Effects of Melatonin on the Proliferation and Apoptosis of Sheep Granulosa Cells under Thermal Stress


Abstract
:1. Introduction
2. Results
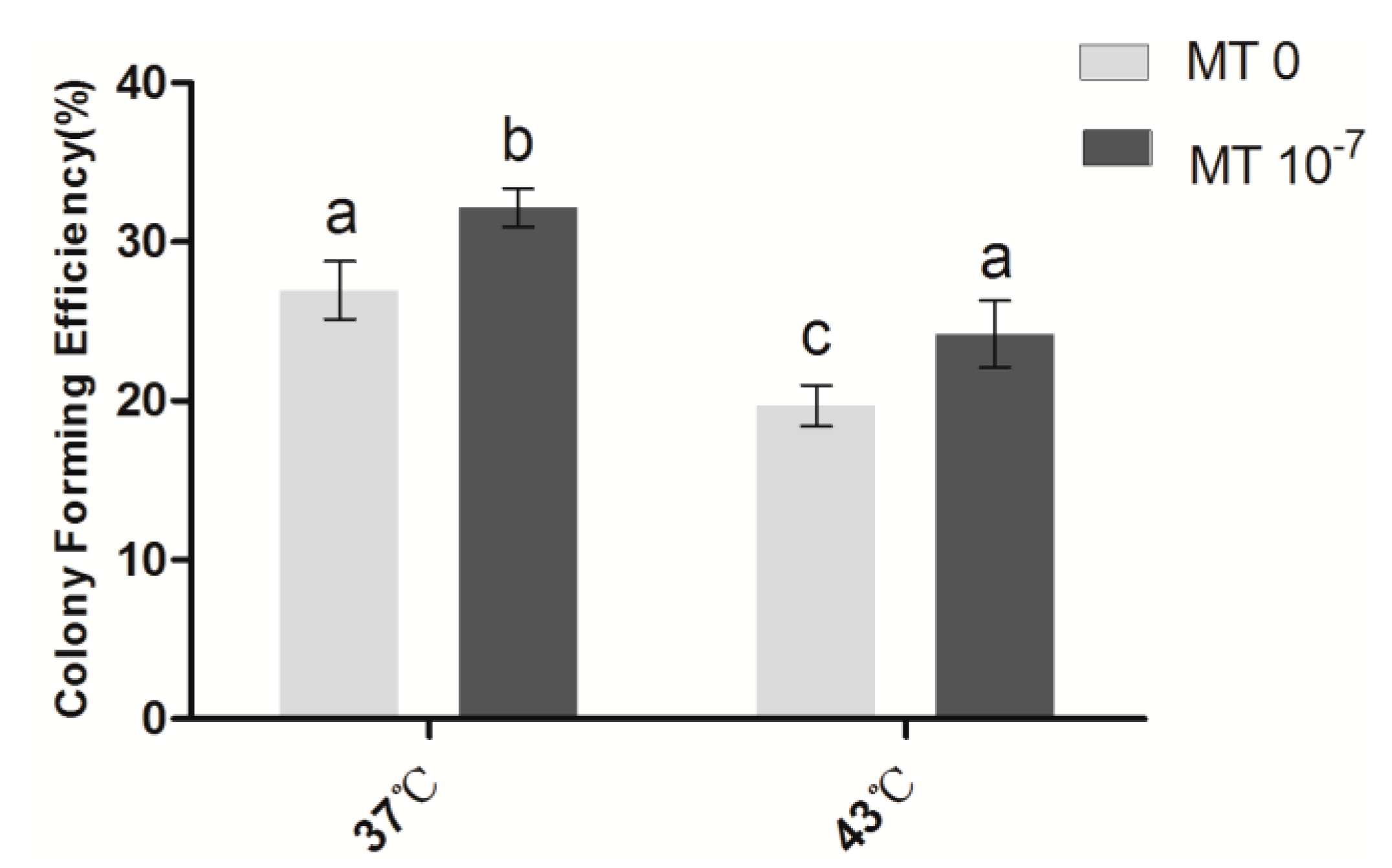
2.1. Effects of Melatonin on the Cloning Efficiency of Granulosa Cells

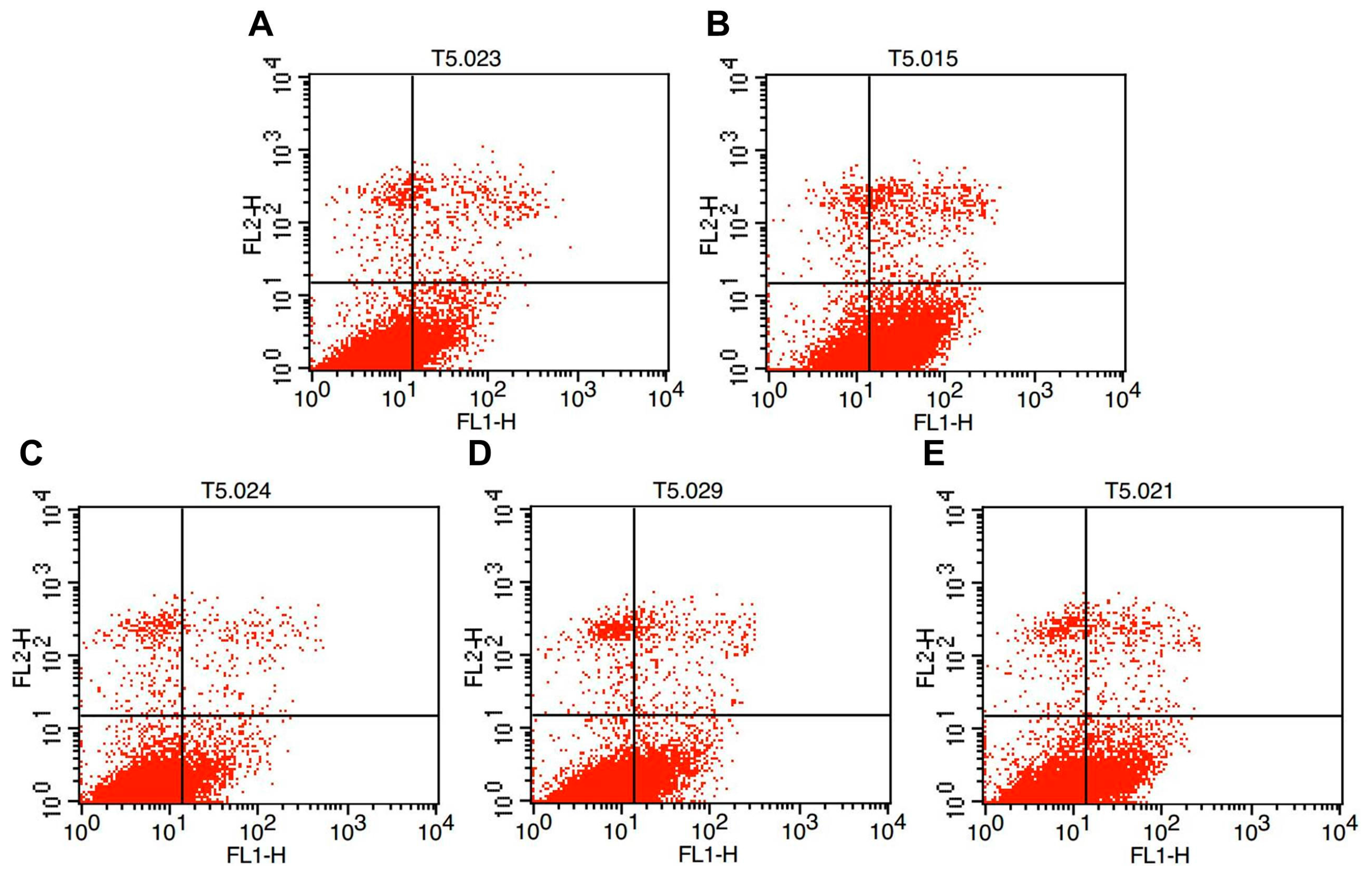
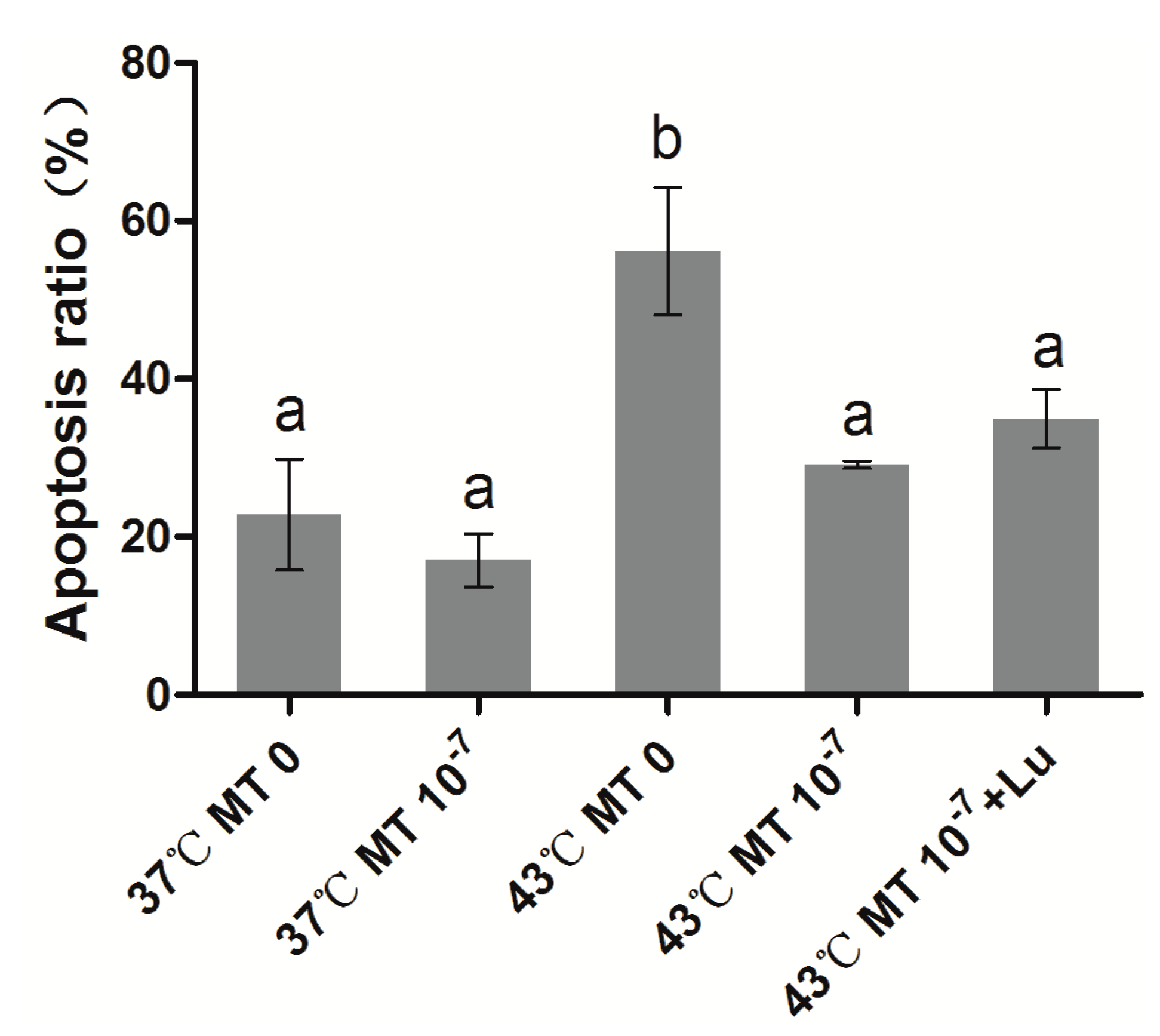
2.2. Effects of Melatonin on Granulosa Cell Apoptosis


2.3. Effects of Melatonin on the Expression of Apoptosis Genes in Sheep Granulosa Cells

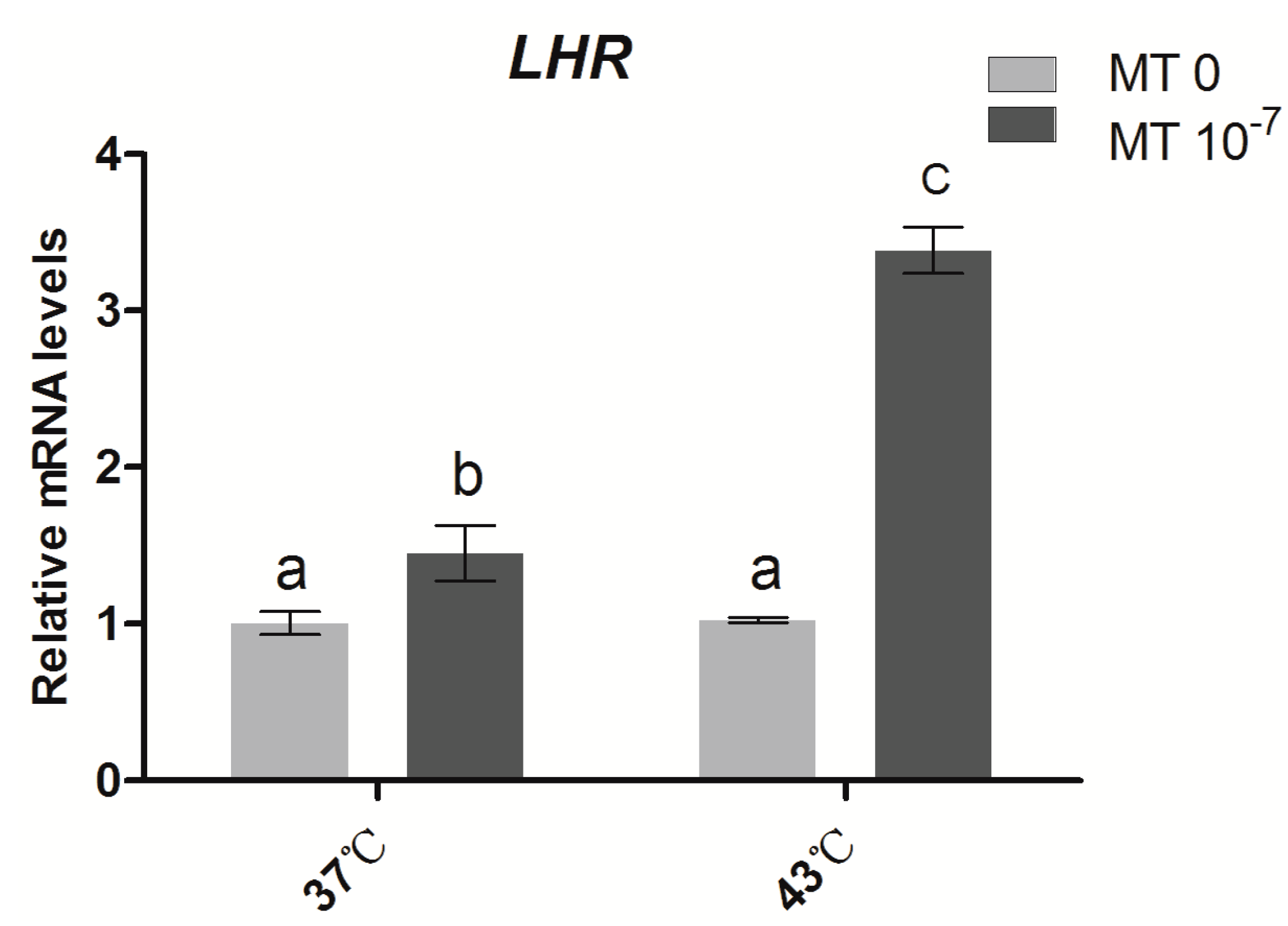
2.4. Effects of Melatonin on the Gene Expression of the Gonadotropin Receptor LHR in Sheep Granulosa Cells

3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Granular Cell Separation and Culture
4.3. Measurement of Colony Forming Efficiency
4.4. Flow Cytometric Analysis of Apoptotic Cells
4.5. RNA Isolation and Quantitative RT-PCR

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Accession Number | Primers | Sequence (5'–3') | Product Size (bp) |
|---|---|---|---|---|
| GAPDH | HM043737 | Forward | GTGTCTGTTGTGGATCTGACCTG | 162 |
| Reverse | AGAAGAGTGAGTGTCGCTGTTGAAGT | |||
| p53 | FJ855223 | Forward | GCACGACCATCCACTACAACTTC | 148 |
| Reverse | GGACAGGCACAAACACGCAC | |||
| Bcl-2 | DQ152929 | Forward | ACTTCGCCGAGATGTCCAG | 138 |
| Reverse | CGACACCTCCGAACTCAAAG | |||
| LHR | L36329 | Forward | TCTGCTCACCCAAGACACTCC | 247 |
| Reverse | GAGGCAATGAGTAGCAGGTAGAG |
4.6. Statistical Analysis
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Mustafi, S.B.; Chakraborty, P.K.; Dey, R.S.; Raha, S. Heat stress up-regulates chaperone heat shock protein 70 and antioxidant manganese superoxide dismutase through reactive oxygen species (ROS), p38MAPK, and Akt. Cell Stress Chaperones 2009, 14, 579–589. [Google Scholar]
- Farinati, F.; Piciocchi, M.; Lavezzo, E.; Bortolami, M.; Cardin, R. Oxidative stress and inducible nitric oxide synthase induction in carcinogenesis. Dig. Dis. 2010, 28, 579–584. [Google Scholar]
- Ziech, D.; Franco, R.; Pappa, A.; Panayiotidis, M.I. Reactive oxygen species (ROS)-induced genetic and epigenetic alterations in human carcinogenesis. Mut.Res.Fundam. Mol. Mechan. Mutagen. 2011, 711, 167–173. [Google Scholar]
- Hansen, P.J. Effects of heat stress on mammalian reproduction. Philos. Trans. R. Soc. B 2009, 364, 3341–3350. [Google Scholar]
- Reiter, R.J. The melatonin rhythm: Both a clock and a calendar. Experientia 1993, 49, 654–664. [Google Scholar]
- Reiter, R.J.; Rosales-Corral, S.; Coto-Montes, A.; Boga, J.A.; Tan, D.X.; Davis, J.M.; Konturek, P.C.; Konturek, S.J.; Brzozowski, T. The photoperiod, circadian regulation and chronodisruption: The requisite interplay between the suprachiasmatic muclei and the pineal and gut melatonin. J. Physiol. Pharmacol. 2011, 62, 269–274. [Google Scholar]
- Anisimov, V.N.; Popovich, I.G.; Zabezhinski, M.A.; Anisimov, S.V.; Vesnushkin, G.M.; Vinogradova, I.A. Melatonin as antioxidant, geroprotector and anticarcinogen. Biochim. Biophys. Acta Bioenerg. 2006, 1757, 573–589. [Google Scholar]
- Hardeland, R.; Pandi-Perumal, S. Melatonin, a potent agent in antioxidative defense: Actions as a natural food constituent, gastrointestinal factor, drug and prodrug. Nutr. Metab. 2005, 2, 22. [Google Scholar]
- Tengattini, S.; Reiter, R.J.; Tan, D.X.; Terron, M.P.; Rodella, L.F.; Rezzani, R. Cardiovascular diseases: Protective effects of melatonin. J. Pineal Res. 2008, 44, 16–25. [Google Scholar]
- Sirotkin, A.; Schaeffer, H. Direct regulation of mammalian reproductive organs by serotonin and melatonin. J. Endocrinol. 1997, 154, 1–5. [Google Scholar]
- Tamura, H.; Nakamura, Y.; Korkmaz, A.; Manchester, L.C.; Tan, D.X.; Sugino, N.; Reiter, R.J. Melatonin and the ovary: Physiological and pathophysiological implications. Fertil. Steril. 2009, 92, 328–343. [Google Scholar]
- Von Gall, C.; Stehle, J.H.; Weaver, D.R. Mammalian melatonin receptors: Molecular biology and signal transduction. Cell Tissue Res. 2002, 309, 151–162. [Google Scholar]
- Cutando, A.C.; Fernandez, J.A.; Valverde, A.L.; Santiago, S.A.; Cachaza, J.A.; Reiter, R.J. A new perspective in oral health: Potential importance and actions of melatonin receptors MT1, MT2, MT3, and RZR/ROR in the oral cavity. Arch. Oral Biol. 2011, 56, 944–950. [Google Scholar]
- Slominski, R.M.; Reiter, R.J.; Loutsevitch, N.S.; Ostrom, R.S.; Slominski, A.T. Melatonin membrane receptors in peripheral tissues: Distrbution and functions. Mol. Cell. Endocrinol. 2012, 351, 152–166. [Google Scholar]
- Slominski, A.T.; Kim, T.K.; Takeda, Y.; Janjetovic, Z.; Brozyna, A.A.; Skobowiat, C.; Wang, J.; Postlethwaite, A.; Li, W.; Tuckey, R.C.; et al. RORα and RORγ are expressed in human skin and serve as receptors for endogenously produced noncalcemic 20-hydroxy- and 20,23-dihydroxy vitamin D. Faseb. J. 2014, 28, 2775–2789. [Google Scholar]
- Reiter, R. Interactions of the pineal hormone melatonin with oxygen-centered free radicals: A brief review. Braz. J. Med. Biol. Res. 1993, 26, 1141–1155. [Google Scholar]
- Reiter, R.J.; Guerrero, J.M.; Escames, G.; Pappolla, M.A.; Acuña-Castroviejo, D. Prophylactic actions of melatonin in oxidative neurotoxicity. Ann. N. Y. Acad. Sci. 1997, 825, 70–78. [Google Scholar]
- Reiter, R.J.; Tan, D.X. Melatonin: A novel protective agent against oxidative injury of the ischemic/reperfused heart. Cardiovasc. Res. 2003, 58, 10–19. [Google Scholar]
- Carloni, S.; Perrone, S.; Buonocore, G.; Longini, M.; Proietti, F.; Balduini, W. Melatonin protects from the long-term consequences of a neonatal hypoxic-ischemic brain injury in rats. J. Pineal Res. 2008, 44, 157–164. [Google Scholar]
- Hung, M.W.; Tipoe, G.L.; Poon, A.M.S.; Reiter, R.J.; Fung, M.L. Protective effect of melatonin against hippocampal injury of rats with intermittent hypoxia. J. Pineal Res. 2008, 44, 214–221. [Google Scholar]
- Maldonado, M.D.; Murillo-Cabezas, F.; Calvo, J.R.; Lardone, P.J.; Tan, D.X.; Guerrero, J.M.; Reiter, R.J. Melatonin as pharmacologic support in burn patients: A proposed solution to thermal injury-related lymphocytopenia and oxidative damage. Crit. Care Med. 2007, 35, 1177–1185. [Google Scholar]
- Şener, G.; Şehirli, A.Ö.; Şatıroğlu, H.; Keyer-Uysal, M.Ç.; Yeğen, B. Melatonin improves oxidative organ damage in a rat model of thermal injury. Burns 2002, 28, 419–425. [Google Scholar]
- Karlidağ, T.; Yalçin, Ş.; Öztürk, A.; Üstündağ, B.; Gök, Ü.; Kaygusuz, İ.; Susaman, N. The role of free oxygen radicals in noise induced hearing loss: Effects of melatonin and methylprednisolone. Auris Nasus Larynx 2002, 29, 147–152. [Google Scholar]
- Bas, E.; Martinez-Soriano, F.; Láinez, J.M.; Marco, J. An experimental comparative study of dexamethasone, melatonin and tacrolimus in noise-induced hearing loss. Acta Oto-Laryngol. 2009, 129, 385–389. [Google Scholar]
- Seggie, J.; Campbell, L.; Brown, G.M.; Grota, L.J. Melatonin and N-acetylserotonin stress responses: Effects of type of stimulation and housing conditions. J. Pineal Res. 1985, 2, 39–49. [Google Scholar]
- Mahlberg, R.; Kunz, D.; Sutej, I.; Kühl, K.P.; Hellweg, R. Melatonin treatment of day-night rhythm disturbances and sundowning in Alzheimer disease: An open-label pilot study using actigraphy. J. Clin. Psychopharmacol. 2004, 24, 456–459. [Google Scholar]
- Blask, D.E. Melatonin, sleep disturbance and cancer risk. Sleep Med. Rev. 2009, 13, 257–264. [Google Scholar]
- Armstrong, S.; Redman, J. Melatonin: A chronobiotic with anti-aging properties? Med Hypotheses 1991, 34, 300–309. [Google Scholar]
- Reiter, R.J.; Tan, D.X.; Poeggeler, B.; Menendez-Pelaez, A.; Chen, L.D.; Saarela, S. Melatonin as a free radical scavenger: Implications for aging and age-related diseases. Ann. N. Y. Acad.Sci. 1994, 719, 1–12. [Google Scholar]
- Bizzarri, M.; Proietti, S.; Cucina, A.; Reiter, R.J. Molecular mechanisms of the pro-apoptotic actions of melatonin in cancer: A review. Expert Opin. Ther. Targets 2013, 17, 1483–1496. [Google Scholar]
- Sainz, R.; Mayo, J.; Rodriguez, C.; Tan, D.X.; Lopez-Burillo, S.; Reiter, R. Melatonin and cell death: Differential actions on apoptosis in normal and cancer cells. Cell Mol. Life Sci. 2003, 60, 1407–1426. [Google Scholar]
- Liu, L.; Xu, Y.; Reiter, R.J. Melatonin inhibits the proliferation of human osteosarcoma cell line MG-63. Bone 2013, 55, 432–438. [Google Scholar]
- Wang, J.; Xiao, X.; Zhang, Y.; Shi, D.; Chen, W.; Fu, L.; Liu, L.; Xie, F.; Kang, T.; Huang, W.; et al. Simultaneous modulation of COX-2, p300, Akt, and Apaf-1 signaling by melatonin to inhibit proliferation and induce apoptosis in breast cancer cells. J. Pineal Res. 2012, 53, 77–90. [Google Scholar]
- Roth, J.A.; Kim, B.-G.; Lin, W.-L.; Cho, M.-I. Melatonin promotes osteoblast differentiation and bone formation. J. Biol. Chem. 1999, 274, 22041–22047. [Google Scholar]
- Kong, X.; Li, X.; Cai, Z.; Yang, N.; Liu, Y.; Shu, J.; Pan, L.; Zuo, P. Melatonin regulates the viability and differentiation of rat midbrain neural stem cells. Cell Mol. Neurobiol. 2008, 28, 569–579. [Google Scholar]
- Li, R.; Albertini, D.F. The road to maturation: Somatic cell interaction and self-organization of the mammalian oocyte. Nat. Rev. Mol. Cell Biol. 2013, 14, 141–152. [Google Scholar]
- Manabe, N.; Goto, Y.; Matsuda-Minehata, F.; Inoue, N.; Maeda, A.; Sakamaki, K.; Miyano, T. Regulation mechanism of selective atresia in porcine follicles: Regulation of granulosa cell apoptosis during atresia. J. Reprod. Dev. 2004, 50, 493–514. [Google Scholar]
- Nakayama, M.; Manabe, N.; Nishihara, S.; Miyamoto, H. Species-specific differences in apoptotic cell localization in granulosa and theca interna cells during follicular atresia in porcine and bovine ovaries. J. Reprod. Dev. 2000, 46, 147–156. [Google Scholar]
- Krisher, R.L. In vivo and in vitro environmental effects on mammalian oocyte quality. Annu. Rev. Anim. Biosci. 2013, 1, 393–417. [Google Scholar]
- Agarwal, A.; Aponte-Mellado, A.; Premkumar, B.J.; Shaman, A.; Gupta, S. The effects of oxidative stress on female reproduction: A review. Reprod. Biol. Endocrinol. 2012, 10, 49. [Google Scholar]
- Agarwal, A.; Virk, G.; Ong, C.; du Plessis, S.S. Effect of oxidative stress on male reproduction. World J. Menʼs Health 2014, 32, 1–17. [Google Scholar]
- Ishikawa, S.; Machida, R.; Hiraga, K.; Hiradate, Y.; Suda, Y.; Tanemura, K. Hanging drop monoculture for selection of optimal antioxidants during in vitro maturation of porcine oocytes. Reprod. Domest. Anim. 2014, 49, e26–e30. [Google Scholar]
- Lian, H.Y.; Gao, Y.; Jiao, G.Z.; Sun, M.J.; Wu, X.F.; Wang, T.Y.; Li, H.; Tan, J.H. Antioxidant supplementation overcomes the deleterious effects of maternal restraint stress-induced oxidative stress on mouse oocytes. Reproduction 2013, 146, 559–568. [Google Scholar]
- Dong, Y.; Bai, Y.; Liu, G.; Wang, Z.; Cao, J.; Chen, Y.; Yang, H. The immunologic and antioxidant effects of L-phenylalanine on the uterine implantation of mice embryos during early pregnancy. Histol. Histopathol. 2014, 29, 1335–1242. [Google Scholar]
- Liu, M.; Yin, Y.; Ye, X.; Zeng, M.; Zhao, Q.; Keefe, D.L.; Liu, L. Resveratrol protects against age-associated infertility in mice. Hum. Reprod. 2013. [Google Scholar] [CrossRef]
- Chen, Z.G.; Luo, L.L.; Xu, J.J.; Zhuang, X.L.; Kong, X.X.; Fu, Y.C. Effects of plant polyphenols on ovarian follicular reserve in aging rats. Biochem. Cell Biol. 2010, 88, 737–745. [Google Scholar]
- Wang, F.; Tian, X.; Zhang, L.; He, C.; Ji, P.; Li, Y.; Tan, D.; Liu, G. Beneficial effect of resveratrol on bovine oocyte maturation and subsequent embryonic development after in vitro fertilization. Fertil. Steril. 2014, 101, 577–586. [Google Scholar]
- Sharma, S.; Ramesh, K.; Hyder, I.; Uniyal, S.; Yadav, V.; Panda, R.P.; Maurya, V.P.; Singh, G.; Kumar, P.; Mitra, A.; et al. Effect of melatonin administration on thyroid hormones, cortisol and expression profile of heat shock proteins in goats (Capra hircus) exposed to heat stress. Small Rumin. Res. 2013, 112, 216–223. [Google Scholar]
- Niu, Z.; Liu, F.; Yan, Q.; Li, W. Effects of different levels of vitamin E on growth performance and immune responses of broilers under heat stress. Poult. Sci. 2009, 88, 2101–2107. [Google Scholar]
- Xue, S.; Mu, D.; Dai, J.; Wang, Z.; Zhao, H.; Yuan, Y.; Peng, G. Preventive effect of glutamine combined with vitamin C on intestinal endotoxemia induced by heat stress in rats. J. Third Milit. Med. Univ. 2011, 17, 1779–1782. (In Chinese) [Google Scholar]
- Garcia-Ispierto, I.; Abdelfatah, A.; López-Gatius, F. Melatonin treatment at dry-off improves reproductive performance postpartum in high-producing dairy cows under heat stress conditions. Reprod. Domest. Anim. 2013, 28, 577–583. [Google Scholar]
- Wang, H.; Gao, G. Effect of hyperthermia on ovary antioxidant capacity and granulosa cells apoptosis of mouse. Heilongjiang Anim. Sci. Vet. Med. 2012, 11, 154–157. (In Chinese) [Google Scholar]
- Cebrian-Serrano, A.; Salvador, I.; Raga, E.; Dinnyes, A.; Silvestre, M. Beneficial effect of melatonin on blastocyst in vitro production from heat-stressed bovine oocytes. Reprod. Domest. Anim. 2013, 48, 738–746. [Google Scholar]
- Gao, C.; Han, H.B.; Tian, X.Z.; Tan, D.X.; Wang, L.; Zhou, G.B.; Zhu, S.E.; Liu, G.S. Melatonin promotes embryonic development and reduces reactive oxygen species in vitrified mouse 2-cell embryos. J. Pineal Res. 2012, 52, 305–311. [Google Scholar]
- Wang, F.; Tian, X.; Zhang, L.; Tan, D.; Reiter, R.J.; Liu, G. Melatonin promotes the in vitro development of pronuclear embryos and increases the efficiency of blastocyst implantation in murine. J. Pineal Res. 2013, 55, 267–274. [Google Scholar]
- Wang, F.; Tian, X.; Zhang, L.; Gao, C.; He, C.; Fu, Y.; Ji, P.Y.; Li, Y.; Liu, G.S. Beneficial effects of melatonin on in vitro bovine embryonic development are mediated by melatonin receptor 1. J. Pineal Res. 2014, 56, 333–342. [Google Scholar]
- Shi, J.M.; Tian, X.Z.; Zhou, G.B.; Wang, L.; Gao, C.; Zhu, S.E.; Zeng, S.-M.; Tian, J.-H.; Liu, G.-S. Melatonin exists in porcine follicular fluid and improves in vitro maturation and parthenogenetic development of porcine oocytes. J. Pineal Res. 2009, 47, 318–323. [Google Scholar]
- Jiang, X.; Wang, X. Cytochrome c promotes caspase-9 activation by inducing nucleotide binding to Apaf-1. J. Biol. Chem. 2000, 275, 31199–31203. [Google Scholar]
- Ratts, V.; Flaws, J.; Kolp, R.; Sorenson, C.; Tilly, J. Ablation of bcl-2 gene expression decreases the numbers of oocytes and primordial follicles established in the post-natal female mouse gonad. Endocrinology 1995, 136, 3665–3668. [Google Scholar]
- Hsu, S.Y.; Lai, R.; Finegold, M.; Hsueh, A. Targeted overexpression of Bcl-2 in ovaries of transgenic mice leads to decreased follicle apoptosis, enhanced folliculogenesis, and increased germ cell tumorigenesis. Endocrinology 1996, 137, 4837–4843. [Google Scholar]
- Baydas, G.; Reiter, R.; Akbulut, M.; Tuzcu, M.; Tamer, S. Melatonin inhibits neural apoptosis induced by homocysteine in hippocampus of rats via inhibition of cytochrome c translocation and caspase-3 activation and by regulating pro-and anti-apoptotic protein levels. Neuroscience 2005, 135, 879–886. [Google Scholar]
- Guha, M.; Maity, P.; Choubey, V.; Mitra, K.; Reiter, R.J.; Bandyopadhyay, U. Melatonin inhibits free radical-mediated mitochondrial-dependent hepatocyte apoptosis and liver damage induced during malarial infection. J. Pineal Res. 2007, 43, 372–381. [Google Scholar]
- Janjetovic, Z.; Zachary, P.; Nahmias, H.S.; Jarrett, S.G.; Kim, T.K.; Reiter, R.J.; Slominski, A.T. Melatonin and its metabolites ameliorate ultraviolet B-induced damage in human epidermal keratinocytes. J. Pineal Res. 2014, 57, 90–102. [Google Scholar]
- Sultana, H.; Kigawa, J.; Kanamori, Y.; Itamochi, H.; Oishi, T.; Sato, S.; Ohwada, M.; Suzuki, M.; Terakawa, N. Chemosensitivity and p53–Bax pathway-mediated apoptosis in patients with uterine cervical cancer. Ann. Oncol. 2003, 14, 214–219. [Google Scholar]
- Toshiyuki, M.; Reed, J.C. Tumor suppressor p53 is a direct transcriptional activator of the human bax gene. Cell 1995, 80, 293–299. [Google Scholar]
- Oh, S.H.; Nan, J.X.; Sohn, D.H.; Kim, Y.C.; Lee, B.H. Salvia miltiorrhiza inhibits biliary obstruction-induced hepatocyte apoptosis by cytoplasmic sequestration of p53. Toxicol. Appl. Pharmacol. 2002, 182, 27–33. [Google Scholar]
- Amaral, J.D.; Xavier, J.M.; Steer, C.J.; Rodrigues, C. The role of p53 in apoptosis. Discov. Med. 2010, 9, 145–152. [Google Scholar]
- Amaral, J.D.; Xavier, J.M.; Steer, C.J.; Rodrigues, C.M. Targeting the p53 pathway of apoptosis. Curr. Pharm. Des. 2010, 16, 2493–2503. [Google Scholar]
- Hsyeh, A.J.; Adashi, E.; Jones, P.B.; Welsh, J.R. Hormonal regulation of the differentiation of cultured ovarian granulosa cells. Endocrine Rev. 1984, 5, 76–127. [Google Scholar]
- Richards, J.S. Maturation of ovarian follicles: Actions and interactions of pituitary and ovarian hormones on follicular cell differentiation. Physiol. Rev. 1980, 60, 51–89. [Google Scholar]
- Jia, X.C.; Hhuen, J. Homologous regulation of hormone receptors: Luteinizing hormone increases its own receptors in cultured rat granulosa cells. Endocrinology 1984, 115, 2433–2439. [Google Scholar]
- Yao, N.; Yang, B.Q.; Liu, Y.; Tan, X.Y.; Lu, C.L.; Yuan, X.H.; Ma, X. Follicle-stimulating hormone regulation of microRNA expression on progesterone production in cultured rat granulosa cells. Endocrine 2010, 38, 158–166. [Google Scholar]
- Maizels, E.T.; Cottom, J.; Jones, J.C.; Hunzicker-Dunn, M. Follicle stimulating hormone (FSH) activates the p38 mitogen-activated protein kinase pathway, inducing small heat shock protein phosphorylation and cell rounding in immature rat ovarian granulosa cells. Endocrinology 1998, 139, 3353–3356. [Google Scholar]
- Boone, D.L.; Carnegie, J.A.; Rippstein, P.U.; Tsang, B.K. Induction of apoptosis in equine chorionic gonadotropin (eCG)-primed rat ovaries by anti-eCG antibody. Biol. Reprod. 1997, 57, 420–427. [Google Scholar]
- Webb, R.; Nicholas, B.; Gong, J.G.; Campbell, B.K.; Gutierrez, C.G.; Garverick, H.A.; Armstrong, D.G. Mechanisms regulating follicular development and selection of the dominant follicle. Reprod. Suppl. 2002, 61, 71–90. [Google Scholar]
- Woo, M.M.; Tai, C.J.; Kang, S.K.; Nathwani, P.S.; Pang, S.F.; Leung, P.C. Direct action of melatonin in human granulosa-luteal cells. J. Clin. Endocrinol. Metab. 2001, 86, 4789–4797. [Google Scholar]
- Lombardo, F.; Gioacchini, G.; Fabbrocini, A.; Candelma, M.; DʼAdamo, R.; Giorgini, E.; Carnevali, O. Melatonin-mediated effects on killifish reproductive axis. Comp. Biochem. Physiol. A 2014, 172, 31–38. [Google Scholar]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fu, Y.; He, C.-J.; Ji, P.-Y.; Zhuo, Z.-Y.; Tian, X.-Z.; Wang, F.; Tan, D.-X.; Liu, G.-S. Effects of Melatonin on the Proliferation and Apoptosis of Sheep Granulosa Cells under Thermal Stress. Int. J. Mol. Sci. 2014, 15, 21090-21104. https://doi.org/10.3390/ijms151121090
Fu Y, He C-J, Ji P-Y, Zhuo Z-Y, Tian X-Z, Wang F, Tan D-X, Liu G-S. Effects of Melatonin on the Proliferation and Apoptosis of Sheep Granulosa Cells under Thermal Stress. International Journal of Molecular Sciences. 2014; 15(11):21090-21104. https://doi.org/10.3390/ijms151121090
Chicago/Turabian StyleFu, Yao, Chang-Jiu He, Peng-Yun Ji, Zhi-Yong Zhuo, Xiu-Zhi Tian, Feng Wang, Dun-Xian Tan, and Guo-Shi Liu. 2014. "Effects of Melatonin on the Proliferation and Apoptosis of Sheep Granulosa Cells under Thermal Stress" International Journal of Molecular Sciences 15, no. 11: 21090-21104. https://doi.org/10.3390/ijms151121090
APA StyleFu, Y., He, C. -J., Ji, P. -Y., Zhuo, Z. -Y., Tian, X. -Z., Wang, F., Tan, D. -X., & Liu, G. -S. (2014). Effects of Melatonin on the Proliferation and Apoptosis of Sheep Granulosa Cells under Thermal Stress. International Journal of Molecular Sciences, 15(11), 21090-21104. https://doi.org/10.3390/ijms151121090