Identification and Characterization of Buffalo 7SK and U6 pol III Promoters and Application for Expression of Short Hairpin RNAs

Abstract
:1. Introduction
2. Results
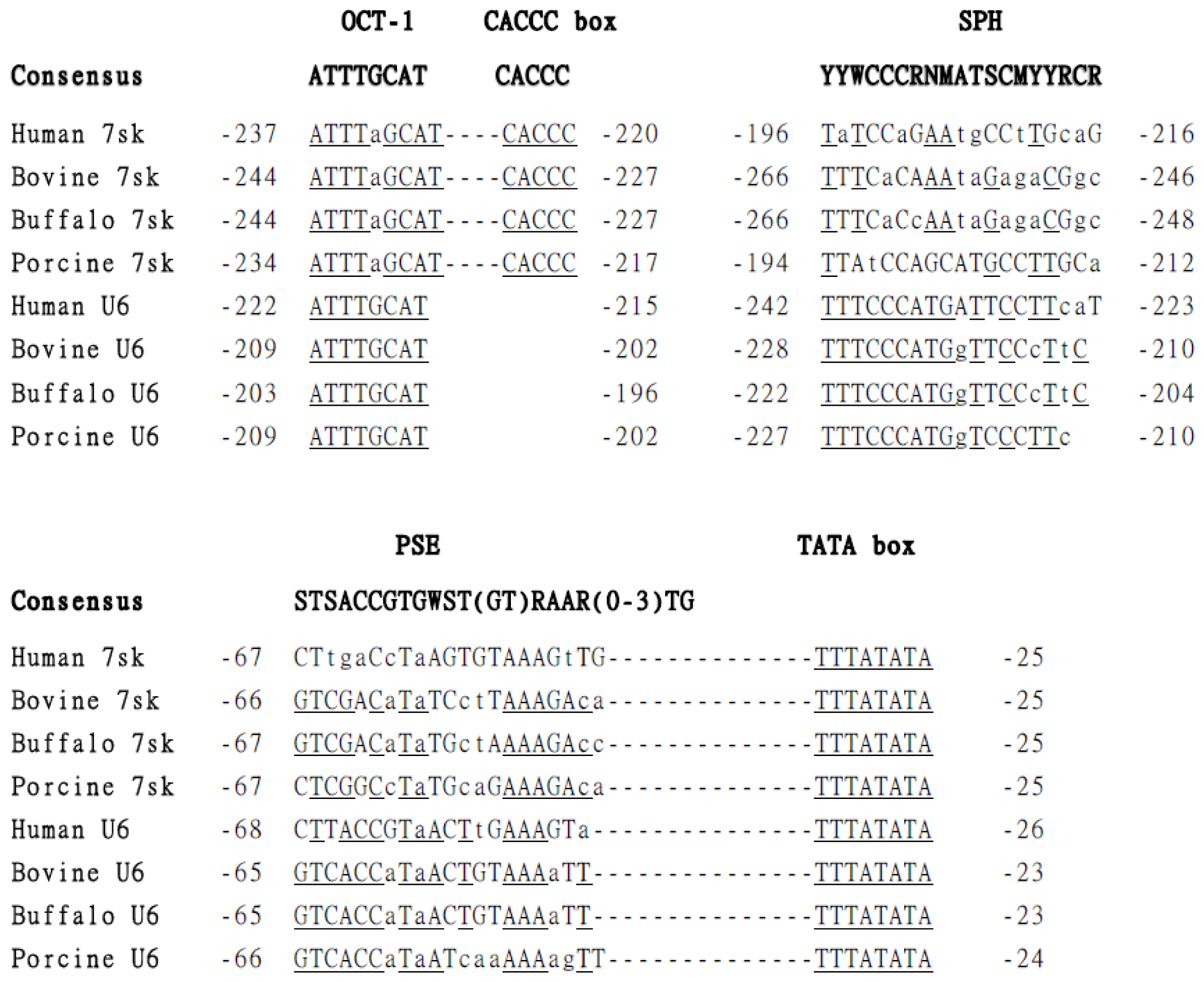
2.1. Identification and Isolation of Buffalo 7SK and U6 Promoters
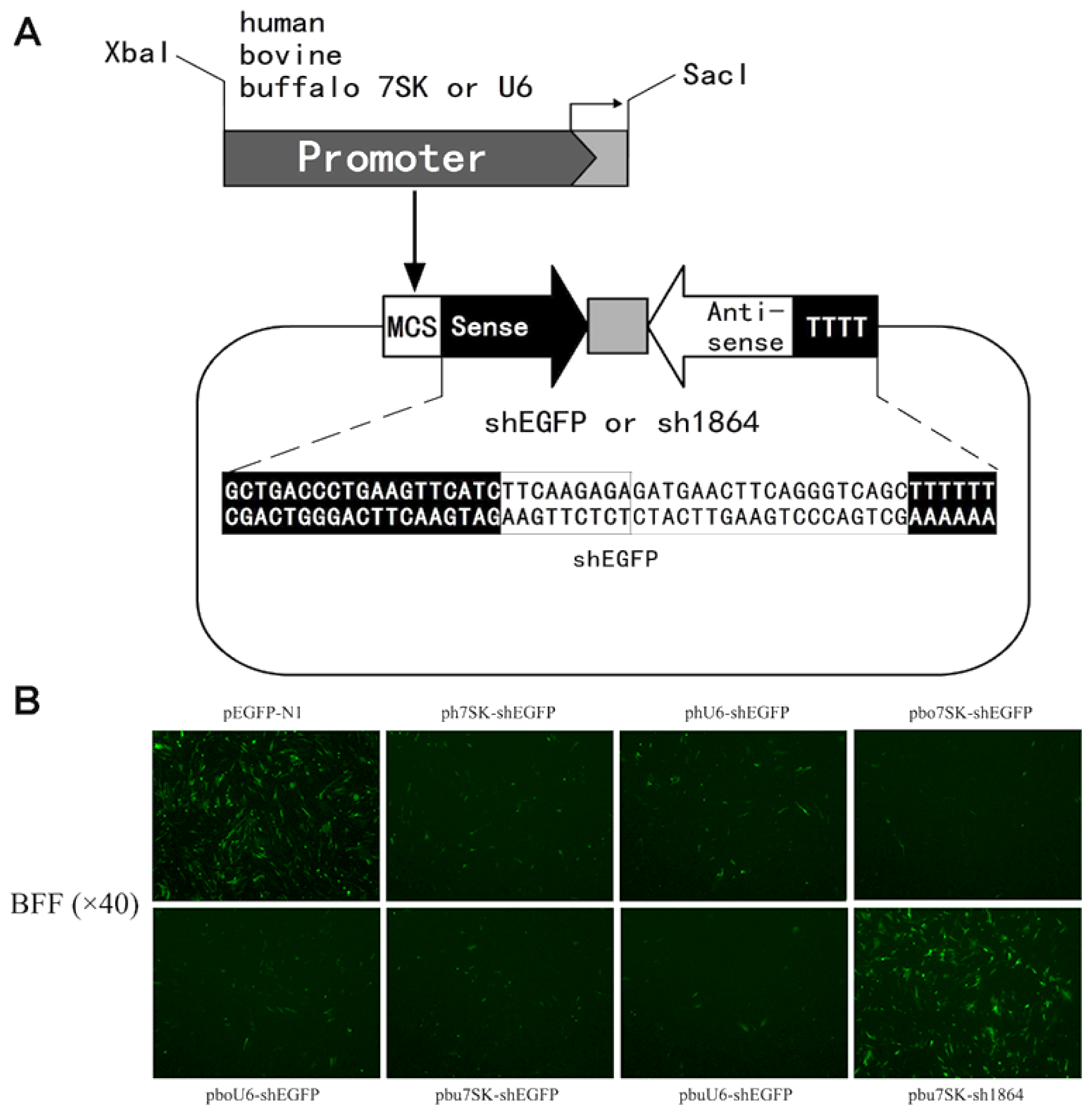
2.2. Construction and Validation of shRNA Expression Vectors
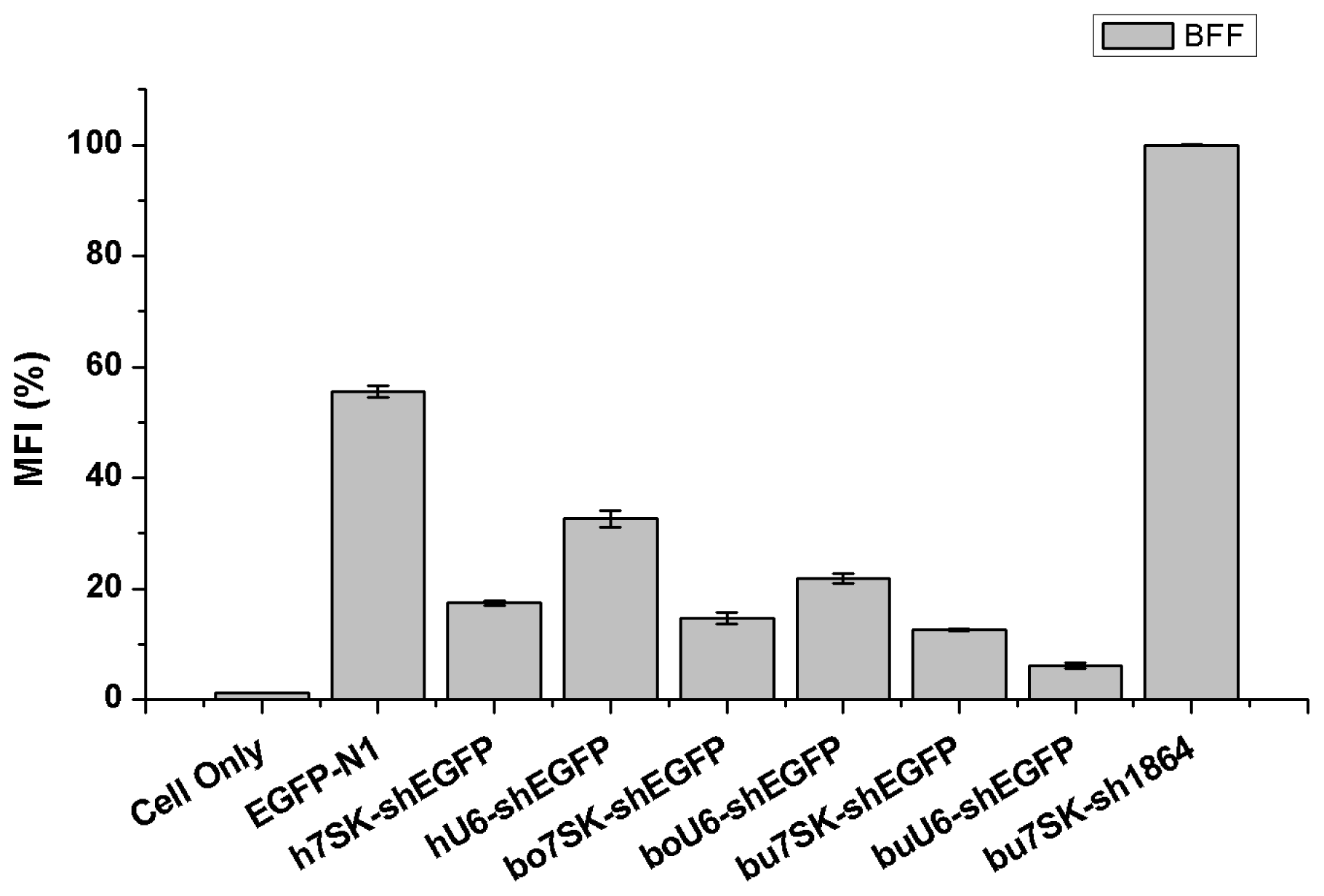
2.3. The bu7SK and buU6 Promoters Direct shRNA-Mediated Knockdown
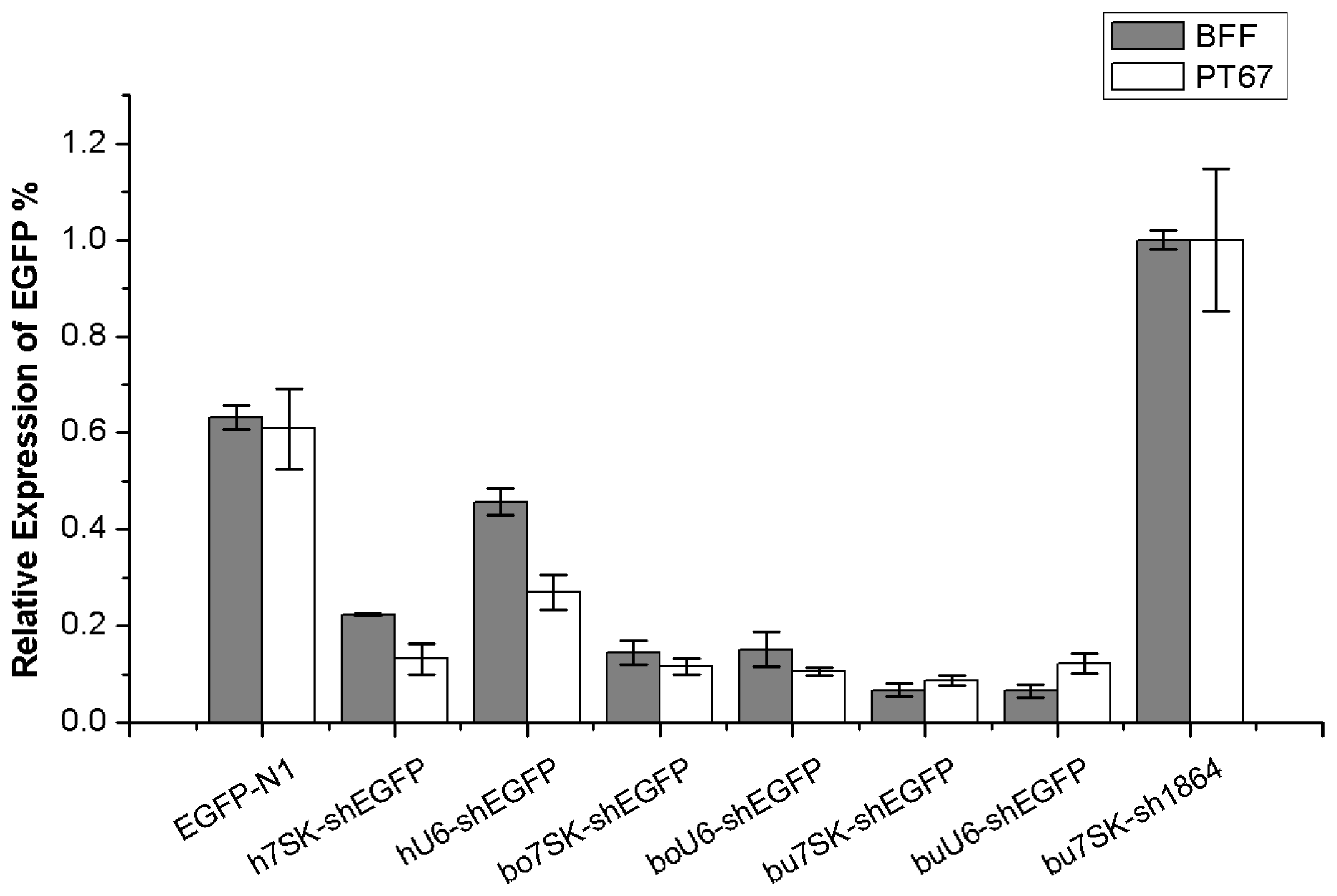
2.4. Comparison of Knockdown Efficiency by qRT-PCR
3. Discussion
4. Experimental Section
4.1. Isolation of the bu7SK and buU6 Promoter from Buffalo Genomic DNA
4.2. Construction of 7SK and U6 shRNA Expression Vectors
4.3. Cell Culture and Transfection
4.4. Flow Cytometry Assay
4.5. Real-Time PCR Assay
5. Conclusions
Acknowledgments
Conflicts of Interest
References
- Fire, A.; Xu, S.; Montgomery, M.K.; Kostas, S.A.; Driver, S.E.; Mello, C.C. Potent and specific genetic interference by double-stranded RNA in caenorhabditis elegans. Nature 1998, 391, 806–811. [Google Scholar]
- Elbashir, S.M.; Harborth, J.; Lendeckel, W.; Yalcin, A.; Weber, K.; Tuschl, T. Duplexes of 21-nucleotide RNAs mediate RNA interference in cultured mammalian cells. Nature 2001, 411, 494–498. [Google Scholar]
- Brummelkamp, T.R.; Bernards, R.; Agami, R. A system for stable expression of short interfering RNAs in mammalian cells. Science 2002, 296, 550–553. [Google Scholar]
- Paul, C.P.; Good, P.D.; Winer, I.; Engelke, D.R. Effective expression of small interfering RNA in human cells. Nat. Biotechnol 2002, 20, 505–508. [Google Scholar]
- Amarzguioui, M.; Rossi, J.J.; Kim, D. Approaches for chemically synthesized siRNA and vector-mediated RNAi. FEBS Lett 2005, 579, 5974–5981. [Google Scholar]
- Schramm, L.; Hernandez, N. Recruitment of RNA polymerase III to its target promoters. Genes Dev 2002, 16, 2593–2620. [Google Scholar]
- Geiduschek, E.P.; Kassavetis, G.A. The RNA polymerase III transcription apparatus. J. Mol. Biol 2001, 310, 1–26. [Google Scholar]
- Murphy, S.; Pierani, A.; Scheidereit, C.; Melli, M.; Roeder, R.G. Purified octamer binding transcription factors stimulate RNA polymerase III-mediated transcription of the 7SK RNA gene. Cell 1989, 59, 1071–1080. [Google Scholar]
- Danzeiser, D.A.; Urso, O.; Kunkel, G.R. Functional characterization of elements in a human U6 small nuclear RNA gene distal control region. Mol. Cell Biol 1993, 13, 4670–4678. [Google Scholar]
- Schaub, M.; Myslinski, E.; Schuster, C.; Krol, A.; Carbon, P. Staf, a promiscuous activator for enhanced transcription by RNA polymerases II and III. EMBO J 1997, 16, 173–181. [Google Scholar]
- Kunkel, G.R.; Cheung, T.C.; Miyake, J.H.; Urso, O.; McNamara-Schroeder, K.J.; Stumph, W.E. Identification of a SPH element in the distal region of a human U6 small nuclear RNA gene promoter and characterization of the sph binding factor in Hela cell extracts. Gene Expr 1996, 6, 59–72. [Google Scholar]
- Rincon, J.C.; Engler, S.K.; Hargrove, B.W.; Kunkel, G.R. Molecular cloning of a cdna encoding human SPH-binding factor, a conserved protein that binds to the enhancer-like region of the U6 small nuclear RNA gene promoter. Nucleic Acids Res 1998, 26, 4846–4852. [Google Scholar]
- Kleinert, H.; Bredow, S.; Benecke, B.J. Expression of a human 7SK RNA gene in vivo requires a novel pol III upstream element. EMBO J 1990, 9, 711–718. [Google Scholar]
- Ma, B.; Hernandez, N. Redundant cooperative interactions for assembly of a human U6 transcription initiation complex. Mol. Cell Biol 2002, 22, 8067–8078. [Google Scholar]
- Kunkel, G.R.; Pederson, T. Upstream elements required for efficient transcription of a human U6 RNA gene resemble those of U1 and U2 genes even though a different polymerase is used. Genes Dev 1988, 2, 196–204. [Google Scholar]
- James Faresse, N.; Canella, D.; Praz, V.; Michaud, J.; Romascano, D.; Hernandez, N. Genomic study of RNA polymerase II and III snapc-bound promoters reveals a gene transcribed by both enzymes and a broad use of common activators. PLoS Genet 2012, 8, e1003028. [Google Scholar]
- Domitrovich, A.M.; Kunkel, G.R. Multiple, dispersed human U6 small nuclear RNA genes with varied transcriptional efficiencies. Nucleic Acids Res 2003, 31, 2344–2352. [Google Scholar]
- Heden, C.G. On the estimation of fifty percent end-points in serological titrimetry. J. Pathol. Bacteriol 1946, 58, 477–481. [Google Scholar]
- Huang, B.; Li, T.; Wang, X.L.; Xie, T.S.; Lu, Y.Q.; da Silva, F.M.; Shi, D.S. Generation and characterization of embryonic stem-like cell lines derived from in vitro fertilization buffalo (Bubalus bubalis) embryos. Reprod. Domest. Anim 2010, 45, 122–128. [Google Scholar]
- Shi, D.; Lu, F.; Wei, Y.; Cui, K.; Yang, S.; Wei, J.; Liu, Q. Buffalos (Bubalus bubalis) cloned by nuclear transfer of somatic cells. Biol. Reprod 2007, 77, 285–291. [Google Scholar]
- Sturm, R.A.; Das, G.; Herr, W. The ubiquitous octamer-binding protein OCT-1 contains a pou domain with a homeo box subdomain. Genes Dev 1988, 2, 1582–1599. [Google Scholar]
- Dahlberg, J.E.; Lund, E. The Genes and Transcription of the Major Small Nuclear RNAs. In Structure and Function of Major and Minor Small Nuclear Ribonucleoprotein Particles; Birnstiel, M.L., Ed.; Springer-Verlag: Berlin, Germany, 1988; Volume 88, pp. 38–70. [Google Scholar]
- Cullen, B.R. Induction of stable RNA interference in mammalian cells. Gene Ther 2006, 13, 503–508. [Google Scholar]
- Cockrill, W.R. The draught buffalo (Bubalus bubalis). Veterinarian 1968, 5, 265–272. [Google Scholar]
- Cockrill, W.R. The water buffalo: A review. Br. Vet. J 1981, 137, 8–16. [Google Scholar]
- Servan de Almeida, R.; Keita, D.; Libeau, G.; Albina, E. Control of ruminant morbillivirus replication by small interfering RNA. J. Gen. Virol 2007, 88, 2307–2311. [Google Scholar]
- Liu, M.; Chen, W.; Ni, Z.; Yan, W.; Fei, L.; Jiao, Y.; Zhang, J.; Du, Q.; Wei, X.; Chen, J.; et al. Cross-inhibition to heterologous foot-and-mouth disease virus infection induced by RNA interference targeting the conserved regions of viral genome. Virology 2005, 336, 51–59. [Google Scholar]
- Kahana, R.; Kuznetzova, L.; Rogel, A.; Shemesh, M.; Hai, D.; Yadin, H.; Stram, Y. Inhibition of foot-and-mouth disease virus replication by small interfering RNA. J. Gen. Virol 2004, 85, 3213–3217. [Google Scholar]
- Xie, Q.; Jiang, J.; Wang, D.; Shi, D.; Xie, T.; Chen, S.; Liu, Q. Construction and analysis of buffalo inhibin α-subunit gene RNAi vectors. Chin. J. Vet. Sci 2011, 31, 1070–1075. [Google Scholar]
- Murphy, S.; di Liegro, C.; Melli, M. The in vitro transcription of the 7SK RNA gene by RNA polymerase III is dependent only on the presence of an upstream promoter. Cell 1987, 51, 81–87. [Google Scholar]
- Lambeth, L.S.; Wise, T.G.; Moore, R.J.; Muralitharan, M.S.; Doran, T.J. Comparison of bovine RNA polymerase III promoters for short hairpin RNA expression. Anim. Genet 2006, 37, 369–372. [Google Scholar]
- Bannister, S.C.; Wise, T.G.; Cahill, D.M.; Doran, T.J. Comparison of chicken 7SK and U6 RNA polymerase III promoters for short hairpin RNA expression. BMC Biotechnol 2007, 7, 79. [Google Scholar]
- Moon, I.S.; Krause, M.O. Common RNA polymerase I, II, and III upstream elements in mouse 7SK gene locus revealed by the inverse polymerase chain reaction. DNA Cell Biol 1991, 10, 23–32. [Google Scholar]
- Chen, X.; Liu, X.; Wang, W.; Wang, J.; Meng, Q.; Zhang, D. Chicken 7SK promoter drives efficient shRNA transcription with species specificity. Res. Vet. Sci 2013, 95, 1006–1011. [Google Scholar]
- Cummins, D.; Doran, T.J.; Tyack, S.; Purcell, D.; Hammond, J. Identification and characterisation of the porcine 7SK RNA polymerase III promoter for short hairpin RNA expression. J. RNAi Gene Silenc 2008, 4, 289–294. [Google Scholar]
- Boyd, D.C.; Greger, I.H.; Murphy, S. In vivo footprinting studies suggest a role for chromatin in transcription of the human 7SK gene. Gene 2000, 247, 33–44. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer name | Sequence |
|---|---|
| bu7SK-F | 5′-TCTAGAGAGACAGACCTGGCTCCAC-3′ a |
| bu7SK-R | 5′-GAGCTCCACATCCGAGACACTCTGC-3′ b |
| buU6-F | 5′-ATCGATGAGCATTCAGTCCGGCAG-3′ c |
| buU6-R | 5′-CTCGAGGCACGGTAAACATGGCTTC-3′ d |
| h7SK-F | 5′-TCTAGACTGCAGTATTTAGCATGCCCC-3′ a |
| h7SK-R | 5′-GAGCTCGAGGTACCCAGGCGGC-3′ b |
| hU6-F | 5′-ATCGATCCCCCGAGTCCAACAC-3′ c |
| hU6-R | 5′-CTCGAGGTGTTTCGTTCTTTCCACAAG-3′ d |
| bo7SK-F | 5′-TCTAGAGAGCGTGAGAGACTCGGAGC-3′ a |
| bo7SK-R | 5′-GAGCTCCACCATGGTAGTTCGCGCAG-3′ b |
| boU6-F | 5′-ATCGATAAGAGGCTCCTGAGCAACG-3′ c |
| boU6-R | 5′-CTCGAGCATTTACCCCCCACAGAATATATAAG-3′ d |
© 2014 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Zhang, X.; Liu, Q.; Luo, C.; Deng, Y.; Cui, K.; Shi, D. Identification and Characterization of Buffalo 7SK and U6 pol III Promoters and Application for Expression of Short Hairpin RNAs. Int. J. Mol. Sci. 2014, 15, 2596-2607. https://doi.org/10.3390/ijms15022596
Zhang X, Liu Q, Luo C, Deng Y, Cui K, Shi D. Identification and Characterization of Buffalo 7SK and U6 pol III Promoters and Application for Expression of Short Hairpin RNAs. International Journal of Molecular Sciences. 2014; 15(2):2596-2607. https://doi.org/10.3390/ijms15022596
Chicago/Turabian StyleZhang, Xiaoxi, Qingyou Liu, Chan Luo, Yanfei Deng, Kuiqing Cui, and Deshun Shi. 2014. "Identification and Characterization of Buffalo 7SK and U6 pol III Promoters and Application for Expression of Short Hairpin RNAs" International Journal of Molecular Sciences 15, no. 2: 2596-2607. https://doi.org/10.3390/ijms15022596
APA StyleZhang, X., Liu, Q., Luo, C., Deng, Y., Cui, K., & Shi, D. (2014). Identification and Characterization of Buffalo 7SK and U6 pol III Promoters and Application for Expression of Short Hairpin RNAs. International Journal of Molecular Sciences, 15(2), 2596-2607. https://doi.org/10.3390/ijms15022596