Influence of Green, Red and Blue Light Emitting Diodes on Multiprotein Complex Proteins and Photosynthetic Activity under Different Light Intensities in Lettuce Leaves (Lactuca sativa L.)


Abstract
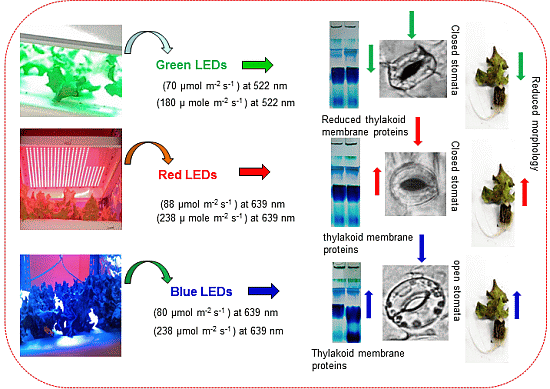
:
1. Introduction
2. Results
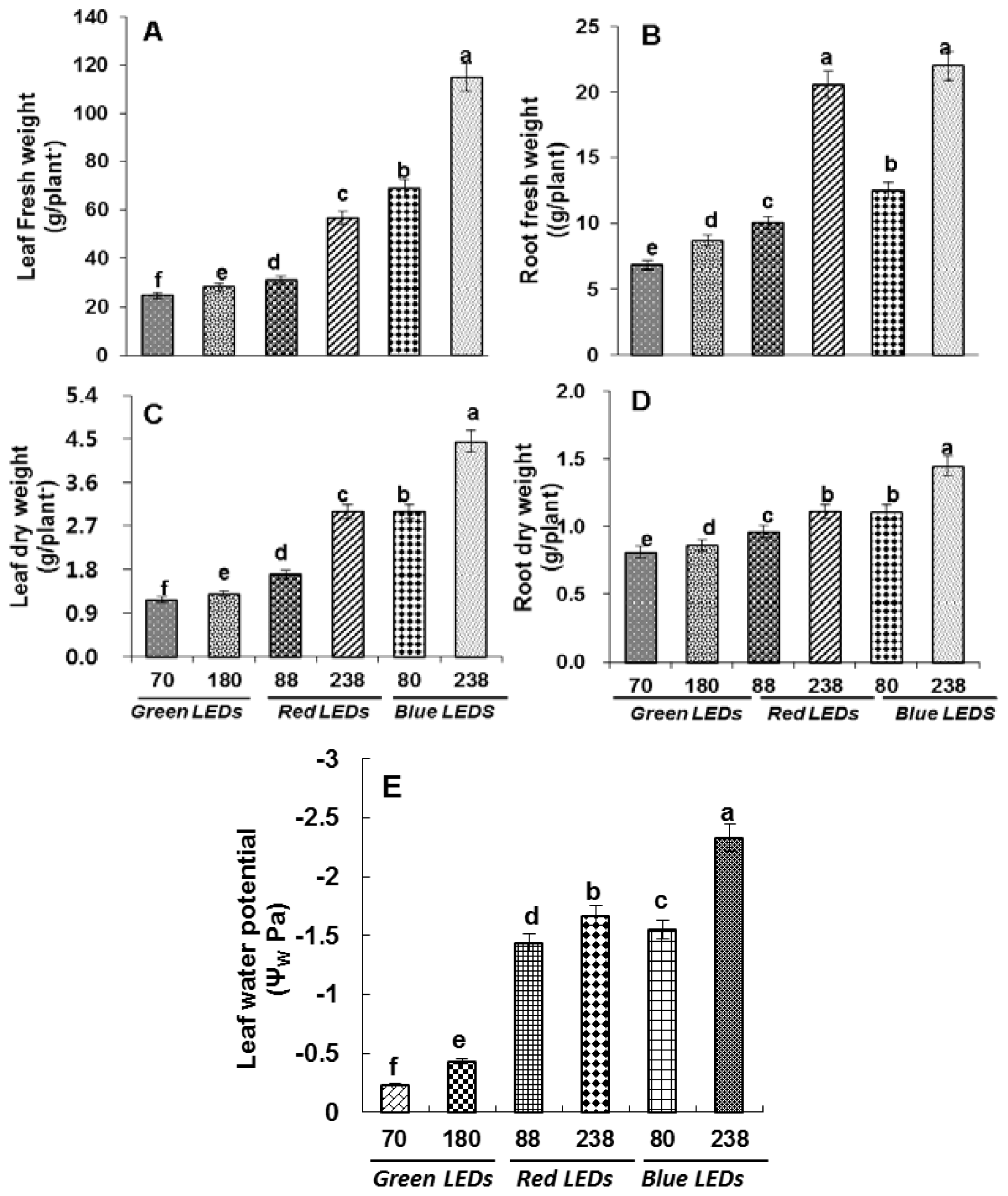
2.1. Growth Analysis and Leaf Water Potential
2.2. Photosynthetic Activity, Stomatal Conductance, Fv/Fm Ratio, and Transpiration
2.3. Stomatal Observations
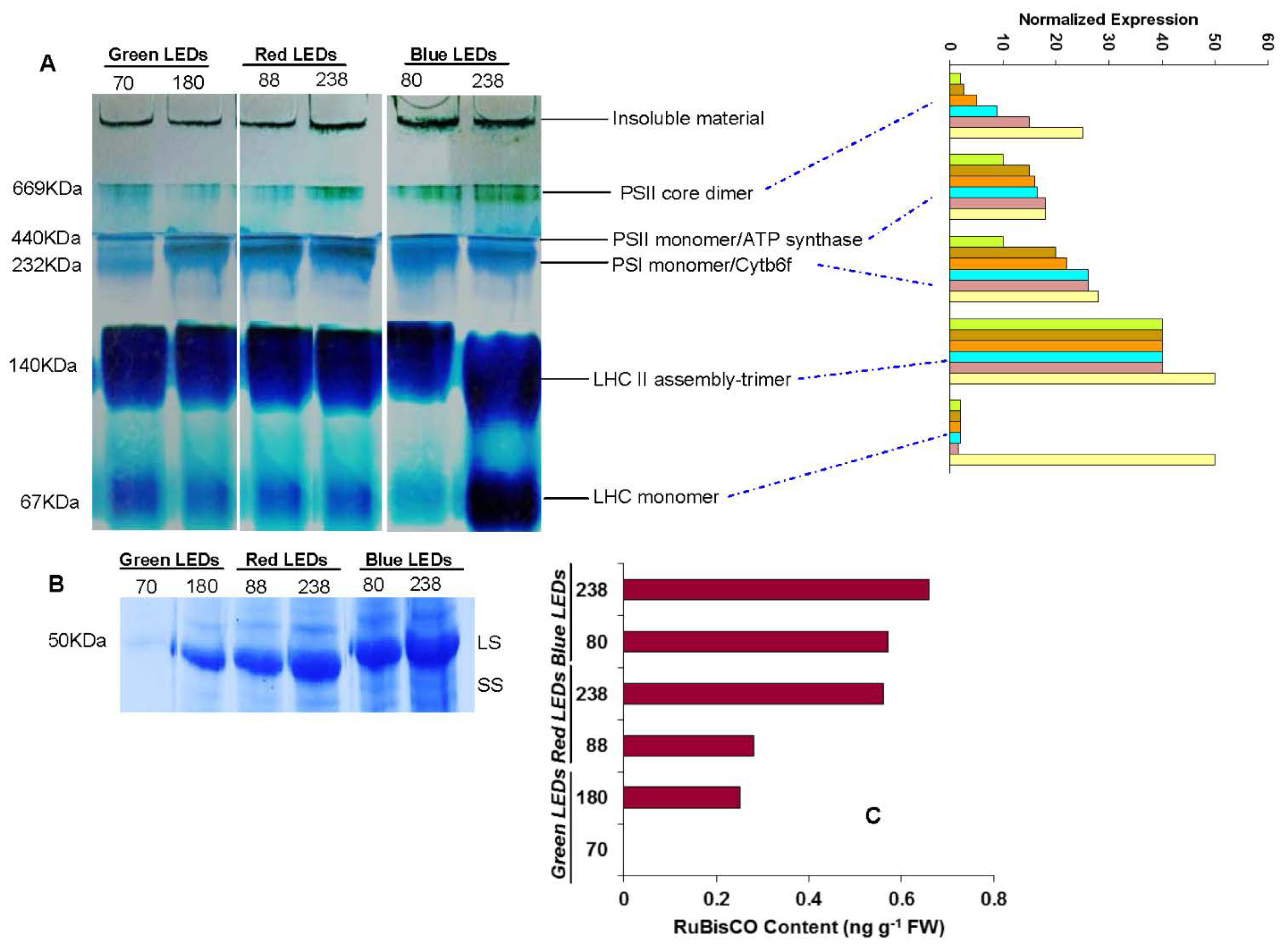
2.4. Thylakoid Membrane Proteins
3. Discussion
4. Material and Methods
4.1. LEDs of Different Light Intensities
4.2. Plant Material and Growth Conditions
4.3. Growth Measurements and ΨW Potential
4.4. Measurement of Photosynthetic Activity
4.5. Observation of Stomata
4.6. Multiprotein Complex Proteins
4.7. RuBisCO Determination by SDS-PAGE
4.8. Statistical Analysis
5. Conclusions
Acknowledgments
Conflicts of Interest
References
- Dong, C.; Fu, Y.; Liu, G.; Liu, H. Low light intensity effects on the growth photosynthetic characteristics 2 antioxidant capacity yield and quality of wheat (Triticum aestivum L) 3 at different growth stages in BLSS. Adv. Space Res. 2014. [Google Scholar] [CrossRef]
- Samuoliene, G.; Brazaityte, A.; Jankauskiene, J.; Virsile, A.; Sirtautas, R.; Novičkovas, A.; Sakalauskienė, S.; Sakalauskaitė, J.; Duchovskis, P. LED irradiance level affects growth and nutritional quality of Brassica microgreens. Cent. Eur. J. Biol. 2013, 8, 1241–1249. [Google Scholar]
- Yano, A.; Fujiwara, K. Plant lighting system with five wavelength-band light-emitting diodes providing photon flux density and mixing ratio control. Plant Methods 2012, 8, 46. [Google Scholar]
- Kim, K.; Kook, H.S.; Jang, Y.J.; Lee, W.H.; Kamala Kannan, S.; Chae, J.C.; Lee, K.J. The effect of blue-light-emitting diodes on antioxidant properties and resistance to Botrytis cinerea in tomato. Plant Pathol. Microbiol. 2013, 4, 49. [Google Scholar]
- Yorio, N.C.; Goins, G.D.; Kagie, H.R.; Wheeler, R.M.; Sager, J.C. Improving spinach radish and lettuce growth under red light-emitting diodes (LEDs) with blue light supplementation. Hort. Sci. 2001, 36, 380–383. [Google Scholar]
- Whitelam, G.; Halliday, K. Light and Plant Development; Blackwell: Oxford, UK, 2007. [Google Scholar]
- Matsuda, R.; Ohashi-Kaneko, K.; Fujiwara, K.; Kurata, K. Effects of blue light deficiency on acclimation of light energy partitioning in PSII and CO2 assimilation capacity to high irradiance in spinach leaves. Plant Cell Physiol 2008, 49, 664–670. [Google Scholar]
- Matsuda, R.; Ohashi-Kaneko, K.; Fujiwara, K.; Goto, E.; Kurata, K. Photosynthetic characteristics of rice leaves grown under red light with or without supplemental blue light. Plant Cell Physiol. 2004, 45, 1870–1874. [Google Scholar]
- Kang, J.H.; Krishnkumar, S.; Sua Atulba, S.L.; Jeong, B.R.; Hwang, S.J. Light intensity and photoperiod influence the growth and development of hydroponically grown leaf lettuce in a closed-type plant factory system. Hort. Environ. Biotechnol. 2013, 54, 501–509. [Google Scholar]
- Sun, J.D.; Nishio, J.N.; Vogelmann, T.C. Green light drives CO2 fixation deep within leaves. Plant Cell Physiol. 1998, 39, 1020–1026. [Google Scholar]
- Frechilla, S.; Talbott, L.D.; Bogomolni, R.A.; Zeiger, E. Reversal of blue light-stimulated stomatal opening by green light. Plant Physiol 2000, 41, 71–176. [Google Scholar]
- Terashima, I.; Fujita, T.; Inoue, T.; Chow, W.S.; Oguchi, R. Green light drives leaf photosynthesis more efficiently than red light in strong white light: Revisiting the enigmatic question of why leaves are green. Plant Cell Physiol. 2009, 50, 684–697. [Google Scholar]
- Olle; Margit; Virsile, A. The effects of light emitting diode on greenhouse plant growth and quality. Agric. Food Sci. 2013, 22, 223–234. [Google Scholar]
- Folta, K.M.; Maruhnich, S.A. Green light: A signal to slow down or stop. J. Exp. Bot. 2007, 58, 3099–3111. [Google Scholar]
- Fan, X.X.; Xu, Z.G.; Liu, X.Y.; Tang, C.M.; Wang, L.W.; Han, X.L. Effect of light intensity on the growth and leaf development of young tomato plants grown under a combination of red and blue light. Sci. Hort. 2013, 153, 50–55. [Google Scholar]
- Lin, K.H.; Huang, M.Y.; Huang, W.D.; Hsu, M.H.; Yang, Z.W.; Yang, C.M. The effects of red blue and white light-emitting diodes on the growth development and edible quality of hydroponically grown lettuce (Lactuca sativa L Var Capitata). Sci. Hort. 2013, 150, 86–91. [Google Scholar]
- Buschmann, C.; Meier, D.; Kleudgen, H.K.; Lichtenthaler, H.K. Regulation of chloroplast development by red and blue light. Photochem. Biol. 1978, 27, 195–198. [Google Scholar]
- Leong, T.Y.; Anderson, J.M. Effect of light quality on the composition and function of thylakoid membranes in Atriplex triangularis. Biochem. Biophys. Acta 1984, 766, 533–541. [Google Scholar]
- Eskins, K.; Jiang, C.Z.; Shibles, R. Light quality and irriadiance effects on pigments light harvesting proteins and Rubsico activity in a chlorophyll and light harvesting deficient soybean mutant. Physiol. Plant 1991, 83, 47–53. [Google Scholar]
- Raju, S.; Shah, S.; Gajbhiye, N. Effect of light intensity on photosynthesis and accumulation of sennosides in plant parts of senna (Cassia angustifolia Vahl). Indian J. Plant Physiol. 2013, 3, 285–289. [Google Scholar]
- Avala, J.A.; Ravetta, D.A. Allocation of photoassimilates to biomass resin and carbohydrate in Grindelia chiloensis as affected by light intensity. J. Field Crop. Res. 2001, 69, 143–149. [Google Scholar]
- Li, H.M.; Xu, Z.G.; Tang, C.M. Effect of light emitting diodes on growth and morphogenesis of upland cotton (Gossypium hirsutum L) plantlets in vitro. Plant Cell Tissue Organ Cult. 2010, 103, 155–163. [Google Scholar]
- Hogewoning, S.W.; Trouwborst, G.; Maljaars, H.; Poorter, H.; Leperen, W.V.; Jeremy, H. Blue light dose-responses of leaf photosynthesis morphology and chemical composition of Cucumis sativus grown under different combinations of red and blue light. J. Exp. Bot. 2010, 61, 3107–3117. [Google Scholar]
- Johkan, M.; Shoji, K.; Goto, F.; Hashida, S.; Yoshihara, T. Blue light-emitting diode light irradiation of seedlings improves seedling quality and growth after transplanting in red leaf lettuce. Hort. Sci. 2010, 45, 1809–1814. [Google Scholar]
- Ohashi-Kaneko, K.; Matsuda, R.; Goto, E.; Fujiwara, K.; Kurata, K. Growth of rice plants under red light with or without supplemental blue light. Soil Sci. Plant Nutr. 2006, 52, 444–452. [Google Scholar]
- Kim, H.H.; Goins, G.D.; Wheeler, R.M.; Sager, J.C. Green-light supplementation for enhanced lettuce growth under red- and blue-light-emitting diodes. Hort. Sci. 2004, 39, 1617–1622. [Google Scholar]
- Timperio, A.M.; D’Amici, G.M.; Barta, C.; Francesco, L.; Zolla, L. Proteomics pigment composition and organization of thylakoid membranes in iron-deficient spinach leaves. J. Exp. Bot. 2007, 58, 3695–3710. [Google Scholar]
- Hattori, T.; Sonobe, K.; Inanaga, S.; An, P.; Wataru, T.; Hideki, A.; Eneji, E.A.; Shigenori, M. Short term stomatal response to light intensity changes and osmotic stress in sorghum seedlings raised with and without silicon. Environ. Exp. Bot. 2007, 60, 177–182. [Google Scholar]
- Kim, H.H.; Wheeler, R.M.; Sager, J.C.; Goins, G.D.; Norikane, J.H. Evaluation of lettuce growth using supplemental green light with red and blue light-emitting diodes in a controlled environment—A review of research at Kennedy Space Center. Acta Hort. 2006, 711, 111–119. [Google Scholar]
- Fernandez, V.; Eichert, T.; del Rio, V.; Lopez-Casado, G.; Heredia-Guerrero, J.A.; Anunciacion, A.; Heredia, A.; Abadia, J. Leaf structural changes associated with iron deficiency chlorosis in field-grown pear and peach: Physiological implications. Plant Soil 2008, 311, 161–172. [Google Scholar]
- Hippler, M.; Klein, J.; Fink, A.; Allinger, T.; Hoerth, P. Towards functional proteomics of membrane protein complexes: analysis of thylakoid membranes from Chlamydomonas reinhardtii. Plant J. 2001, 28, 595–606. [Google Scholar]
- Stitt, M.; Schulze, D. Does RuBisCO control the rate of photosynthesis and plant growth? An exercise in molecular Ecophysiology. Plant Cell Environ. 1994, 17, 465–487. [Google Scholar]
- Imsande, J. Iron-sulfur clusters: Formation perturbation and physiological functions. Plant Physiol. Biochem. 1999, 37, 87–97. [Google Scholar]
- Briat, J.F.; Curie, C.; Gaymard, F. Iron utilization and metabolism in plants. Curr. Opin. Plant Biol. 2007, 10, 276–282. [Google Scholar]
- Kraty, B.A. Three non-circulating hydroponic methods for growing lettuce. Acta Hort. 2009, 843, 65–72. [Google Scholar]
- Kugler, M.; Jansch, L.; Kruft, V.; Schmitz, U.K.; Braun, H.P. Analysis of the chloroplast protein complexes by blue native polyacrylamide gel electrophoresis (BN-PAGE). Photosyn. Res. 1997, 53, 35–44. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Light intensity (μmol m−2 s−1) | Stomatal density (Mean ± SE, mm2) | ||
|---|---|---|---|
| Green LEDs | Red LEDs | Blue LEDs | |
| 70 | 0.7 ± 0.11 | ||
| 180 | 1.0 ± 0.11 | ||
| 88 | 1.4 ± 0.11 | ||
| 238 | 1.85 ± 0.11 | ||
| 80 | 1.7 ± 0.11 | ||
| 238 | 2.3 ± 0.11 | ||
| Light sources | Light intensity (μmol m−2 s−1) | Peak wave length λp (nm) |
|---|---|---|
| Green | 70 | 522 |
| Green | 180 | 522 |
| Red | 88 | 639 |
| Red | 238 | 639 |
| Blue | 80 | 470 |
| Blue | 238 | 470 |
© 2014 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Muneer, S.; Kim, E.J.; Park, J.S.; Lee, J.H. Influence of Green, Red and Blue Light Emitting Diodes on Multiprotein Complex Proteins and Photosynthetic Activity under Different Light Intensities in Lettuce Leaves (Lactuca sativa L.). Int. J. Mol. Sci. 2014, 15, 4657-4670. https://doi.org/10.3390/ijms15034657
Muneer S, Kim EJ, Park JS, Lee JH. Influence of Green, Red and Blue Light Emitting Diodes on Multiprotein Complex Proteins and Photosynthetic Activity under Different Light Intensities in Lettuce Leaves (Lactuca sativa L.). International Journal of Molecular Sciences. 2014; 15(3):4657-4670. https://doi.org/10.3390/ijms15034657
Chicago/Turabian StyleMuneer, Sowbiya, Eun Jeong Kim, Jeong Suk Park, and Jeong Hyun Lee. 2014. "Influence of Green, Red and Blue Light Emitting Diodes on Multiprotein Complex Proteins and Photosynthetic Activity under Different Light Intensities in Lettuce Leaves (Lactuca sativa L.)" International Journal of Molecular Sciences 15, no. 3: 4657-4670. https://doi.org/10.3390/ijms15034657
APA StyleMuneer, S., Kim, E. J., Park, J. S., & Lee, J. H. (2014). Influence of Green, Red and Blue Light Emitting Diodes on Multiprotein Complex Proteins and Photosynthetic Activity under Different Light Intensities in Lettuce Leaves (Lactuca sativa L.). International Journal of Molecular Sciences, 15(3), 4657-4670. https://doi.org/10.3390/ijms15034657