Leptin Activates RhoA/ROCK Pathway to Induce Cytoskeleton Remodeling in Nucleus Pulposus Cells

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
2.1. Leptin Activated the RhoA Signaling in NP Cells
2.2. Leptin Increased Phosphorylation of LIMK1 and Cofilin-2
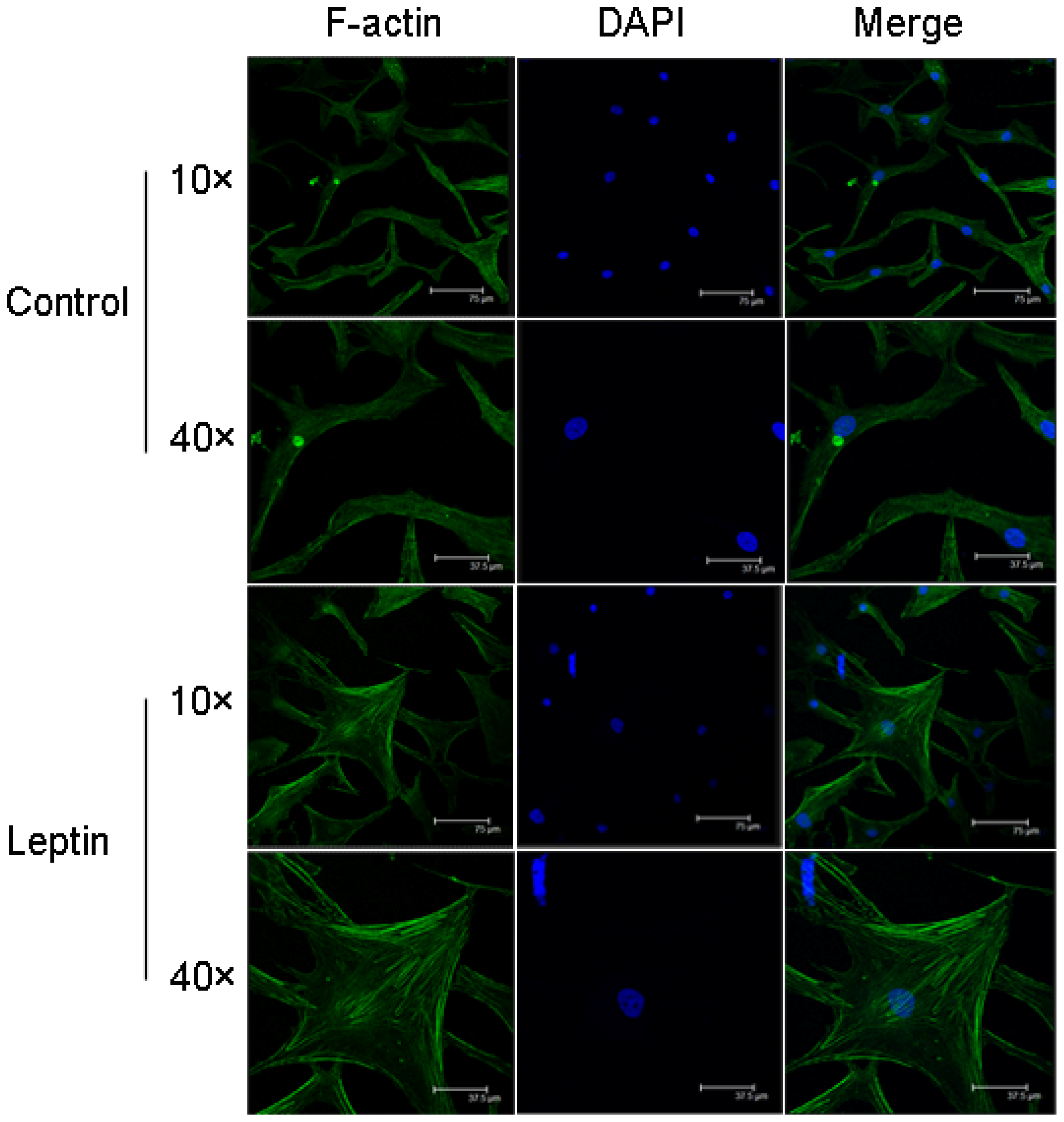
2.3. Leptin-Induced Cytoskeletal Reorganization in NP Cells
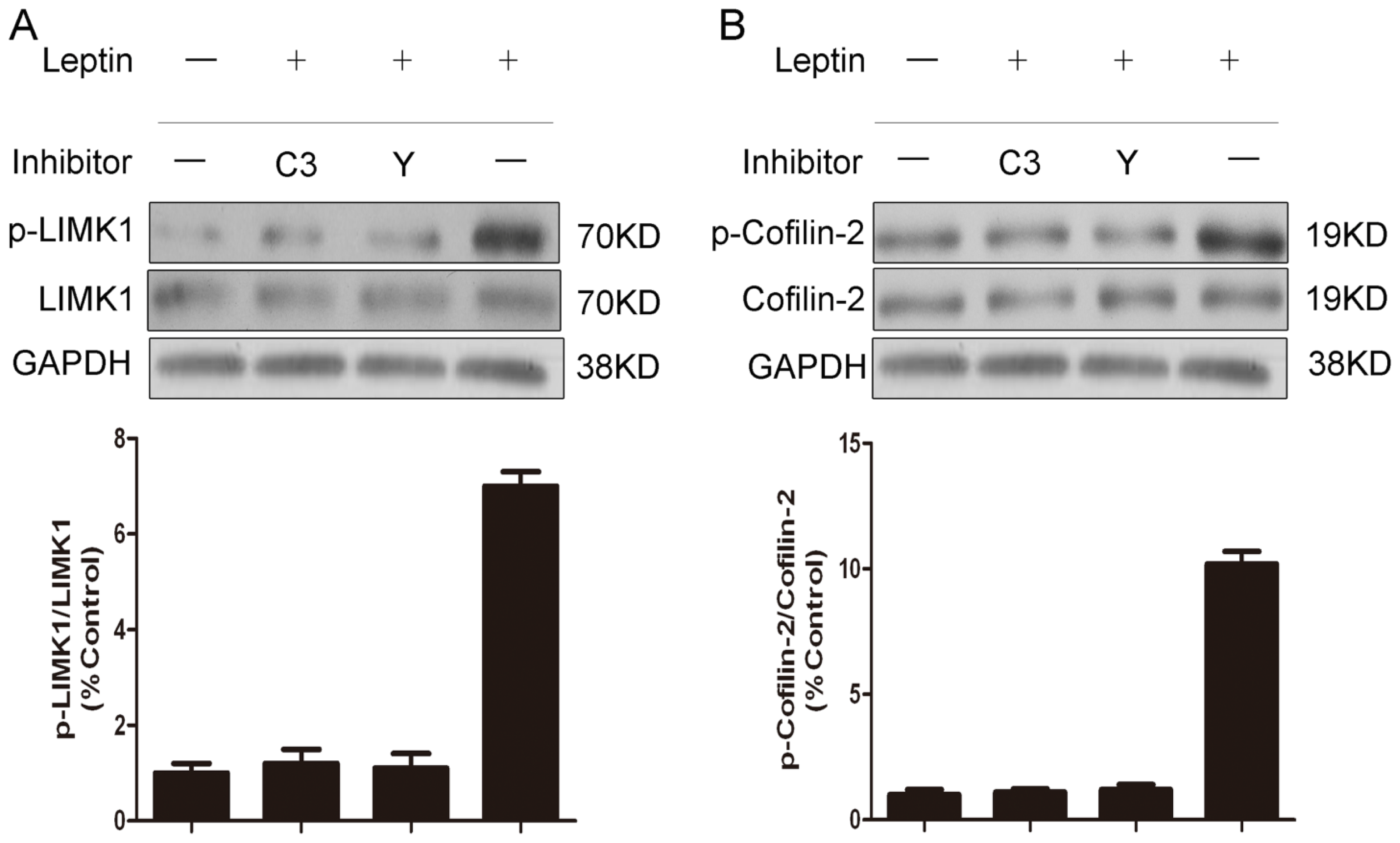
2.4. RhoA and ROCK Inhibition Prevented Leptin-Induced Phosphorylation of LIMK1 and Cofilin-2
2.5. Pharmacological Inhibition of RhoA/ROCK Prevented Leptin-Induced F-Actin Remodeling in NP Cells
2.6. Discussion
3. Experimental Section
3.1. Ethics Statement
3.2. Nucleus Pulposus Cell Isolation and Culture
3.3. Basic Structure of Raichu Probes
3.4. Nucleus Pulposus Cell Imageing and FRET Microscopy
3.5. Western Blotting Analysis
3.6. Immunofluorescence Microscopy
3.7. Statistical Analysis
4. Conclusions
Acknowledgments
Conflicts of Interest
References
- Haslam, D.W.; James, W.P. Obesity. Lancet 2005, 366, 1197–1209. [Google Scholar]
- Guh, D.P.; Zhang, W.; Bansback, N.; Amarsi, Z.; Birmingham, C.L.; Anis, A.H. The incidence of co-morbidities related to obesity and overweight: A systematic review and meta-analysis. BMC Public Health 2009, 9, 88. [Google Scholar]
- Samartzis, D.; Karppinen, J.; Mok, F.; Fong, D.Y.; Luk, K.D.; Cheung, K.M. A population-based study of juvenile disc degeneration and its association with overweight and obesity, low back pain, and diminished functional status. J. Bone Joint Surg. Am 2011, 93, 662–670. [Google Scholar]
- Bostman, O.M. Prevalence of obesity among patients admitted for elective orthopaedic surgery. Int. J. Obes. Relat. Metab. Disord 1994, 18, 709–713. [Google Scholar]
- L’Allemand, D.; Schmidt, S.; Rousson, V.; Brabant, G.; Gasser, T.; Gruters, A. Associations between body mass, leptin, IGF-I and circulating adrenal androgens in children with obesity and premature adrenarche. Eur. J. Endocrinol 2002, 146, 537–543. [Google Scholar]
- Halaas, J.L.; Gajiwala, K.S.; Maffei, M.; Cohen, S.L.; Chait, B.T.; Rabinowitz, D.; Lallone, R.L.; Burley, S.K.; Friedman, J.M. Weight-reducing effects of the plasma protein encoded by the obese gene. Science 1995, 269, 543–546. [Google Scholar]
- Denver, R.J.; Bonett, R.M.; Boorse, G.C. Evolution of leptin structure and function. Neuroendocrinology 2011, 94, 21–38. [Google Scholar]
- Ozenoglu, A.; Balci, H.; Ugurlu, S.; Caglar, E.; Uzun, H.; Sarkis, C.; Gunay, C.; Eker, E. The relationships of leptin, adiponectin levels and paraoxonase activity with metabolic and cardiovascular risk factors in females treated with psychiatric drugs. Clinics 2008, 63, 651–660. [Google Scholar]
- Gruber, H.E.; Ingram, J.A.; Hoelscher, G.L.; Hanley, E.N., Jr. Leptin expression by annulus cells in the human intervertebral disc. Spine J 2007, 7, 437–443. [Google Scholar]
- Zhao, C.Q.; Liu, D.; Li, H.; Jiang, L.S.; Dai, L.Y. Expression of leptin and its functional receptor on disc cells: Contribution to cell proliferation. Spine 2008, 33, E858–E864. [Google Scholar]
- Li, Z.; Shen, J.; Wu, W.K.; Yu, X.; Liang, J.; Qiu, G.; Liu, J. Leptin induces cyclin D1 expression and proliferation of human nucleus pulposus cells via JAK/STAT, PI3K/Akt and MEK/ERK pathways. PLoS One 2012, 7, e53176. [Google Scholar]
- Li, Z.; Shen, J.; Wu, W.K.; Yu, X.; Liang, J.; Qiu, G.; Liu, J. The role of leptin on the organization and expression of cytoskeleton elements in nucleus pulposus cells. J. Orthop. Res 2013, 31, 847–857. [Google Scholar]
- Li, S.; Duance, V.C.; Blain, E.J. Zonal variations in cytoskeletal element organization, mRNA and protein expression in the intervertebral disc. J. Anat 2008, 213, 725–732. [Google Scholar]
- Schram, K.; Ganguly, R.; No, E.K.; Fang, X.; Thong, F.S.; Sweeney, G. Regulation of MT1-MMP and MMP-2 by leptin in cardiac fibroblasts involves Rho/ROCK-dependent actin cytoskeletal reorganization and leads to enhanced cell migration. Endocrinology 2011, 152, 2037–2047. [Google Scholar]
- Liang, J.; Feng, J.; Wu, W.K.; Xiao, J.; Wu, Z.; Han, D.; Zhu, Y.; Qiu, G. Leptin-mediated cytoskeletal remodeling in chondrocytes occurs via the RhoA/ROCK pathway. J. Orthop. Res 2011, 29, 369–374. [Google Scholar]
- Zeidan, A.; Javadov, S.; Karmazyn, M. Essential role of Rho/ROCK-dependent processes and actin dynamics in mediating leptin-induced hypertrophy in rat neonatal ventricular myocytes. Cardiovasc. Res 2006, 72, 101–111. [Google Scholar]
- Thirone, A.C.; Speight, P.; Zulys, M.; Rotstein, O.D.; Szaszi, K.; Pedersen, S.F.; Kapus, A. Hyperosmotic stress induces Rho/Rho kinase/LIM kinase-mediated cofilin phosphorylation in tubular cells: Key role in the osmotically triggered F-actin response. Am. J. Physiol. Cell Physiol 2009, 296, C463–C475. [Google Scholar]
- Zeidan, A.; Paylor, B.; Steinhoff, K.J.; Javadov, S.; Rajapurohitam, V.; Chakrabarti, S.; Karmazyn, M. Actin cytoskeleton dynamics promotes leptin-induced vascular smooth muscle hypertrophy via RhoA/ROCK- and phosphatidylinositol 3-kinase/protein kinase B-dependent pathways. J. Pharmacol. Exp. Ther 2007, 322, 1110–1116. [Google Scholar]
- Pallu, S.; Francin, P.J.; Guillaume, C.; Gegout-Pottie, P.; Netter, P.; Mainard, D.; Terlain, B.; Presle, N. Obesity affects the chondrocyte responsiveness to leptin in patients with osteoarthritis. Arthritis Res. Ther 2010, 12, R112. [Google Scholar]
- Amano, M.; Nakayama, M.; Kaibuchi, K. Rho-kinase/ROCK: A key regulator of the cytoskeleton and cell polarity. Cytoskeleton 2010, 67, 545–554. [Google Scholar]
- Eichorst, J.P.; Lu, S.; Xu, J.; Wang, Y. Differential RhoA dynamics in migratory and stationary cells measured by FRET and automated image analysis. PLoS One 2008, 3, e4082. [Google Scholar]
- Chow, S.E.; Wang, J.S.; Lin, M.R.; Lee, C.L. Downregulation of p57kip(2) promotes cell invasion via LIMK/cofilin pathway in human nasopharyngeal carcinoma cells. J. Cell. Biochem 2011, 112, 3459–3468. [Google Scholar]
- Blain, E.J. Involvement of the cytoskeletal elements in articular cartilage homeostasis and pathology. Int. J. Exp. Pathol 2009, 90, 1–15. [Google Scholar]
- Li, S.; Jia, X.; Duance, V.C.; Blain, E.J. The effects of cyclic tensile strain on the organisation and expression of cytoskeletal elements in bovine intervertebral disc cells: An in vitro study. Eur. Cells Mater 2011, 21, 508–522. [Google Scholar]
- Li, S.; Duance, V.C.; Blain, E.J. F-actin cytoskeletal organization in intervertebral disc health and disease. Biochem. Soc. Trans 2007, 35, 683–685. [Google Scholar]
- Abbott, R.D.; Howe, A.K.; Langevin, H.M.; Iatridis, J.C. Live free or die: Stretch-induced apoptosis occurs when adaptive reorientation of annulus fibrosus cells is restricted. Biochem. Biophys. Res. Commun 2012, 421, 361–366. [Google Scholar]
- Nakamura, T.; Aoki, K.; Matsuda, M. Monitoring spatio-temporal regulation of Ras and Rho GTPase with GFP-based FRET probes. Methods 2005, 37, 146–153. [Google Scholar]
- Nakamura, T.; Kurokawa, K.; Kiyokawa, E.; Matsuda, M. Analysis of the spatiotemporal activation of rho GTPases using Raichu probes. Methods Enzymol 2006, 406, 315–332. [Google Scholar]






© 2014 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Li, Z.; Liang, J.; Wu, W.K.K.; Yu, X.; Yu, J.; Weng, X.; Shen, J. Leptin Activates RhoA/ROCK Pathway to Induce Cytoskeleton Remodeling in Nucleus Pulposus Cells. Int. J. Mol. Sci. 2014, 15, 1176-1188. https://doi.org/10.3390/ijms15011176
Li Z, Liang J, Wu WKK, Yu X, Yu J, Weng X, Shen J. Leptin Activates RhoA/ROCK Pathway to Induce Cytoskeleton Remodeling in Nucleus Pulposus Cells. International Journal of Molecular Sciences. 2014; 15(1):1176-1188. https://doi.org/10.3390/ijms15011176
Chicago/Turabian StyleLi, Zheng, Jinqian Liang, William Ka Kei Wu, Xin Yu, Jun Yu, Xisheng Weng, and Jianxiong Shen. 2014. "Leptin Activates RhoA/ROCK Pathway to Induce Cytoskeleton Remodeling in Nucleus Pulposus Cells" International Journal of Molecular Sciences 15, no. 1: 1176-1188. https://doi.org/10.3390/ijms15011176
APA StyleLi, Z., Liang, J., Wu, W. K. K., Yu, X., Yu, J., Weng, X., & Shen, J. (2014). Leptin Activates RhoA/ROCK Pathway to Induce Cytoskeleton Remodeling in Nucleus Pulposus Cells. International Journal of Molecular Sciences, 15(1), 1176-1188. https://doi.org/10.3390/ijms15011176