The Effect of Physical and Chemical Cues on Hepatocellular Function and Morphology


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
2.1. Influence on Functional Protein Expression
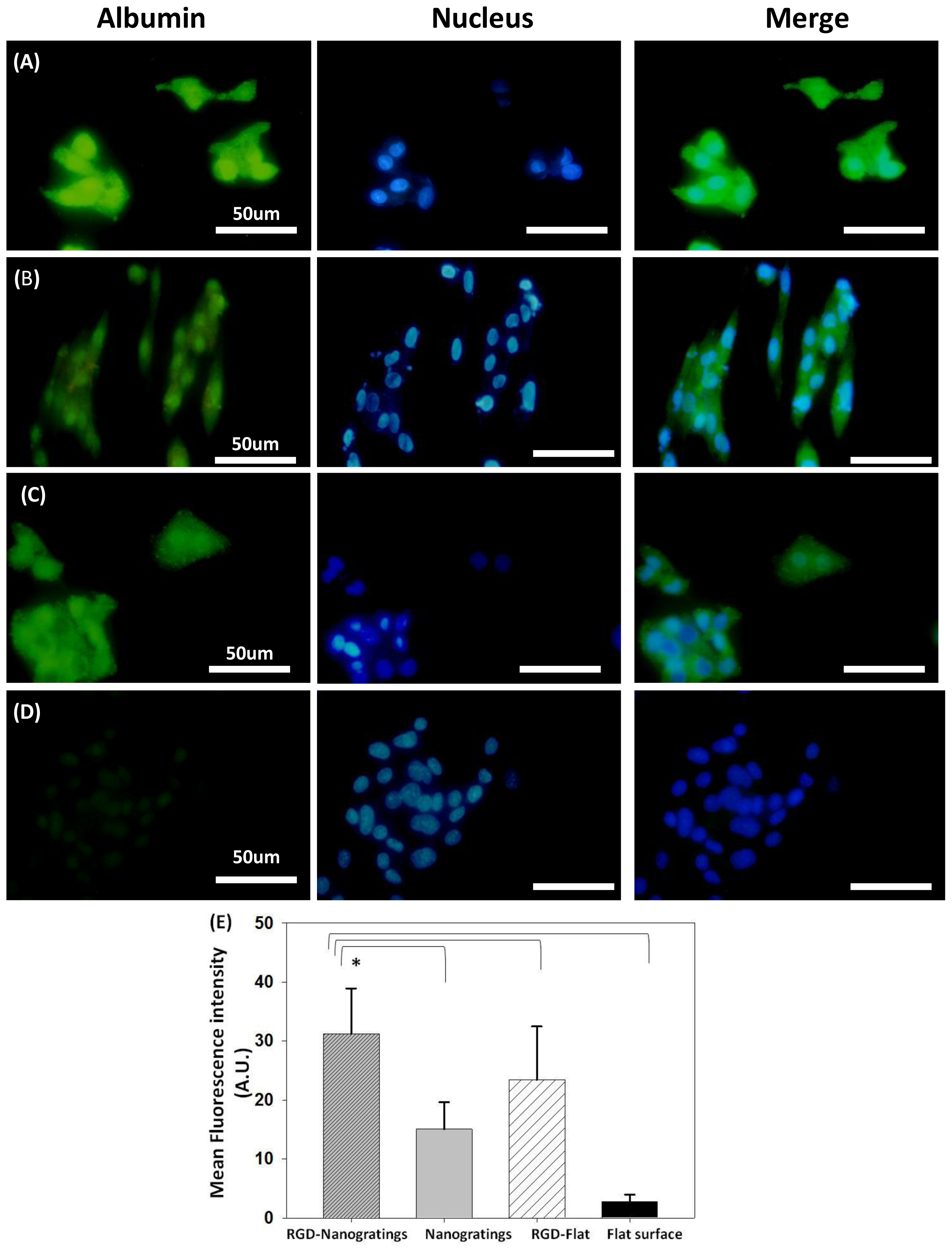
2.1.1. Albumin
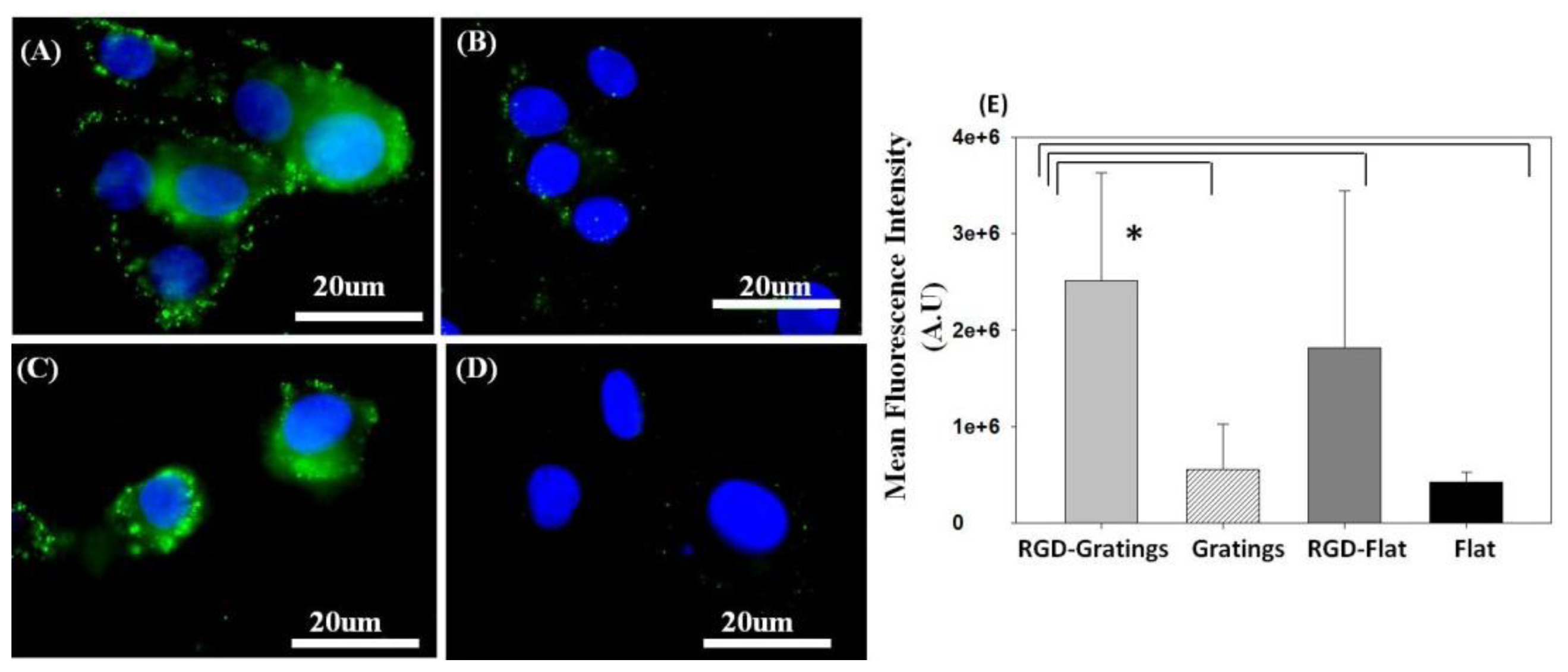
2.1.2. Transferrin
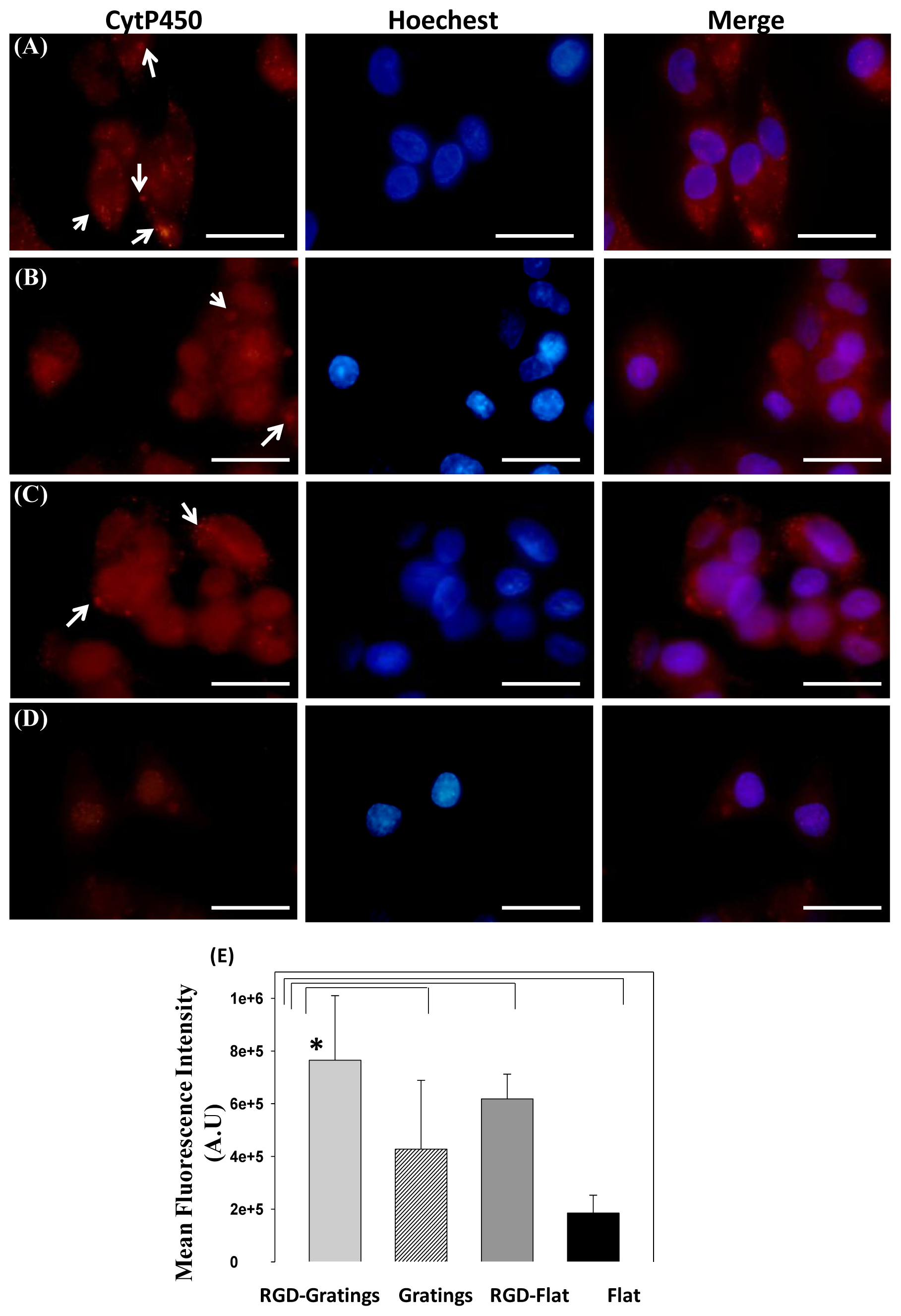
2.1.3. Cytochrome P-450
2.2. Influence on the Structure of Hepatic Cell Line (HepG2)
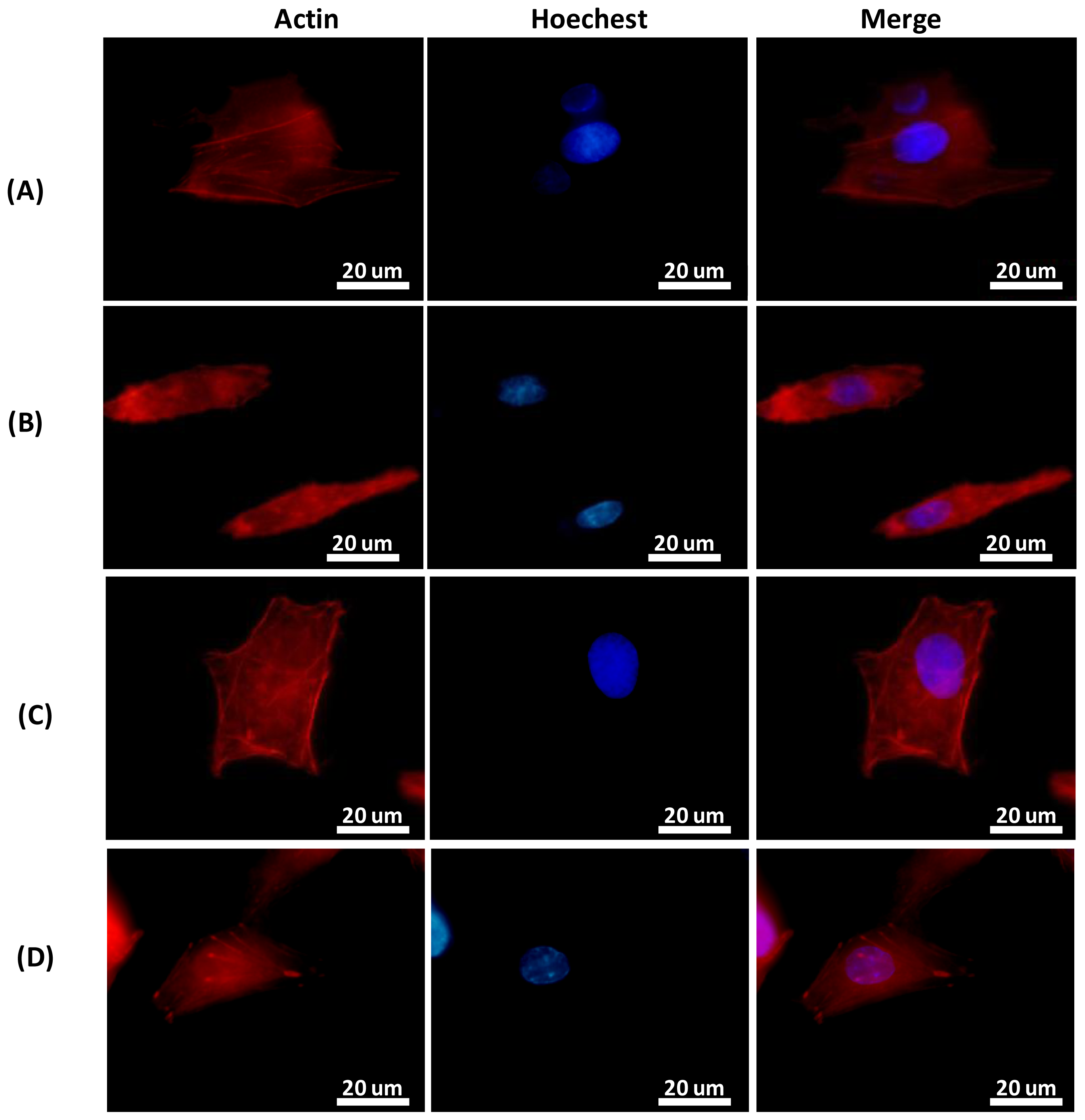
2.2.1. Cell Alignment and Actin Filament Rearrangement
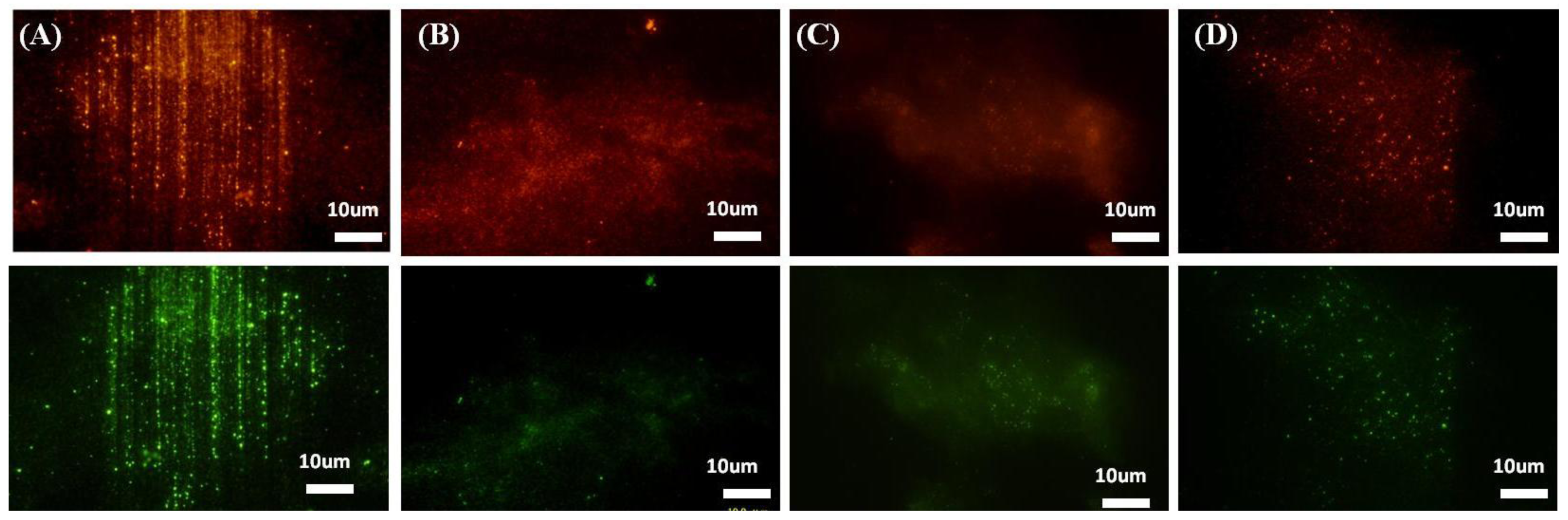
2.2.2. Integrin Mediated Focal Adhesion
2.2.3. Alteration in Natural Extracellular Matrix Assembly
3. Experimental Section
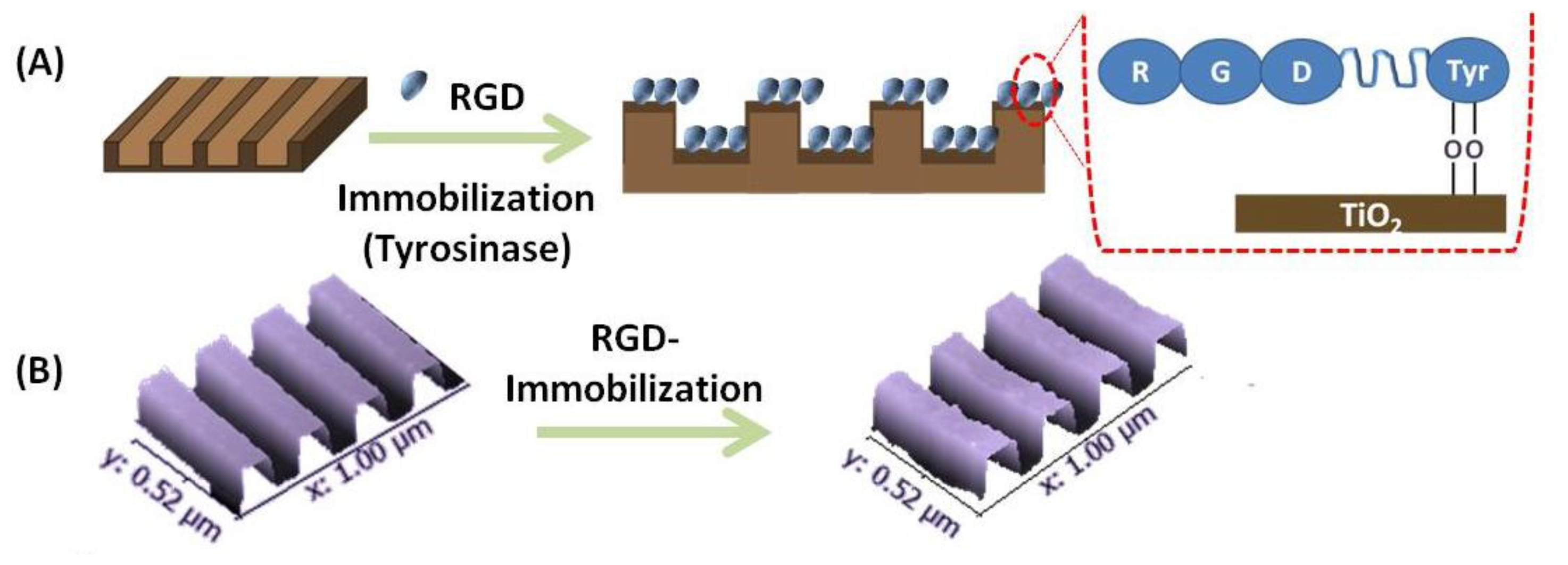
3.1. Immobilization of RGD on TiO2 Nanopatterned Substrates
3.2. Hepatic Cell Line Culture
3.3. Immunostaining and Fluorescence Detection
3.4. Scanning Electron Microscope Investigation
4. Conclusions
Supplementary Information
ijms-15-04299-s001.pdfAcknowledgments
Conflicts of Interest
References
- Engel, J.; Chiquet, M. An over view of extracellular matrix and function. In The Extracellular Matrix: An Over View; Mecham, R.P., Ed.; Springer: Berlin/Heidelberg, Germany, 2011; pp. 1–41. [Google Scholar]
- Lukashev, M.E.; Werb, Z. ECM signalling: Orchestrating cell behaviour and misbehaviour. Trends Cell Biol. 1998, 8, 437–441. [Google Scholar]
- Pamula, E.; de Cupere, V.; Dufrene, Y.F.; Rouxhet, P.G. Nanoscale organization of adsorbed collagen: Influence of substrate hydrophobicity and adsorption time. J. Colloid Interface Sci. 2004, 271, 80–91. [Google Scholar]
- Bozec, L.; van der Heijden, G.; Horton, M. Collagen fibrils: Nanoscale ropes. Biophys. J 2007, 92, 70–75. [Google Scholar]
- Vaday, G.G.; Lider, O. Extracellular matrix moieties cytokines and enzymes: Dynamic effects on immune cell behavior and inflammation. J. Leukoc. Biol. 2000, 67, 149–159. [Google Scholar]
- Bosman, F.T.; Stamenkovic, I. Functional structure and composition of the extracellular matrix. J. Pathol. 2003, 200, 423–428. [Google Scholar]
- Zeisberg, M.; Maeshima, Y.; Mosterman, B.; Kalluri, R. Renal fibrosis Extracellular matrix microenvironment regulates migratory behavior of activated tubular epithelial cells. Am. J. Pathol. 2002, 160, 2001–2008. [Google Scholar]
- Heino, J. The collagen receptor integrins have distinct ligand recognition and signaling functions. Matrix Biol. 2000, 19, 319–323. [Google Scholar]
- Ahmed, N.; Riley, C.; Rice, G.; Quinn, M. Role of integrin receptors for fibronectin collagen and laminin in the regulation of ovarian carcinoma functions in response to a matrix microenvironment. Clin. Exp. Metastasis 2005, 22, 391–402. [Google Scholar]
- Biggs, M.J.P.; Ritchard, R.G.; Dalby, M.J. Nanotopographical modification: A regulator of cellular function through focal adhesions. Nanomed.: Nanotechnol. Biol. Med. 2010, 6, 619–633. [Google Scholar]
- Wozniak, M.A.; Modzelewska, K.; Kwong, L.; Keely, P.J. Focal adhesion regulation of cell behavior. Biochim. Biophys. Acta 2004, 1692, 103–119. [Google Scholar]
- Maheshwari, G.; Brown, G.; Lauffenburger, D.A.; Wells, A.; Griffith, L.G. Cell adhesion and motility depend on nanoscale RGD clustering. J. Cell Sci. 2000, 113, 1677–1686. [Google Scholar]
- Rowley, J.A.; Mooney, D.J. Alginate type and RGD density control myoblast phenotype. J. Biomed. Mater. Res. 2002, 60, 217–223. [Google Scholar]
- Wang, N.; Ingber, D.E. Control of cytoskeletal mechanics by extracellular cell shape and mechanical tension. Biophys. J. 1994, 66, 2181–2189. [Google Scholar]
- Abrams, G.A.; Goodman, S.L.; Nealy, P.F.; Franco, M.; Murphy, C.J. Nanoscale topography of the basement membrane underlying the corneal epithelium of the rhesus macaque. Cell Tissue Res. 2000, 299, 39–46. [Google Scholar]
- Hornberger, L.K.; Singhroy, S.; Cavalle-Garrido, T.; Tsang, W.; Keeley, F.; Rabinovitch, M. Synthesis of extracellular matrix and adhesion through β1 integrins are critical for fetal ventricular myocyte proliferation. Circ. Res. 2000, 87, 508–515. [Google Scholar]
- Wu, C.; Dedhar, S. Integrin-linked kinase (ILK) and its interactors: A new paradigm for the coupling of extracellular matrix to actin cytoskeleton and signaling complexes. J. Cell Biol. 2001, 155, 505–510. [Google Scholar]
- Zhu, B.; Lu, Q.; Yin, J.; Hu, J.; Wang, Z. Alignment of osteoblast-like cells and cell-produced collagen matrix induced by nanogrooves. Tissue Eng. 2005, 11, 825–834. [Google Scholar]
- Watari, S.; Hayashi, K.; Wood, J.A.; Russell, P.; Nealey, P.F.; Murphy, C.J.; Genetos, D.C. Modulation of osteogenic differentiation in hMSCs cells by submicron topographically-patterned ridges and grooves. Biomaterials 2012, 33, 128–136. [Google Scholar]
- Kulangara, K.; Yang, Y.; Yang, J.; Leong, K.W. Nanotopography as modulator of human mesenchymal stem cells function. Biomaterials 2012, 33, 4998–5003. [Google Scholar]
- Biela, S.A.; Su, Y.; Spatz, J.P.; Kemkemer, R. Different sensitivity of human endothelial cells smooth muscle cells and fibroblasts to topography in the nano-micro range. Acta Biomater. 2009, 5, 2460–2466. [Google Scholar]
- Choi, C.-H.; Hagvallb, S.H.; Wu, B.M.; Dunn, J.C.Y.; Beygui, R.E.; Kim, C.-J. Cell interaction with three-dimensional sharp-tip nanotopography. Biomaterials 2007, 28, 1672–1679. [Google Scholar]
- Wang, P.-Y.; Wu, T.-H.; Chao, P.-H.G.; Kuo, W.-H.; Wang, M.-J.; Hsu, C.-C.; Tsai, W.-B. Modulation of cell attachment and collagen production of anterior cruciate ligament cells via submicron grooves/ridges structures with different cell affinity. Biotechnol. Bioeng. 2013, 110, 327–337. [Google Scholar]
- Kong, Y.P.; Tu, C.; Donovan, P.J.; Yee, A.F. Expression of Oct4 in human embryonic stem cells is dependant on nanotopographicalconfiguration. Acta Biomater. 2013, 9, 6369–6380. [Google Scholar]
- Takayama, G.; Taniguchi, A.; Okano, T. Identification of Differentiall expressed genes in hepatocyte/endothelial cell co-culture system. Tissue Eng. 2007, 13, 159–166. [Google Scholar]
- Bierwolf, J.; Lutgehetmann, M.; Feng, K.; Erbes, J.; Deichmann, S.; Toronyi, E.; Stieglitz, C.; Nashan, B.; Ma, P.X.; Pollok, J.M. Primary rat hepatocyte culture on 3D nanofibrous polymer scaffolds for toxicology and pharmaceutical research. Biotech. Bioeng. 2011, 108, 141–150. [Google Scholar]
- Nishida, Y.; Taniguchi, A. Induction of albumin expression in HepG2 cells using immobilized simplified recombinant fibronectin protein. In Vitro Cell. Dev. Biol.-Anim. 2013, 49, 400–407. [Google Scholar]
- Hoessa, A.; Teuschera, N.; Thormanna, A.; Aurichb, H.; Heilmanna, A. Cultivation Of Hepatoma Cell Line HepG2 On Nanoporous Aluminum Oxide Membranes. Acta Biomater. 2007, 3, 43–50. [Google Scholar]
- Nakazawa, K.; Lee, S.-W.; Fukuda, J.; Yang, D.-H.; Kunitake, T. Hepatocyte spheroid formation on a titanium dioxide gel surface and hepatocyte long-term culture. J. Mater. Sci. 2006, 17, 359–364. [Google Scholar]
- Abdellatef, S.A.; Ohi, A.; Nabatame, T.; Taniguchi, A. Induction of hepatocyte functional protein expression by submicron/nano-patterning substrates to mimic in vivo structures. Biomater. Sci. 2014, 2, 330–338. [Google Scholar]
- Prockop, D.J.; Fertala, A. The collagen fibril: The almost crystalline structure. J. Struct. Biol. 1998, 122, 111–118. [Google Scholar]
- Bokhari, M.; Carnachan, R.J.; Cameron, N.R.; Przyborski, S.A. during toxicological challenge. J. Anat. 2007, 211, 567–576. [Google Scholar]
- Soldatow, V.Y.; LeCluyse, E.L.; Griffth, L.G.; Rusyn, I. In vitro models for liver toxicity testing. Toxicol. Res. 2013, 2, 23–39. [Google Scholar]
- Dalby, M.J.; Gadegaard, N.; Tare, R.; Andar, A.; Riehle, M.O.; Herzyk, P.; Wilkinson, C.D.W.; Oreffo, R.O.C. The control of human mesenchymal cell differentiation using nanoscale symmetry and disorder. Nat. Mater. 2007, 6, 997–1003. [Google Scholar]
- You, M.-H.; Kwak, M.K.; Kim, D.-H.; Kim, K.; Levchenko, A.; Kim, D.-Y.; Suh, K.-Y. Synergistically Enhanced osteogenic differentiation of human mesenchymal stem cells by culture on nanostructured surfaces with induction media. Biomacromolecules 2010, 11, 1856–1862. [Google Scholar]
- Hamashima, Y.; Harter, J.G.; Coons, A.H. The Localization of Albumin and fibrinogen in human liver cells. J. Cell Biol. 1964, 20, 271–279. [Google Scholar]
- Kocarek, T.A.; Schuetz, E.G.; Guzelian, P.S. Expression of multiple forms of cytochrome p450 mrnas in primary cultures of rat hepatocytes maintained on matrigel. Mol. Pharmacol. 1993, 43, 328–334. [Google Scholar]
- Shaw, P.M.; Weiss, M.C.; Adesnik, M. hepatocyte nuclear factor 3 is a major determinant of cyp2c6 promoter activity in hepatoma cells. Mol. Pharmacol. 1994, 46, 79–87. [Google Scholar]
- Cai, K.; Frant, M.; Bossert, J.; Hildebrand, G.; Liefeith, K.; Jandt, K.D. Surface functionalized titanium thin films: Zeta-potential protein adsorption and cell proliferation. Colloids Surf. B 2006, 50, 1–8. [Google Scholar]
- Porté-Durrieu, M.C.; Guillemot, F.; Pallu, S.; Labrugère, C.; Brouillaudc, B.; Bareille, R.; Amédée, J.; Barthe, N.; Dard, M.; Baquey, C. Cyclo-(DfKRG) peptide grafting onto ti-6al-4v: physical characterization and interest towards human osteoprogenitor cells adhesion. Biomaterials 2004, 25, 4837–4846. [Google Scholar]
- Lange, R.; Lüthen, F.; Beck, U.; Rychly, J.; Baumann, A.; Nebe, B. Cell-extracellular matrix interaction and physico-chemical characteristics of titanium surfaces depend on the roughness of the material. Biomol. Eng. 2002, 19, 255–261. [Google Scholar]
- Curtis, A.S.; Casey, B.; Gallagher, J.O.; Pasqui, D.; Wood, M.A.; Wilkinson, C.D. Substratum nanotopography and the adhesion of biological cells Are symmetry or regularity of nanotopography important? Biophys. Chem. 2001, 94, 275–283. [Google Scholar]
- Sato, M.; Aslani, A.; Sambito, M.A.; Kalkhoran, N.M.; Slamovich, E.B.; Webster, T.J. Nanocrystalline hydroxyapatite/titania coatings on titanium improves osteoblast adhesion. J. Biomed. Mater. Res. A 2008, 84, 265–272. [Google Scholar]
- Park, K.M.; Park, K.D. Facile surface Immobilization of cell adhesive peptide onto TiO2 substrate via tyrosinase-catalyzed oxidative reaction. J. Mater. Chem. 2011, 21, 15906–15908. [Google Scholar]
- Burgess, A.; Vigneron, S.; Brioudes, E.; Labbé, J.-C.; Lorca, T.; Castro, A. Loss of human Greatwall results in G2 arrest and multiple mitotic defects due to deregulation of the cyclin B-Cdc2/PP2A balance. Proc. Natl. Acad. Sci. USA 2010, 107, 12564–12569. [Google Scholar]
- Herrema, H.; Czajkowska, D.; Théard, D.; van der Wouden, J.M.; Kalicharan, D.; Zolghadr, B.; Hoekstra, D.; van Ijzendoorn, S.C.D. Rho Kinase myosin-ii and p42/44 mapk control extracellular matrix-mediated apical bile canalicular lumen morphogenesis in HepG2 cells. Mol. Biol. Cell 2006, 17, 3291–3303. [Google Scholar]









© 2014 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Abdellatef, S.A.; Ohi, A.; Nabatame, T.; Taniguchi, A. The Effect of Physical and Chemical Cues on Hepatocellular Function and Morphology. Int. J. Mol. Sci. 2014, 15, 4299-4317. https://doi.org/10.3390/ijms15034299
Abdellatef SA, Ohi A, Nabatame T, Taniguchi A. The Effect of Physical and Chemical Cues on Hepatocellular Function and Morphology. International Journal of Molecular Sciences. 2014; 15(3):4299-4317. https://doi.org/10.3390/ijms15034299
Chicago/Turabian StyleAbdellatef, Shimaa A., Akihiko Ohi, Toshihide Nabatame, and Akiyoshi Taniguchi. 2014. "The Effect of Physical and Chemical Cues on Hepatocellular Function and Morphology" International Journal of Molecular Sciences 15, no. 3: 4299-4317. https://doi.org/10.3390/ijms15034299
APA StyleAbdellatef, S. A., Ohi, A., Nabatame, T., & Taniguchi, A. (2014). The Effect of Physical and Chemical Cues on Hepatocellular Function and Morphology. International Journal of Molecular Sciences, 15(3), 4299-4317. https://doi.org/10.3390/ijms15034299