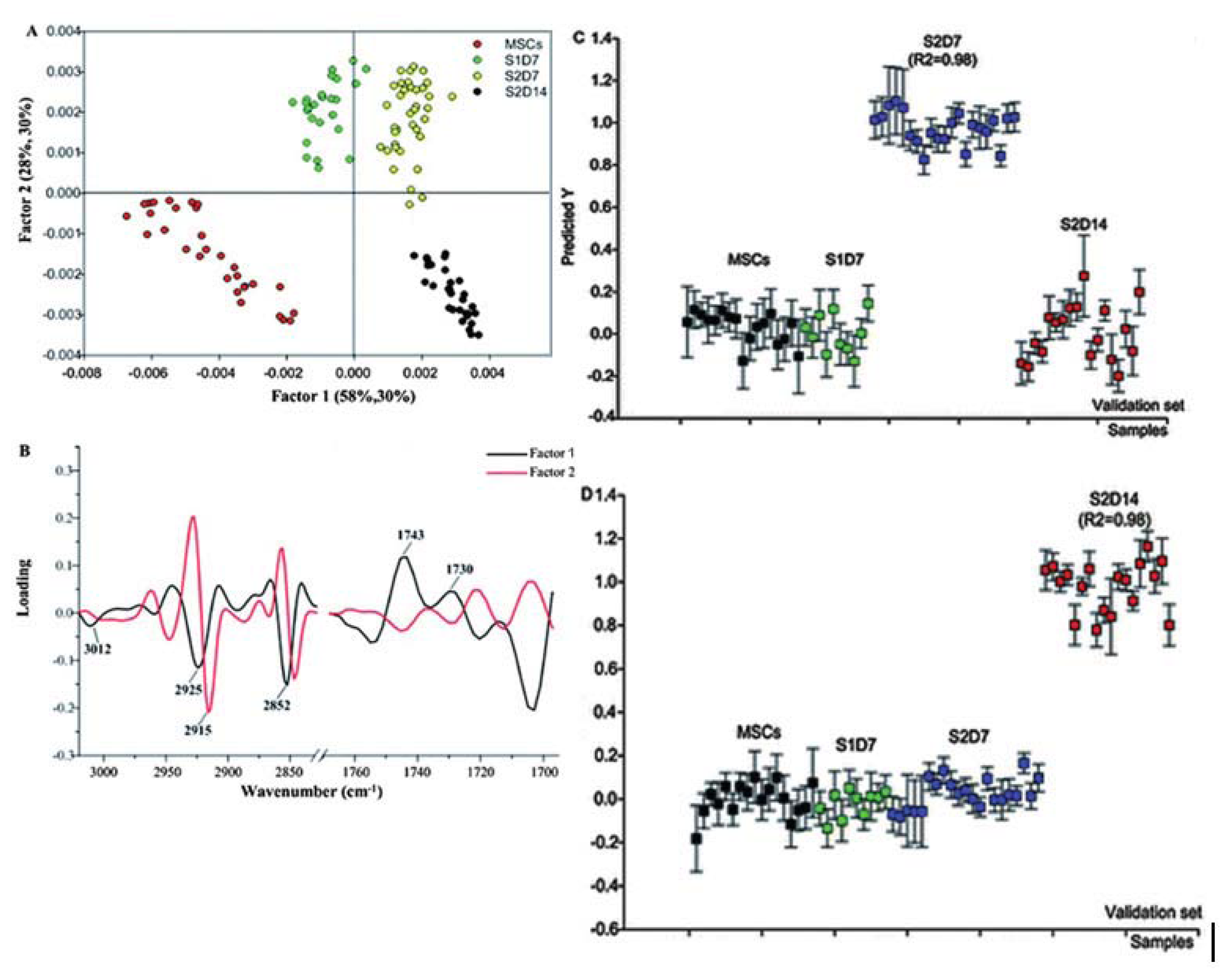
7.1. Mesenchymal Stem Cells
7.2. Corneal Stem Cells
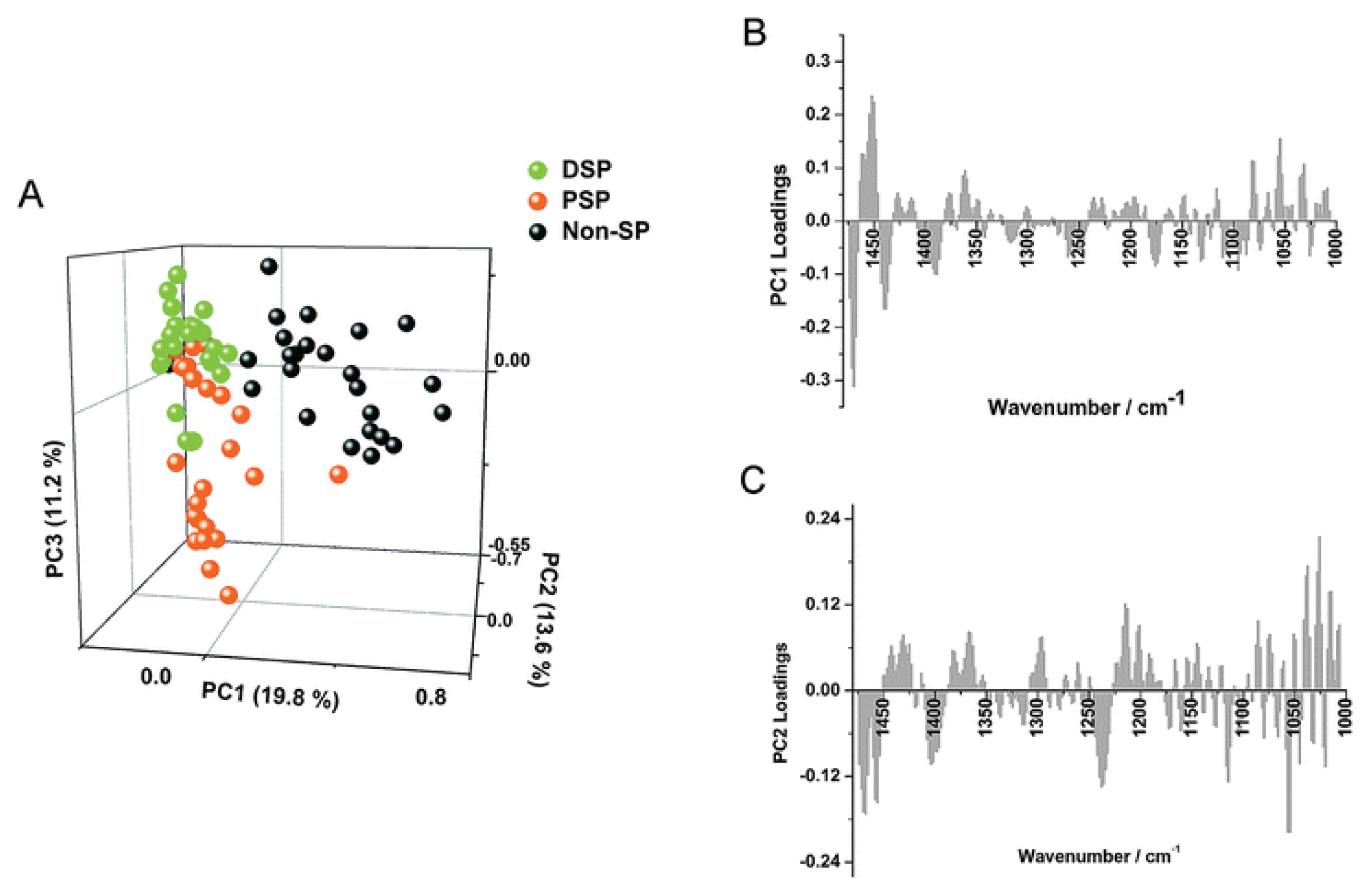
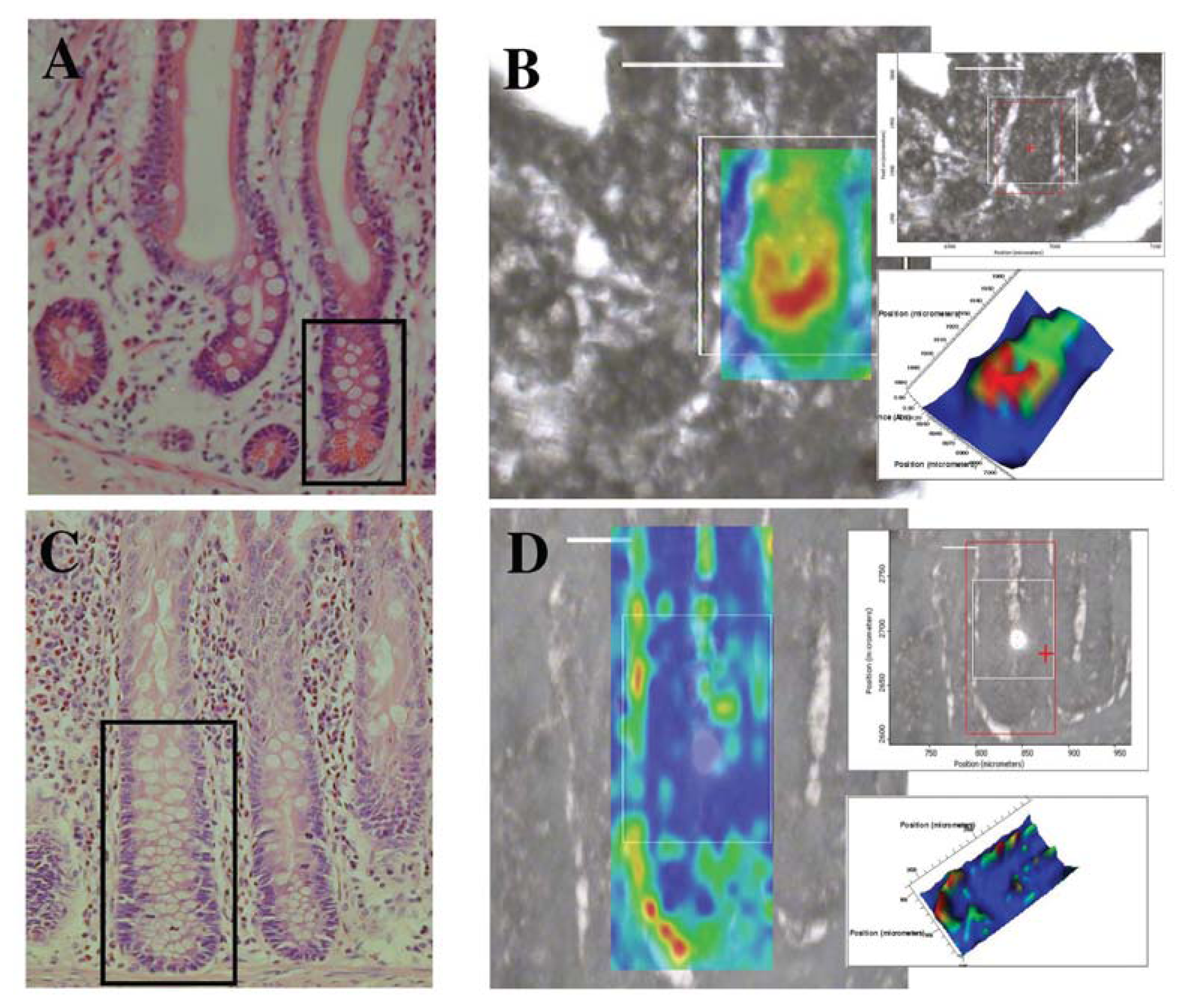
7.3. Gastrointestinal Crypt Stem Cells
7.4. Other Applications of FTIR Microspectroscopy to Stem Cell Research
7.4.1. Haematopoietic Stem Cells
7.4.2. Cancer Stem Cells
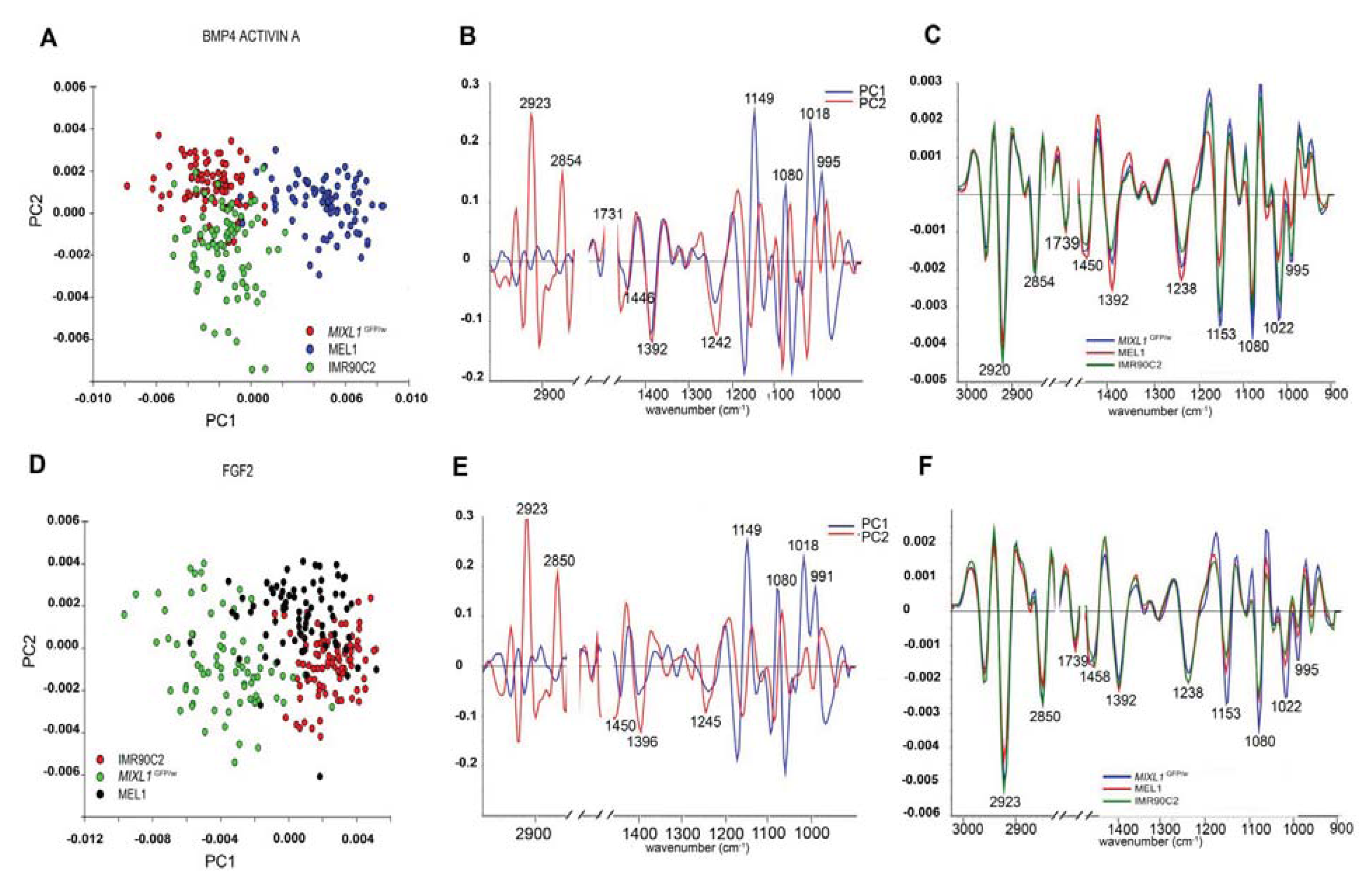
7.5. Unique Macromolecular Chemical Signature Differences Exist between Undifferentiated and Differentiated Stem Cells
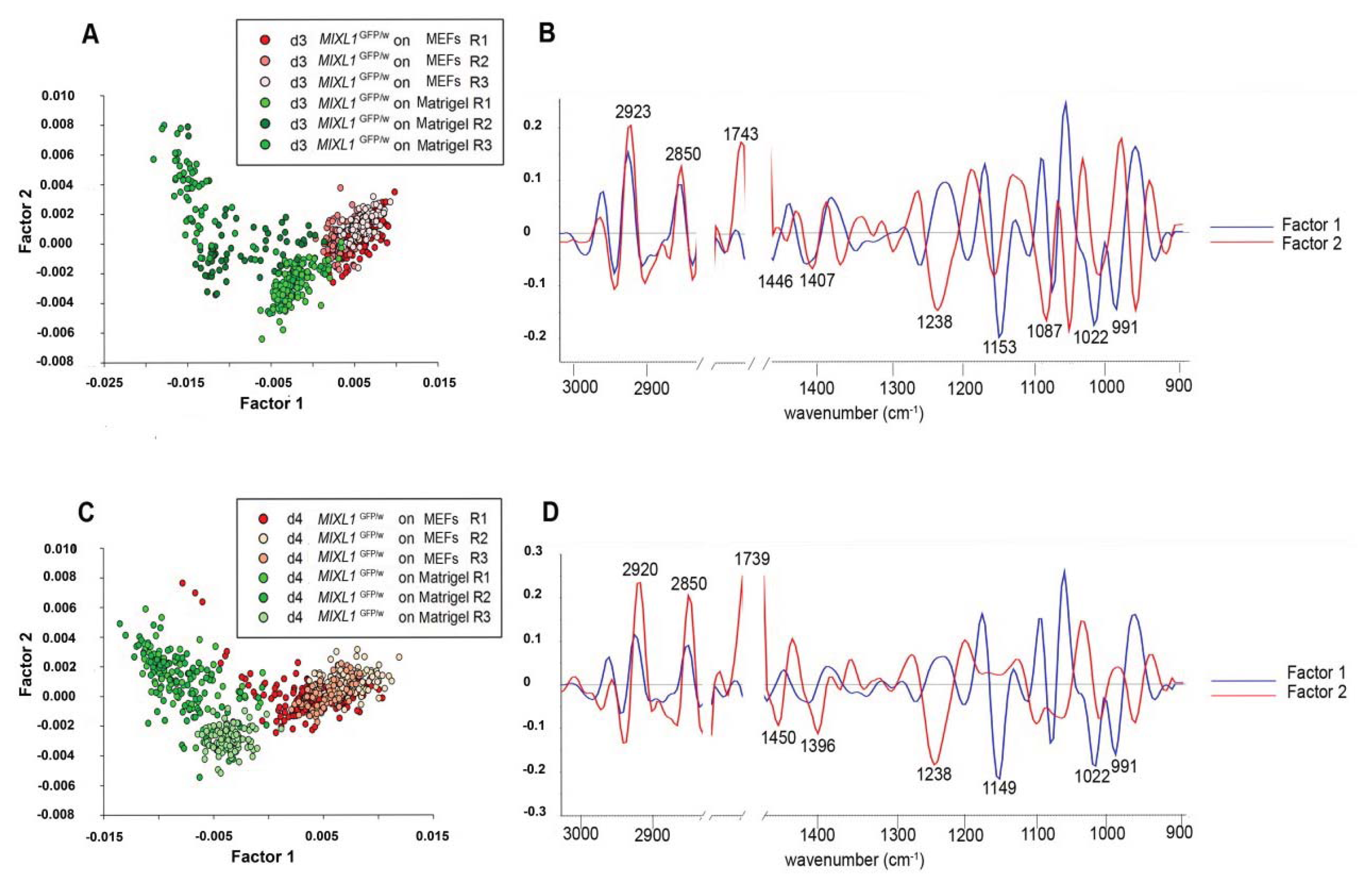
7.6. FTIR Spectroscopic Signatures of Stem Cells are Influenced by the Growth Environment
7.7. The Importance of Correlative Methodologies for the Interpretation of Stem Cell Spectral Signatures
8. Future Directions
Acknowledgments
Conflicts of Interest
References
- Thomson, J.A.; Itskovitz-Eldor, J.; Shapiro, S.S.; Waknitz, M.A.; Swiergiel, J.J.; Marshall, V.S.; Jones, J.M. Embryonic stem cell lines derived from human blastocysts. Science 1998, 282, 1145–1147. [Google Scholar]
- Takahashi, K.; Yamanaka, S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell 2006, 126, 663–676. [Google Scholar]
- Wagers, A.J.; Weissman, I.L. Plasticity of adult stem cells. Cell 2004, 116, 639–648. [Google Scholar]
- Griffith, L.G.; Naughton, G. Tissue engineering—Current challenges and expanding opportunities. Science 2002, 295, 1009–1014. [Google Scholar]
- Langer, R. Tissue engineering: Perspectives, challenges, and future directions. Tissue Eng 2007, 13, 1–2. [Google Scholar]
- Taupin, P. Derivation of embryonic stem cells for cellular therapy: Challenges and new strategies. Med. Sci. Monit. 2006, 12, RA75–RA78. [Google Scholar]
- Hoffman, L.M.; Carpenter, M.K. Characterization and culture of human embryonic stem cells. Nat. Biotechnol 2005, 23, 699–708. [Google Scholar]
- Walsh, M.J.; Hammiche, A.; Fellous, T.G.; Nicholson, J.M.; Cotte, M.; Susini, J.; Fullwood, N.J.; Martin-Hirsch, P.L.; Alison, M.R.; Martin, F.L. Tracking the cell hierarchy in the human intestine using biochemical signatures derived by mid-infrared microspectroscopy. Stem Cell Res 2009, 3, 15–27. [Google Scholar]
- German, M.J.; Pollock, H.M.; Zhao, B.; Tobin, M.J.; Hammiche, A.; Bentley, A.; Cooper, L.J.; Martin, F.L.; Fullwood, N.J. Characterization of putative stem cell populations in the cornea using synchrotron infrared microspectroscopy. Investig. Ophthalmol. Vis. Sci 2006, 47, 2417–2421. [Google Scholar]
- Downes, A.; Mouras, R.; Elfick, A. Optical spectroscopy for noninvasive monitoring of stem cell differentiation. J. Biomed. Biotechnol 2010, 2010, 1–11. [Google Scholar]
- Blout, E.R.; Mellors, R.C. Infrared spectra of tissues. Science 1949, 110, 137–138. [Google Scholar]
- Woernley, D.L. Infrared absorption curves for normal and neoplastic tissues and related biological substances. Cancer Res 1952, 12, 516–523. [Google Scholar]
- Mantsch, H.H.; Yang, P.W.; Casal, H.L. Infrared spectrometry of living systems: Current trends and perspectives. J. Mol. Struct 1986, 141, 237–242. [Google Scholar]
- Byler, D.M.; Susi, H. Examination of the secondary structure of proteins by deconvolved ftir spectra. Biopolymers 1986, 25, 469–487. [Google Scholar]
- Jackson, M.; Mantsch, H.H. The use and misuse of ftir spectroscopy in the determination of protein structure. Crit. Rev. Biochem. Mol. Biol 1995, 30, 95–120. [Google Scholar]
- Bassan, P.; Sachdeva, A.; Kohler, A.; Hughes, C.; Henderson, A.; Boyle, J.; Shanks, J.H.; Brown, M.; Clarke, N.W.; Gardner, P. Ftir microscopy of biological cells and tissue: Data analysis using resonant mie scattering (rmies) emsc algorithm. Analyst 2012, 137, 1370–1377. [Google Scholar]
- Vaccari, L.; Birarda, G.; Grenci, G.; Pacor, S.; Businaro, L. Synchrotron radiation infrared microspectroscopy of single living cells in microfluidic devices: Advantages, disadvantages and future perspectives. J. Phys.: Conf. Ser 2012, 359, 12007–12016. [Google Scholar]
- Lewis, E.N.; Treado, P.J.; Reeder, R.C.; Story, G.M.; Dowrey, A.E.; Marcott, C.; Levin, I.W. Fourier transform spectroscopic imaging using an infrared focal-plane array detector. Anal. Chem 1995, 67, 3377–3381. [Google Scholar]
- Nasse, M.J.; Bellehumeur, B.; Ratti, S.; Olivieri, C.; Buschke, D.; Squirrell, J.; Eliceiri, K.; Ogle, B.; Patterson, C.S.; Giordano, M.; et al. Opportunities for multiple-beam synchrotron-based mid-infrared imaging at ireni. Vib. Spectrosc 2012, 60, 10–15. [Google Scholar]
- Miller, L.M. Biomedical Applications of infrared Microspectroscopy Using Synchrotron Radiation. In Infrared and Raman Spectroscopic Imaging; WILEY-VCH Verlag GmbH: Weinheim, Germany, 2009. [Google Scholar] [CrossRef]
- Haibach, F.; Erlich, A.; Deutsch, E. Mid-infrared absorption spectroscopy using quantum cascade lasers. Available online: http://proceedings.spiedigitallibrary.org/proceeding.aspx?articleid=1350892 (accessed on 26 July 2013).
- Weida, M.J.; Yee, B. Quantum cascade laser-based replacement for FTIR microscopy. Available online: http://adsabs.harvard.edu/abs/2011SPIE.7902E..33W (accessed on 26 July 2013).
- Savitzky, A.; Golay, M.J. Smoothing and differentiation of data by simplified least squares procedures. Anal. Chem 1964, 36, 1627–1639. [Google Scholar]
- Martens, H.; Stark, E. Extended multiplicative signal correction and spectral interference subtraction: New preprocessing methods for near infrared spectroscopy. J. Pharm. Biomed. Anal 1991, 9, 625–635. [Google Scholar]
- Wold, S.; Esbensen, K.; Geladi, P. Principal component analysis. Chemom. Intell. Lab. Syst 1987, 2, 37–52. [Google Scholar]
- Hughes, C.; Liew, M.; Sachdeva, A.; Bassan, P.; Dumas, P.; Hart, C.A.; Brown, M.D.; Clarke, N.W.; Gardner, P. Sr-ftir spectroscopy of renal epithelial carcinoma side population cells displaying stem cell-like characteristics. Analyst 2010, 135, 3133–3141. [Google Scholar]
- Ye, D.; Tanthanuch, W.; Thumanu, K.; Sangmalee, A.; Parnpai, R.; Heraud, P. Discrimination of functional hepatocytes derived from mesenchymal stem cells using ftir microspectroscopy. Analyst 2012, 137, 4774–4784. [Google Scholar]
- Geladi, P.; Kowalski, B.R. Partial least-squares regression: A tutorial. Anal. Chim. Acta 1986, 185, 1–17. [Google Scholar]
- Tanthanuch, W.; Thumanu, K.; Lorthongpanich, C.; Parnpai, R.; Heraud, P. Neural differentiation of mouse embryonic stem cells studied by ftir spectroscopy. J. Mol. Struct 2010, 967, 189–195. [Google Scholar]
- Agatonovic-Kustrin, S.; Beresford, R. Basic concepts of artificial neural network (ann) modeling and its application in pharmaceutical research. J. Pharm. Biomed. Anal 2000, 22, 717–727. [Google Scholar]
- Pijanka, J.K.; Kumar, D.; Dale, T.; Yousef, I.; Parkes, G.; Untereiner, V.; Yang, Y.; Dumas, P.; Collins, D.; Manfait, M.; et al. Vibrational spectroscopy differentiates between multipotent and pluripotent stem cells. Analyst 2010, 135, 3126–3132. [Google Scholar]
- Heraud, P.; Ng, E.S.; Caine, S.; Yu, Q.C.; Hirst, C.; Mayberry, R.; Bruce, A.; Wood, B.R.; McNaughton, D.; Stanley, E.G.; et al. Fourier transform infrared microspectroscopy identifies early lineage commitment in differentiating human embryonic stem cells. Stem Cell Res 2010, 4, 140–147. [Google Scholar]
- Ami, D.; Neri, T.; Natalello, A.; Mereghetti, P.; Doglia, S.M.; Zanoni, M.; Zuccotti, M.; Garagna, S.; Redi, C.A. Embryonic stem cell differentiation studied by FT-IR spectroscopy. Biochim. Biophys. Acta (BBA) 2008, 1783, 98–106. [Google Scholar]
- Thumanu, K.; Tanthanuch, W.; Ye, D.; Sangmalee, A.; Lorthongpanich, C.; Parnpai, R.; Heraud, P. Spectroscopic signature of mouse embryonic stem cell-derived hepatocytes using synchrotron fourier transform infrared microspectroscopy. J. Biomed. Opt. 2011, 16. [Google Scholar] [CrossRef]
- Sandt, C.; Féraud, O.; Oudrhiri, N.; Bonnet, M.L.; Meunier, M.C.; Valogne, Y.; Bertrand, A.; Raphaël, M.; Griscelli, F.; Turhan, A.G.; et al. Identification of spectral modifications occurring during reprogramming of somatic cells. PLoS One 2012, 7, e30743. [Google Scholar]
- Cao, J.; Ng, E.S.; McNaughton, D.; Stanley, E.G.; Elefanty, A.G.; Tobin, M.J.; Heraud, P. Fourier transform infrared microspectroscopy reveals unique phenotypes for human embryonic and induced pluripotent stem cell lines and their progeny. J. Biophotonics 2013. [Google Scholar] [CrossRef]
- Kansiz, M.; Christian Schuster, K.; McNaughton, D.; Lendl, B. Sequential injection/mid-infrared spectroscopic analysis of an acetone-butanol-ethanol fermentation: Analyte cross-correlation effects. Spectrosc. Lett 2005, 38, 677–702. [Google Scholar]
- Sandt, C.; Frederick, J.; Dumas, P. Profiling pluripotent stem cells and organelles using synchrotron radiation infrared microspectroscopy. J. Biophotonics 2013, 6, 60–72. [Google Scholar]
- Krafft, C.; Salzer, R.; Seitz, S.; Ern, C.; Schieker, M. Differentiation of individual human mesenchymal stem cells probed by ftir microscopic imaging. Analyst 2007, 132, 647–653. [Google Scholar]
- Chonanant, C.; Jearanaikoon, N.; Leelayuwat, C.; Limpaiboon, T.; Tobin, M.J.; Jearanaikoon, P.; Heraud, P. Characterisation of chondrogenic differentiation of human mesenchymal stem cells using synchrotron ftir microspectroscopy. Analyst 2011, 136, 2542–2551. [Google Scholar]
- Faoláin, E.Ó.; Hunter, M.B.; Byrne, J.M.; Kelehan, P.; McNamara, M.; Byrne, H.J.; Lyng, F.M. A study examining the effects of tissue processing on human tissue sections using vibrational spectroscopy. Vib. Spectrosc. 2005, 38, 121–127. [Google Scholar]
- Nakamura, T.; Kelly, J.G.; Trevisan, J.; Cooper, L.J.; Bentley, A.J.; Carmichael, P.L.; Scott, A.D.; Cotte, M.; Susini, J.; Martin-Hirsch, P.L. Microspectroscopy of spectral biomarkers associated with human corneal stem cells. Mol. Vis 2010, 16, 359–368. [Google Scholar]
- Walsh, M.J.; Fellous, T.G.; Hammiche, A.; Lin, W.-R.; Fullwood, N.J.; Grude, O.; Bahrami, F.; Nicholson, J.M.; Cotte, M.; Susini, J.; et al. Fourier transform infrared microspectroscopy identifies symmetric po2—Modifications as a marker of the putative stem cell region of human intestinal crypts. Stem Cells 2008, 26, 108–118. [Google Scholar]
- Zelig, U.; Dror, Z.; Iskovich, S.; Zwielly, A.; Ben-Harush, M.; Nathan, I.; Mordechai, S.; Kapelushnik, J. Biochemical analysis and quantification of hematopoietic stem cells by infrared spectroscopy. J. Biomed. Opt. 2010, 15. [Google Scholar] [CrossRef]
- Whelan, D.R.; Bambery, K.R.; Heraud, P.; Tobin, M.J.; Diem, M.; McNaughton, D.; Wood, B.R. Monitoring the reversible b to a-like transition of DNA in eukaryotic cells using fourier transform infrared spectroscopy. Nucleic Acids Res 2011, 39, 5439–5448. [Google Scholar]
- Sjögren-Jansson, E.; Zetterström, M.; Moya, K.; Lindqvist, J.; Strehl, R.; Eriksson, P.S. Large-scale propagation of four undifferentiated human embryonic stem cell lines in a feeder-free culture system. Dev. Dyn 2005, 233, 1304–1314. [Google Scholar]
- Maitra, A.; Arking, D.E.; Shivapurkar, N.; Ikeda, M.; Stastny, V.; Kassauei, K.; Sui, G.; Cutler, D.J.; Liu, Y.; Brimble, S.N.; et al. Genomic alterations in cultured human embryonic stem cells. Nat Genet 2005, 37, 1099–1103. [Google Scholar]
- Akopian, V.; Andrews, P.; Beil, S.; Benvenisty, N.; Brehm, J.; Christie, M.; Ford, A.; Fox, V.; Gokhale, P.; Healy, L.; et al. Comparison of defined culture systems for feeder cell free propagation of human embryonic stem cells. In Vitro Cell. Dev.Biol 2010, 46, 247–258. [Google Scholar]
- Yanes, O.; Clark, J.; Wong, D.; Patti, G.; Sánchez-ruiz, A.; Benton, H.; Trauger, S.; Desponts, C.; Ding, S.; Siuzdak, G. Metabolic oxidation regulates embryonic stem cell differentiation. Nat. Chem. Biol 2010, 6, 411–417. [Google Scholar]
- Cao, J.; Ng, E.; McNaughton, D.; Stanley, E.; Elefanty, A.; Tobin, M.J.; Heraud, P. Fourier transform infrared microspectroscopy reveals that tissue culture conditions affect the macromolecular phenotype of human embryonic stem cells. Analyst 2013, 138, 4147–4160. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Band maxima (cm−1) | Band assignments |
|---|---|
| ~2962 | C–H stretching peak of lipids and proteins |
| ~2923 | C–H asymmetrical stretching of lipid groups and protein |
| ~2850 | C–H symmetrical stretching of lipid groups |
| ~1743 | C=O stretching of lipid esters |
| ~1685 | β-turn protein secondary structure |
| ~1654 | α-helical protein secondary structure |
| ~1635 | β-pleated sheet protein secondary structure |
| ~1554 | Overall protein absorbance |
| ~1458 | Methyl and methylene groups from lipids and protein |
| ~1396 | COO− stretching vibrations of amino acid side chains |
| ~1238 | P=O asymmetrical stretching of PO2 phosphodiester groups from phosphorylated molecules |
| ~1080 | P=O symmetrical stretching of PO2 phosphodiester groups from phosphorylated molecules, and glycogen |
| ~1153 | C–O vibrations from glycogen and other carbohydrates |
| ~1050 | C–O vibrations from glycogen and other carbohydrates |
| ~1022 | C–O vibrations from glycogen and other carbohydrates |
| ~995 | C–O stretch from RNA ribose chain and other carbohydrates |
| ~950 | C–C vibrations from nucleic acids |
© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Cao, J.; Ng, E.S.; McNaughton, D.; Stanley, E.G.; Elefanty, A.G.; Tobin, M.J.; Heraud, P. The Characterisation of Pluripotent and Multipotent Stem Cells Using Fourier Transform Infrared Microspectroscopy. Int. J. Mol. Sci. 2013, 14, 17453-17476. https://doi.org/10.3390/ijms140917453
Cao J, Ng ES, McNaughton D, Stanley EG, Elefanty AG, Tobin MJ, Heraud P. The Characterisation of Pluripotent and Multipotent Stem Cells Using Fourier Transform Infrared Microspectroscopy. International Journal of Molecular Sciences. 2013; 14(9):17453-17476. https://doi.org/10.3390/ijms140917453
Chicago/Turabian StyleCao, Julie, Elizabeth S. Ng, Donald McNaughton, Edouard G. Stanley, Andrew G. Elefanty, Mark J. Tobin, and Philip Heraud. 2013. "The Characterisation of Pluripotent and Multipotent Stem Cells Using Fourier Transform Infrared Microspectroscopy" International Journal of Molecular Sciences 14, no. 9: 17453-17476. https://doi.org/10.3390/ijms140917453
APA StyleCao, J., Ng, E. S., McNaughton, D., Stanley, E. G., Elefanty, A. G., Tobin, M. J., & Heraud, P. (2013). The Characterisation of Pluripotent and Multipotent Stem Cells Using Fourier Transform Infrared Microspectroscopy. International Journal of Molecular Sciences, 14(9), 17453-17476. https://doi.org/10.3390/ijms140917453