Dimers of G-Protein Coupled Receptors as Versatile Storage and Response Units

Abstract
:1. Introduction
2. Results and Discussion
2.1. On the Chances of GPCR Homo- and Heterodimerization
2.2. Some Notes Related to Methods and Paradigms
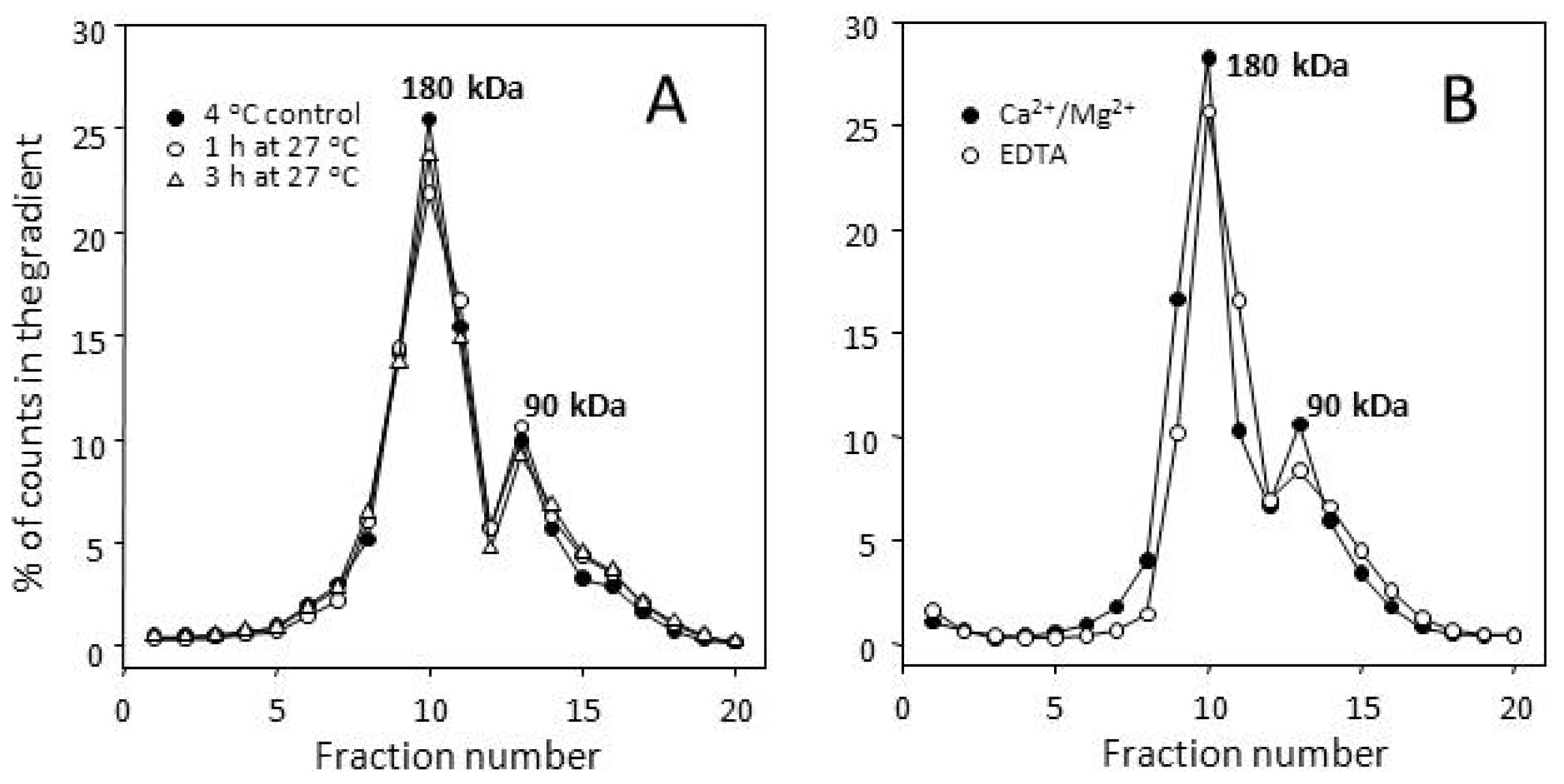
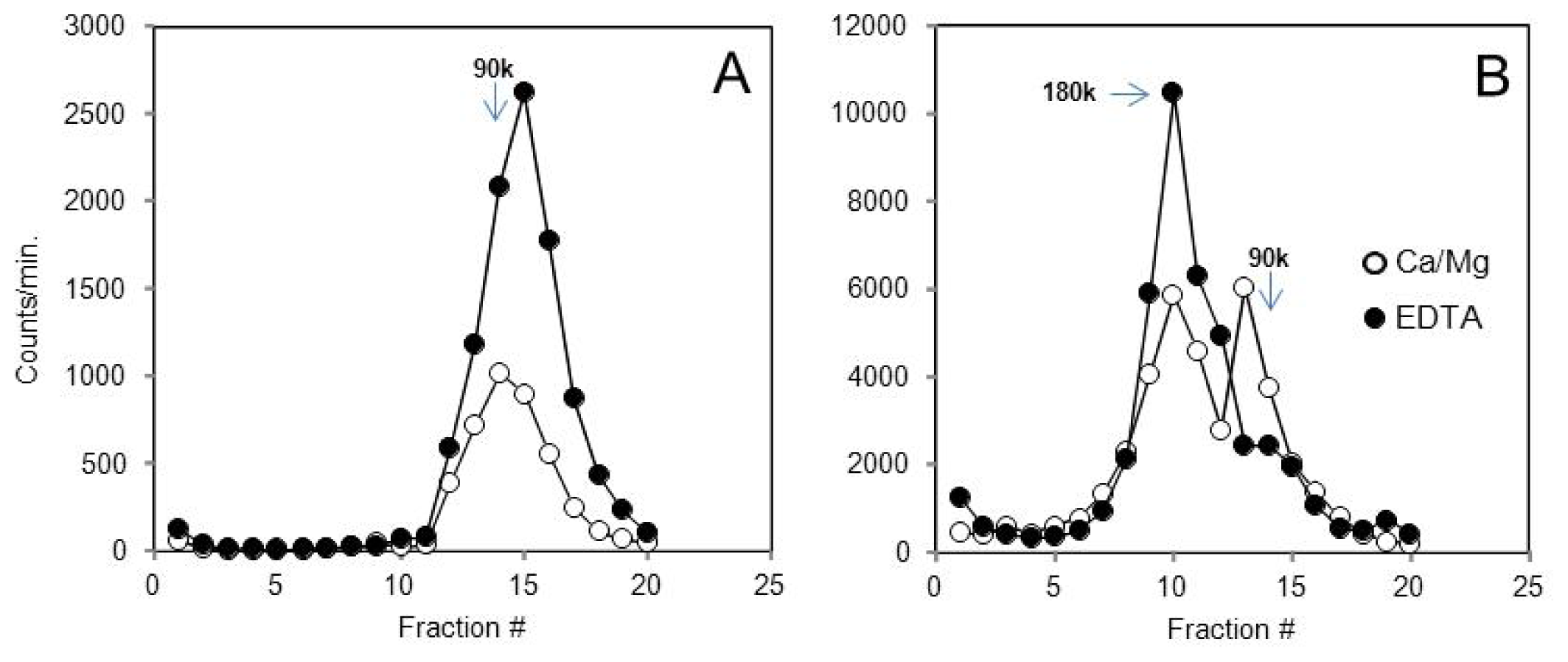
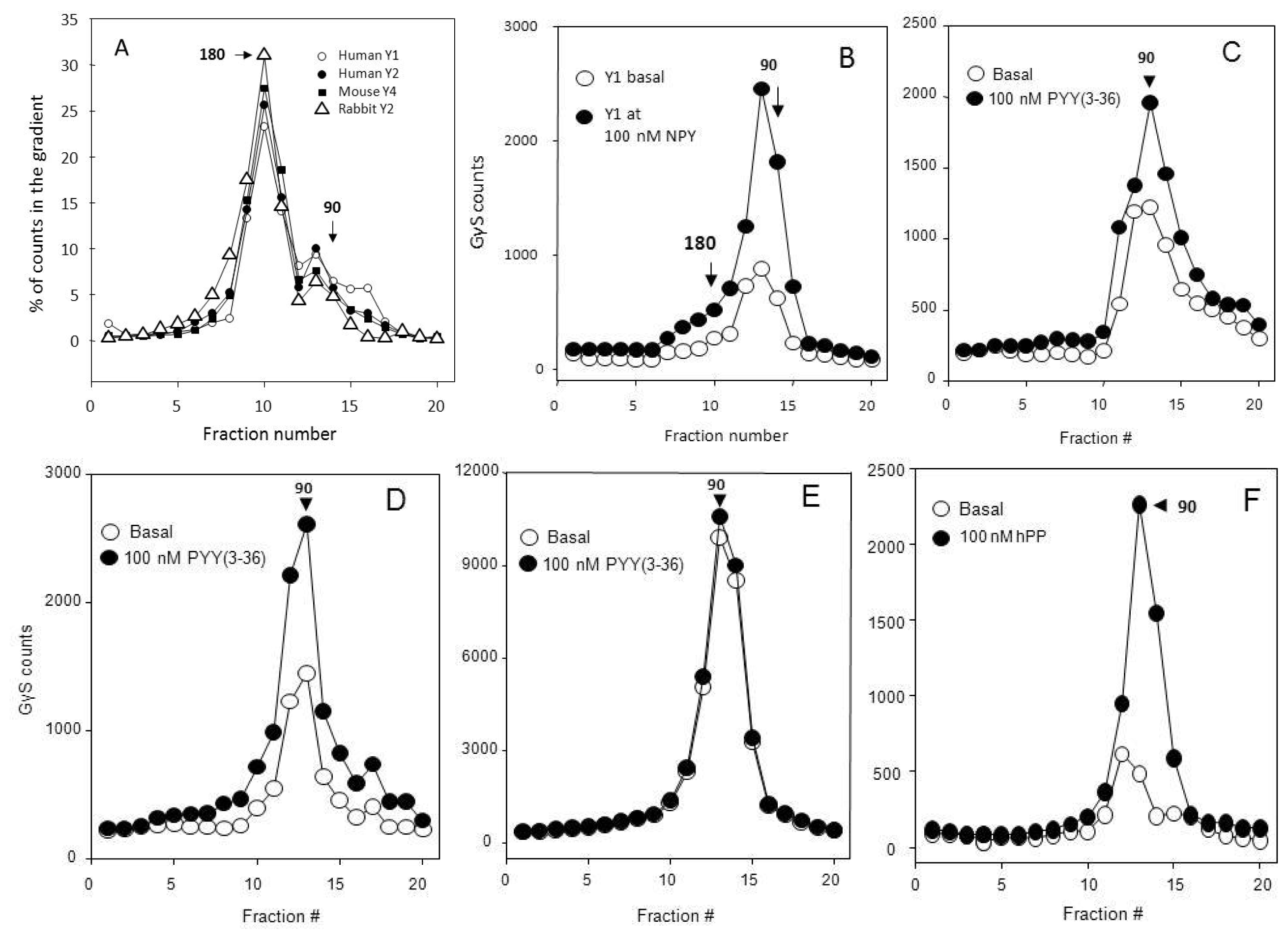
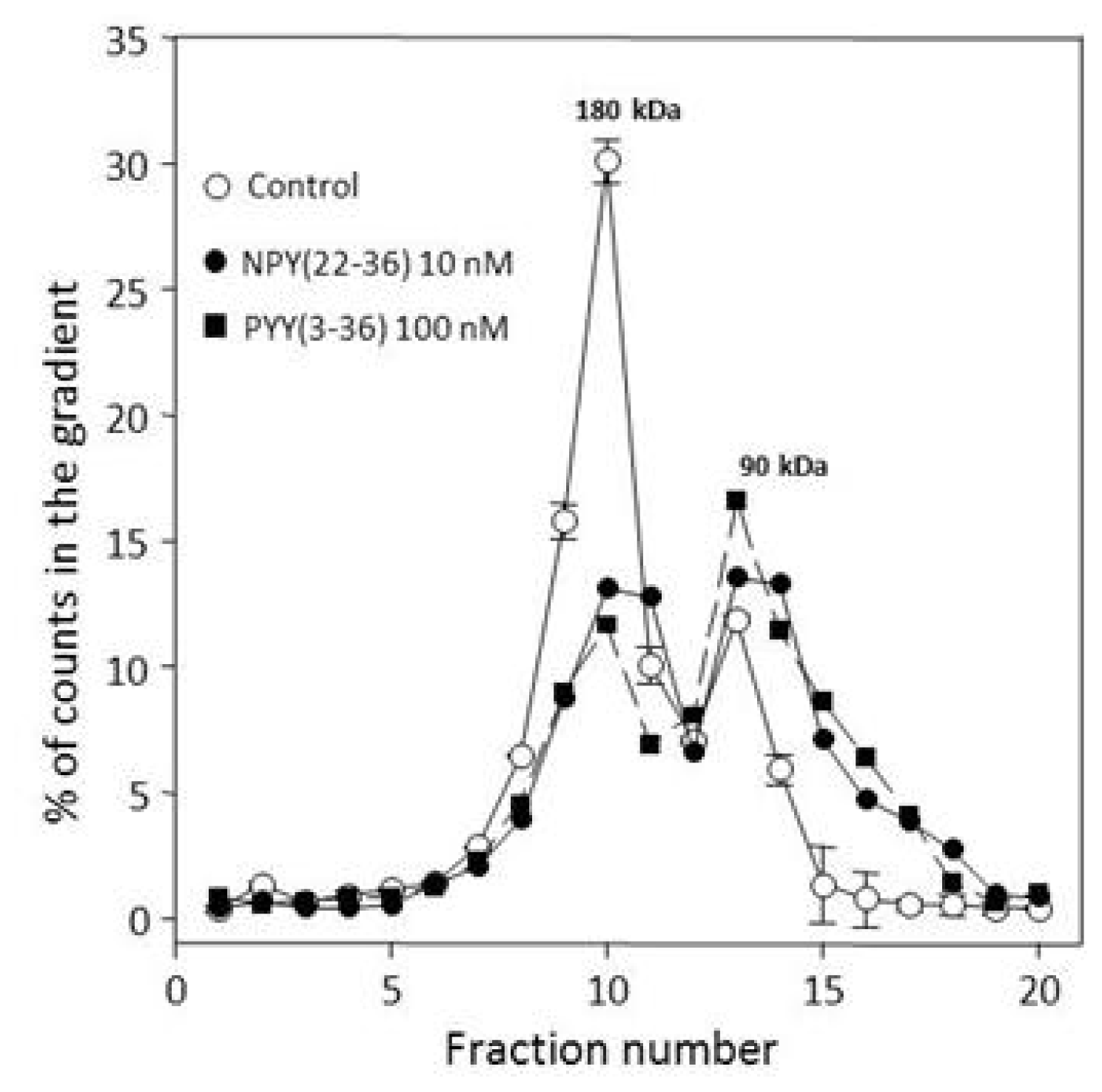
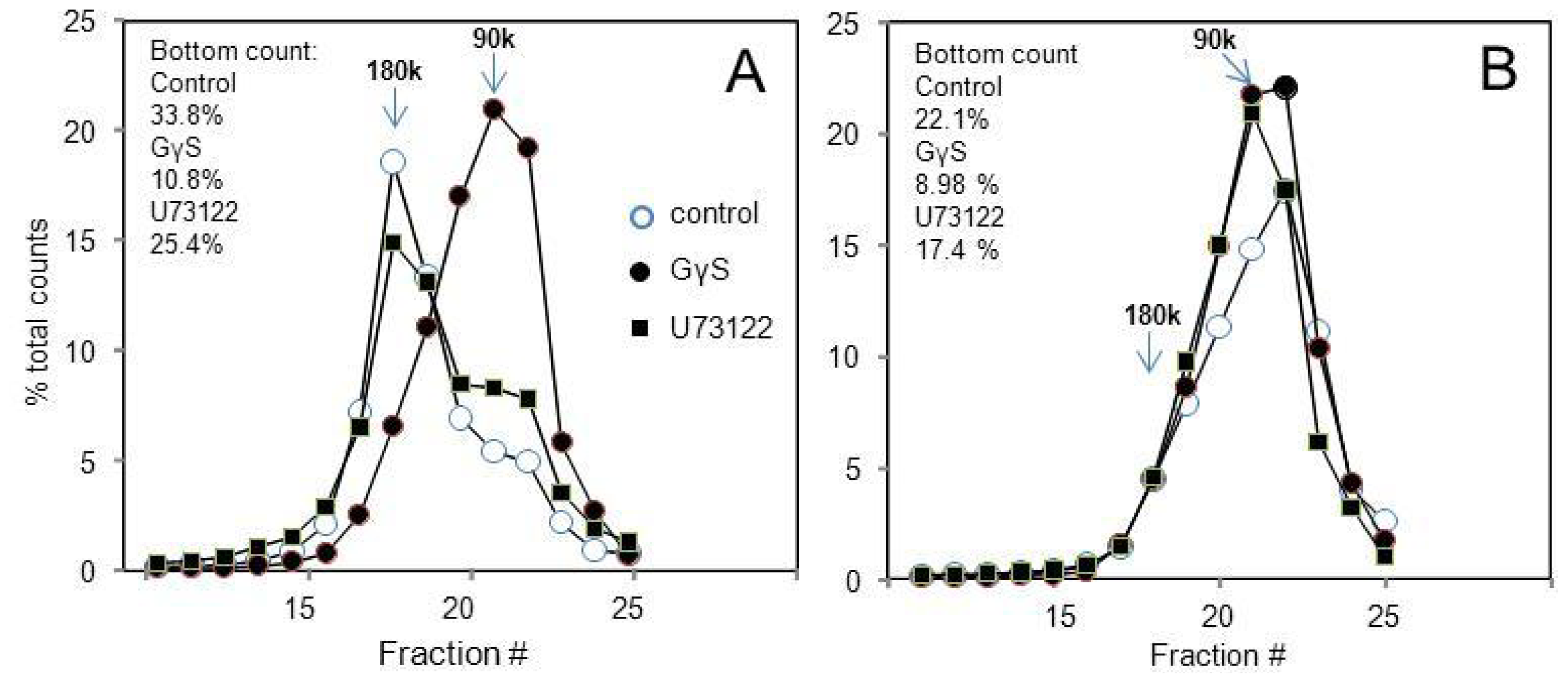
2.3. Many Peptide Receptor Dimers Can Be Studied Using the Binding of Native Agonists
2.4. Aggregative Clustering Aided by Arrestins Should Help GPCR Cycling
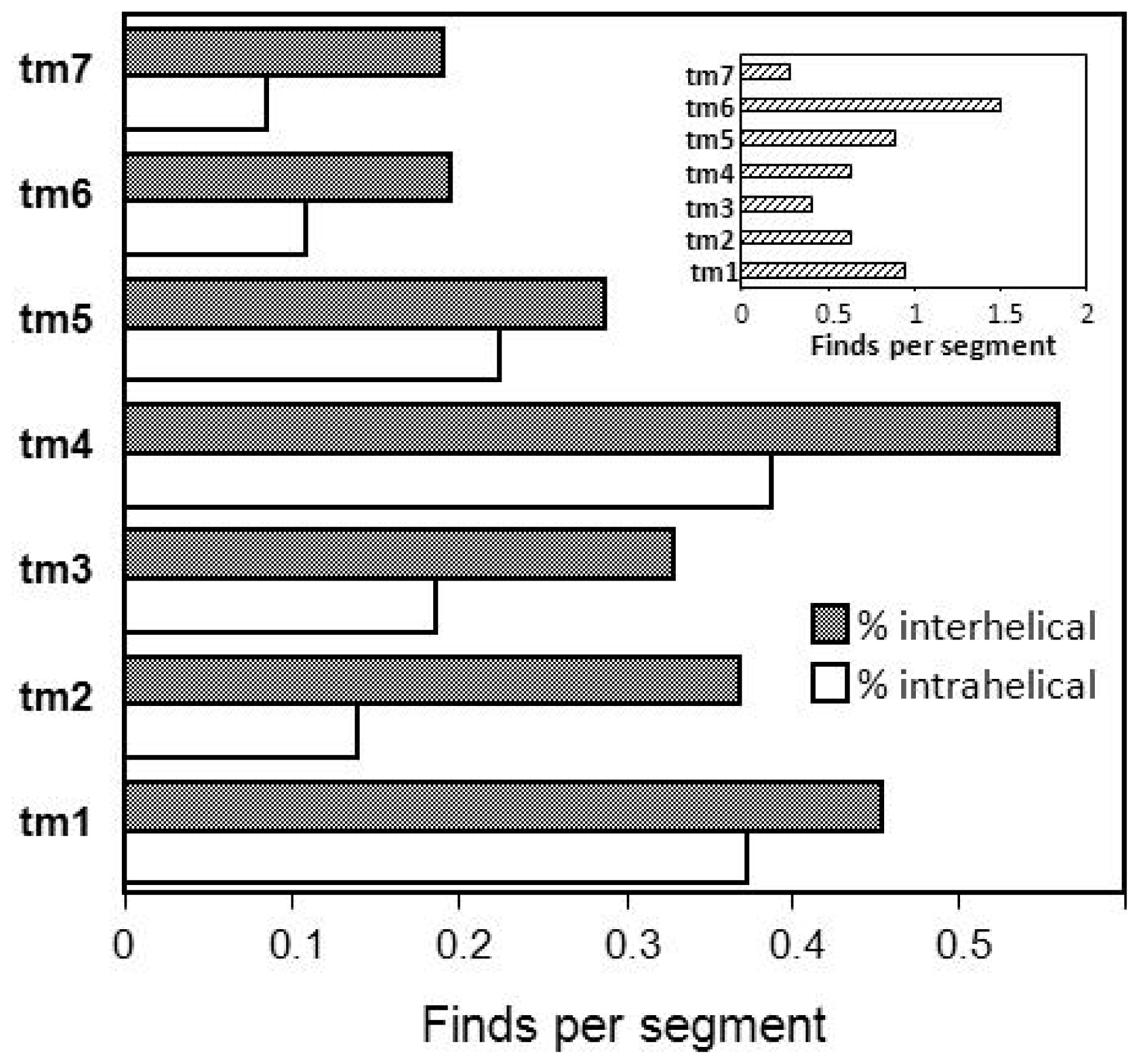
2.5. Hydrophobic Transmembrane Motifs and GPCR Dimerization
2.6. Association of Intracellular Basic Clusters with Transducers Could Support Dimerization of GPCRs
2.7. The Transductionally Stable C-GPCR Dimers Point to Possibly Similar Design in Other GPCR Groups
3. Experimental Section
3.1. Materials
3.2. Receptor and Nucleotide Site Labeling
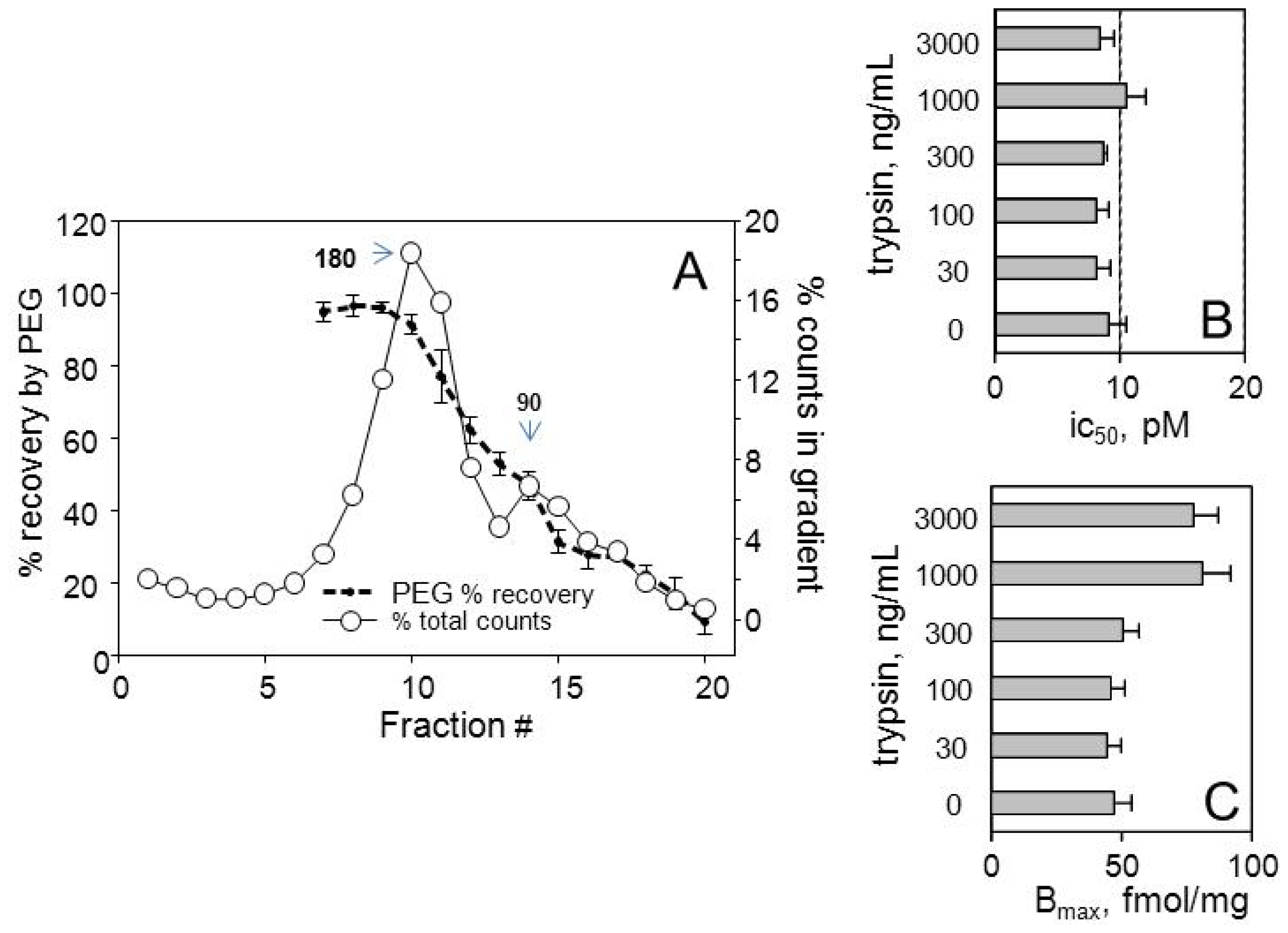
3.3. Characterization of Receptors in Density Gradients
3.4. Immunodetection of G-Protein α-Subunits Coupled to Receptors
4. Conclusions
Acknowledgments
Conflicts of Interest
References
- Palczewski, K.; Kumasaka, T.; Hori, T.; Behnke, C.A.; Motoshima, H.; Fox, B.A.; Le Trong, I.; Teller, D.C.; Okada, T.; Stenkamp, R.E.; et al. Crystal structure of rhodopsin: A G protein-coupled receptor. Science 2000, 289, 739–745. [Google Scholar]
- Fredriksson, R.; Lagerstrom, M.C.; Lundin, L.G.; Schioth, H.B. The G-protein-coupled receptors in the human genome form five main families Phylogenetic analysis paralogon groups and fingerprints. Mol. Pharmacol. 2003, 63, 1256–1272. [Google Scholar]
- Guo, W.; Shi, L.; Filizola, M.; Weinstein, H.; Javitch, J.A. Crosstalk in G-protein-coupled receptors: Changes at the transmembrane homodimer interface determine activation. Proc. Natl. Acad. Sci. USA 2005, 102, 17495–17500. [Google Scholar]
- Kota, P.; Reeves, P.J.; Rajbhandary, U.L.; Khorana, H.G. Opsin is present as dimers in COS1 cells: Identification of amino acids at the dimeric interface. Proc. Natl. Acad. Sci. USA 2006, 103, 3054–3059. [Google Scholar]
- Johnston, J.M.; Wang, H.; Provasi, D.; Filizola, M. Assessing the relative stability of dimer interfaces in G-protein-coupled receptors. PLoS Comput. Biol. 2012, 8, e1002649. [Google Scholar]
- Yasuda, D.; Okuno, T.; Yokomizo, T.; Hori, T.; Hirota, N.; Hashidate, T.; Miyano, M.; Shimizu, T.; Nakamura, M. Helix 8 of leukotriene B4 type-2 receptor is required for the folding to pass the quality control in the endoplasmic reticulum. FASEB J. 2009, 23, 1470–1481. [Google Scholar]
- Lee, S.J.; Montell, C. Regulation of the rhodopsin protein phosphatase RDGC through interaction with calmodulin. Neuron 2001, 32, 1097–1106. [Google Scholar]
- Duchene, J.; Schanstra, J.P.; Pecher, C.; Pizard, A.; Susini, C.; Esteve, J.P.; Bascands, J.L.; Girolami, J.P. A novel protein-protein interaction between a G-protein-coupled receptor and the phosphatase SHP-2 is involved in bradykinin-induced inhibition of cell proliferation. J. Biol. Chem. 2002, 277, 40375–40383. [Google Scholar]
- Hammad, M.M.; Kuang, Y.Q.; Yan, R.; Allen, H.; Dupre, D.J. Na+/H+ exchanger regulatory factor-1 is involved in chemokine receptor homodimer CCR5 internalization and signal transduction but does not affect CXCR4 homodimer or CXCR4-CCR5 heterodimer. J. Biol. Chem. 2010, 285, 34653–34664. [Google Scholar]
- Parker, M.S.; Sah, R.; Sheriff, S.; Balasubramaniam, A.; Parker, S.L. Internalization of cloned pancreatic polypeptide receptors is accelerated by all types of Y4 agonists. Regul. Pept. 2005, 132, 91–101. [Google Scholar]
- Gray, N.W.; Fourgeaud, L.; Huang, B.; Chen, J.; Cao, H.; Oswald, B.J.; Hemar, A.; McNiven, M.A. Dynamin 3 is a component of the postsynapse where it interacts with mGluR5 and Homer. Curr. Biol. 2003, 13, 510–515. [Google Scholar]
- Christopoulos, A.; Christopoulos, G.; Morfis, M.; Udawela, M.; Laburthe, M.; Couvineau, A.; Kuwasako, K.; Tilakaratne, N.; Sexton, P.M. Novel receptor partners and function of receptor activity-modifying proteins. J. Biol. Chem. 2003, 278, 3293–3297. [Google Scholar]
- Heroux, M.; Breton, B.; Hogue, M.; Bouvier, M. Assembly and signaling of CRLR and RAMP1 complexes assessed by BRET. Biochemistry 2007, 46, 7022–7033. [Google Scholar]
- Roy, S.J.; Glazkova, I.; Frechette, L.; Iorio-Morin, C.; Binda, C.; Petrin, D.; Trieu, P.; Robitaille, M.; Angers, S.; Hebert, T.E.; et al. Novel gel-free proteomics approach identifies RNF5 and JAMP as modulators of GPCR stability. Mol. Endocrinol. 2013, 27, 1245–1266. [Google Scholar]
- Parker, S.L.; Parker, M.S. Ligand association with the rabbit kidney and brain Y1 Y2 and Y5-like neuropeptide Y (NPY) receptors shows large subtype-related differences in sensitivity to chaotropic and alkylating agents. Regul. Pept. 2000, 87, 59–72. [Google Scholar]
- Parker, S.L.; Parker, M.S.; Sah, R.; Sallee, F.R.; Balasubramaniam, A. Parallel inactivation of Y2 receptor and G-proteins in CHO cells by pertussis toxin. Regul. Pept. 2007, 139, 128–135. [Google Scholar]
- Baneres, J.L.; Parello, J. Structure-based analysis of GPCR function: Evidence for a novel pentameric assembly between the dimeric leukotriene B4 receptor BLT1 and the G-protein. J. Mol. Biol. 2003, 329, 815–829. [Google Scholar]
- Parker, M.S.; Sah, R.; Balasubramaniam, A.; Sallee, F.R.; Park, E.A.; Sweatman, T.; Parker, S.L. Dimers of the neuropeptide Y (NPY) Y2 receptor show asymmetry in agonist affinity and association with G-proteins. J. Recept. Signal Transduct. 2008, 28, 437–451. [Google Scholar]
- Dinger, M.C.; Bader, J.E.; Kobor, A.D.; Kretzschmar, A.K.; Beck-Sickinger, A.G. Homodimerization of neuropeptide y receptors investigated by fluorescence resonance energy transfer in living cells. J. Biol. Chem. 2003, 278, 10562–10571. [Google Scholar]
- Berglund, M.M.; Schober, D.A.; Statnick, M.A.; McDonald, P.H.; Gehlert, D.R. The use of bioluminescence resonance energy transfer 2 to study neuropeptide Y receptor agonist-induced beta-arrestin 2 interaction. J. Pharmacol. Exp. Ther. 2003, 306, 147–156. [Google Scholar]
- Fotiadis, D.; Liang, Y.; Filipek, S.; Saperstein, D.A.; Engel, A.; Palczewski, K. Atomic-force microscopy: Rhodopsin dimers in native disc membranes. Nature 2003, 421, 127–128. [Google Scholar]
- Liang, Y.; Fotiadis, D.; Filipek, S.; Saperstein, D.A.; Palczewski, K.; Engel, A. Organization of the G-protein-coupled receptors rhodopsin and opsin in native membranes. J. Biol. Chem. 2003, 278, 21655–21662. [Google Scholar]
- Jastrzebska, B.; Maeda, T.; Zhu, L.; Fotiadis, D.; Filipek, S.; Engel, A.; Stenkamp, R.E.; Palczewski, K. Functional characterization of rhodopsin monomers and dimers in detergents. J. Biol. Chem. 2004, 279, 54663–54675. [Google Scholar]
- Parker, S.L.; Parker, M.S.; Estes, A.M.; Wong, Y.Y.; Sah, R.; Sweatman, T.; Park, E.A.; Balasubramaniam, A.; Sallee, F.R. The neuropeptide Y (NPY) Y2 receptors are largely dimeric in the kidney but monomeric in the forebrain. J. Recept. Signal. Transduct. 2008, 28, 245–263. [Google Scholar]
- Whorton, M.R.; Bokoch, M.P.; Rasmussen, S.G.; Huang, B.; Zare, R.N.; Kobilka, B.; Sunahara, R.K. A monomeric G-protein-coupled receptor isolated in a high-density lipoprotein particle efficiently activates its G-protein. Proc. Natl. Acad. Sci. USA 2007, 104, 7682–7687. [Google Scholar]
- Dupre, D.J.; Chen, Z.; Le Gouill, C.; Theriault, C.; Parent, J.L.; Rola-Pleszczynski, M.; Stankova, J. Trafficking ubiquitination and down-regulation of the human platelet-activating factor receptor. J. Biol. Chem. 2003, 278, 48228–48235. [Google Scholar]
- Qian, R.; Chen, C.; Colley, K.J. Location and mechanism of alpha 26-sialyltransferase dimer formation Role of cysteine residues in enzyme dimerization localization activity and processing. J. Biol. Chem. 2001, 276, 28641–28649. [Google Scholar]
- Parker, S.L.; Parker, M.S.; Sallee, F.R.; Balasubramaniam, A. Oligomerization of neuropeptide Y (NPY) Y2 receptors in CHO cells depends on functional pertussis toxin-sensitive G-proteins. Regul. Pept. 2007, 144, 72–81. [Google Scholar]
- Parker, S.L.; Parker, M.S.; Sah, R.; Balasubramaniam, A.; Sallee, F.R. Pertussis toxin induces parallel loss of neuropeptide Y Y(1) receptor dimers and G(i) alpha subunit function in CHO cells. Eur. J. Pharmacol. 2008, 579, 13–25. [Google Scholar]
- Estes, A.M.; McAllen, K.; Parker, M.S.; Sah, R.; Sweatman, T.; Park, E.A.; Balasubramaniam, A.; Sallee, F.R.; Walker, M.W.; Parker, S.L. Maintenance of Y receptor dimers in epithelial cells depends on interaction with G-protein heterotrimers. Amino Acids 2011, 40, 371–380. [Google Scholar]
- Fotiadis, D.; Liang, Y.; Filipek, S.; Saperstein, D.A.; Engel, A.; Palczewski, K. The G protein-coupled receptor rhodopsin in the native membrane. FEBS Lett. 2004, 564, 281–288. [Google Scholar]
- Sah, R.; Balasubramaniam, A.; Parker, M.S.; Sallee, F.; Parker, S.L. Neuropeptide Y as a partial agonist of the Y1 receptor. Eur. J. Pharmacol. 2005, 525, 60–68. [Google Scholar]
- Jastrzebska, B.; Fotiadis, D.; Jang, G.F.; Stenkamp, R.E.; Engel, A.; Palczewski, K. Functional and structural characterization of rhodopsin oligomers. J. Biol. Chem. 2006, 281, 11917–11922. [Google Scholar]
- Whorton, M.R.; Jastrzebska, B.; Park, P.S.; Fotiadis, D.; Engel, A.; Palczewski, K.; Sunahara, R.K. Efficient coupling of transducin to monomeric rhodopsin in a phospholipid bilayer. J. Biol. Chem. 2008, 283, 4387–4394. [Google Scholar]
- Arcemisbehere, L.; Sen, T.; Boudier, L.; Balestre, M.N.; Gaibelet, G.; Detouillon, E.; Orcel, H.; Mendre, C.; Rahmeh, R.; Granier, S.; et al. Leukotriene BLT2 receptor monomers activate the G(i2) GTP-binding protein more efficiently than dimers. J. Biol. Chem. 2011, 285, 6337–6347. [Google Scholar]
- Jiang, H.; Kuang, Y.; Wu, Y.; Smrcka, A.; Simon, M.I.; Wu, D. Pertussis toxin-sensitive activation of phospholipase C by the C5a and fMet-Leu-Phe receptors. J. Biol. Chem. 1996, 271, 13430–13434. [Google Scholar]
- Fredriksson, R.; Schioth, H.B. The repertoire of G-protein-coupled receptors in fully sequenced genomes. Mol. Pharmacol. 2005, 67, 1414–1425. [Google Scholar]
- McCann, S.M.; Ojeda, S.R.; Fawcett, C.P.; Krulich, L. Catecholaminergic control of gonadotropin and prolactin secretion with particular reference to the possible participation of dopamine. Adv. Neurol. 1974, 5, 435–445. [Google Scholar]
- Kalra, S.P.; Allen, L.G.; Sahu, A.; Kalra, P.S.; Crowley, W.R. Gonadal steroids and neuropeptide Y-opioid-LHRH axis: Interactions and diversities. J. Steroid Biochem. 1988, 30, 185–193. [Google Scholar]
- Wirmer, A.; Bradler, S.; Heinrich, R. Homology of insect corpora allata and vertebrate adenohypophysis? Arthropod Struct. Dev. 2012, 41, 409–417. [Google Scholar]
- Pierobon, P. Coordinated modulation of cellular signaling through ligand-gated ion channels in Hydra vulgaris (Cnidaria Hydrozoa). Int. J. Dev. Biol. 2012, 56, 551–565. [Google Scholar]
- Periole, X.; Knepp, A.M.; Sakmar, T.P.; Marrink, S.J.; Huber, T. Structural determinants of the supramolecular organization of G-protein-coupled receptors in bilayers. J. Am. Chem. Soc. 2012, 134, 10959–10965. [Google Scholar]
- Huang, J.; Chen, S.; Zhang, J.J.; Huang, X.Y. Crystal structure of oligomeric beta1-adrenergic G-protein-coupled receptors in ligand-free basal state. Nat. Struct. Mol. Biol. 2013, 20, 419–425. [Google Scholar]
- Sheikh, S.P.; Sheikh, M.I.; Schwartz, T.W. Y2-type receptors for peptide YY on renal proximal tubular cells in the rabbit. Am. J. Physiol. 1989, 257, F978–F984. [Google Scholar]
- Brann, M.R.; Young, W.S., 3rd. Localization and quantitation of opsin and transducin mRNAs in bovine retina by in situ hybridization histochemistry. FEBS Lett. 1986, 200, 275–278. [Google Scholar]
- Hilal-Dandan, R.; Villegas, S.; Gonzalez, A.; Brunton, L.L. The quasi-irreversible nature of endothelin binding and G-protein-linked signaling in cardiac myocytes. J. Pharmacol. Exp. Ther. 1997, 281, 267–273. [Google Scholar]
- Hilton, J.M.; Dowton, M.; Houssami, S.; Sexton, P.M. Identification of key components in the irreversibility of salmon calcitonin binding to calcitonin receptors. J. Endocrinol. 2000, 166, 213–226. [Google Scholar]
- Parker, M.S.; Berglund, M.M.; Lundell, I.; Parker, S.L. Blockade of pancreatic polypeptide-sensitive neuropeptide Y (NPY) receptors by agonist peptides is prevented by modulators of sodium transport Implications for receptor signaling and regulation. Peptides 2001, 22, 887–898. [Google Scholar]
- Brighton, P.J.; Szekeres, P.G.; Wise, A.; Willars, G.B. Signaling and ligand binding by recombinant neuromedin U receptors: Evidence for dual coupling to Galphaq/11 and Galphai and an irreversible ligand-receptor interaction. Mol. Pharmacol. 2004, 66, 1544–1556. [Google Scholar]
- Dautzenberg, F.M.; Neysari, S. Irreversible binding kinetics of neuropeptide Y ligands to Y2 but not to Y1 and Y5 receptors. Pharmacology 2005, 75, 21–29. [Google Scholar]
- Parker, S.L.; Parker, M.S.; Sah, R.; Balasubramaniam, A.; Sallee, F.R. Self-regulation of agonist activity at the Y receptors. Peptides 2007, 28, 203–213. [Google Scholar]
- Parker, M.S.; Sah, R.; Parker, S.L. Surface masking shapes the traffic of the neuropeptide Y Y2 receptor. Peptides 2012, 37, 40–48. [Google Scholar]
- Estes, A.M.; Wong, Y.Y.; Parker, M.S.; Sallee, F.R.; Balasubramaniam, A.; Parker, S.L. Neuropeptide Y (NPY) Y2 receptors of rabbit kidney cortex are largely dimeric. Regul. Pept. 2008, 150, 88–94. [Google Scholar]
- Okamoto, T.; Murayama, Y.; Hayashi, Y.; Inagaki, M.; Ogata, E.; Nishimoto, I. Identification of a Gs activator region of the beta 2-adrenergic receptor that is autoregulated via protein kinase A-dependent phosphorylation. Cell 1991, 67, 723–730. [Google Scholar]
- Laporte, S.A.; Oakley, R.H.; Holt, J.A.; Barak, L.S.; Caron, M.G. The interaction of beta-arrestin with the AP-2 adaptor is required for the clustering of beta 2-adrenergic receptor into clathrin-coated pits. J. Biol. Chem. 2000, 275, 23120–23126. [Google Scholar]
- Innamorati, G.; Le Gouill, C.; Balamotis, M.; Birnbaumer, M. The long and the short cycle Alternative intracellular routes for trafficking of G-protein-coupled receptors. J. Biol. Chem. 2001, 276, 13096–13103. [Google Scholar]
- Han, J.; Reddig, K.; Li, H.S. Prolonged Gq activity triggers fly rhodopsin endocytosis and degradation and reduces photoreceptor sensitivity. EMBO J. 2007, 26, 4966–4973. [Google Scholar]
- Adamian, L.; Jackups, R., Jr; Binkowski, T.A.; Liang, J. Higher-order interhelical spatial interactions in membrane proteins. J. Mol. Biol. 2003, 327, 251–272. [Google Scholar]
- Lopez-Gimenez, J.F.; Canals, M.; Pediani, J.D.; Milligan, G. The alpha1B-adrenoceptor exists as a higher-order oligomer: Effective oligomerization is required for receptor maturation surface delivery and function. Mol. Pharmacol. 2007, 71, 1015–1029. [Google Scholar]
- Kim, S.K.; Jacobson, K.A. Computational prediction of homodimerization of the A3 adenosine receptor. J. Mol. Graph. Model. 2006, 25, 549–561. [Google Scholar]
- Harikumar, K.G.; Dong, M.; Cheng, Z.; Pinon, D.I.; Lybrand, T.P.; Miller, L.J. Transmembrane segment peptides can disrupt cholecystokinin receptor oligomerization without affecting receptor function. Biochemistry 2006, 45, 14706–14716. [Google Scholar]
- Brosig, B.; Langosch, D. The dimerization motif of the glycophorin A transmembrane segment in membranes: Importance of glycine residues. Protein Sci. 1998, 7, 1052–1056. [Google Scholar]
- Li, R.; Gorelik, R.; Nanda, V.; Law, P.B.; Lear, J.D.; DeGrado, W.F.; Bennett, J.S. Dimerization of the transmembrane domain of Integrin alphaIIb subunit in cell membranes. J. Biol. Chem. 2004, 279, 26666–26673. [Google Scholar]
- Senes, A.; Gerstein, M.; Engelman, D.M. Statistical analysis of amino acid patterns in transmembrane helices: The GxxxG motif occurs frequently and in association with beta-branched residues at neighboring positions. J. Mol. Biol. 2000, 296, 921–936. [Google Scholar]
- Russ, W.P.; Engelman, D.M. The GxxxG motif: A framework for transmembrane helix-helix association. J. Mol. Biol. 2000, 296, 911–919. [Google Scholar]
- Uberti, M.A.; Hall, R.A.; Minneman, K.P. Subtype-specific dimerization of alpha 1-adrenoceptors: Effects on receptor expression and pharmacological properties. Mol. Pharmacol. 2003, 64, 1379–1390. [Google Scholar]
- Chakera, A.; Seeber, R.M.; John, A.E.; Eidne, K.A.; Greaves, D.R. The duffy antigen/receptor for chemokines exists in an oligomeric form in living cells and functionally antagonizes CCR5 signaling through hetero-oligomerization. Mol. Pharmacol. 2008, 73, 1362–1370. [Google Scholar]
- Harmar, A.J. Family-B G-protein-coupled receptors. Genome Biol. 2001, 2, 3013.1–3013.10. [Google Scholar]
- Parker, M.S.; Park, E.A.; Sallee, F.R.; Parker, S.L. Two intracellular helices of G-protein coupling receptors could generally support oligomerization and coupling with transducers. Amino Acids 2011, 40, 261–268. [Google Scholar]
- Baltoumas, F.A.; Theodoropoulou, M.C.; Hamodrakas, S.J. Interactions of the alpha-subunits of heterotrimeric G-proteins with GPCRs effectors and RGS proteins: A critical review and analysis of interacting surfaces conformational shifts structural diversity and electrostatic potentials. J. Struct. Biol. 2013, 182, 209–218. [Google Scholar]
- Ikezu, T.; Okamoto, T.; Ogata, E.; Nishimoto, I. Amino acids 356–372 constitute a Gi-activator sequence of the alpha 2-adrenergic receptor and have a Phe substitute in the G-protein-activator sequence motif. FEBS Lett. 1992, 311, 29–32. [Google Scholar]
- Cen, B.; Yu, Q.; Guo, J.; Wu, Y.; Ling, K.; Cheng, Z.; Ma, L.; Pei, G. Direct binding of beta-arrestins to two distinct intracellular domains of the delta opioid receptor. J. Neurochem. 2001, 76, 1887–1894. [Google Scholar]
- Mercier, J.F.; Salahpour, A.; Angers, S.; Breit, A.; Bouvier, M. Quantitative assessment of beta 1- and beta 2-adrenergic receptor homo- and heterodimerization by bioluminescence resonance energy transfer. J. Biol. Chem. 2002, 277, 44925–44931. [Google Scholar]
- Parent, A.; Hamelin, E.; Germain, P.; Parent, J.L. Rab11 regulates the recycling of the beta2-adrenergic receptor through a direct interaction. Biochem. J. 2009, 418, 163–172. [Google Scholar]
- Hu, J.; Hu, K.; Liu, T.; Stern, M.K.; Mistry, R.; Challiss, R.A.; Costanzi, S.; Wess, J. Novel structural and functional insights into m3 muscarinic receptor dimer/oligomer formation. J. Biol. Chem. 2013, 288, 34777–34790. [Google Scholar]
- Bessis, A.S.; Rondard, P.; Gaven, F.; Brabet, I.; Triballeau, N.; Prezeau, L.; Acher, F.; Pin, J.P. Closure of the Venus flytrap module of mGlu8 receptor and the activation process: Insights from mutations converting antagonists into agonists. Proc. Natl. Acad. Sci. USA 2002, 99, 11097–11102. [Google Scholar]
- Yanagawa, M.; Yamashita, T.; Shichida, Y. Comparative fluorescence resonance energy transfer analysis of metabotropic glutamate receptors: Implications about the dimeric arrangement and rearrangement upon ligand bindings. J. Biol. Chem. 2011, 286, 22971–22981. [Google Scholar]
- Moyle, W.R.; Campbell, R.K.; Myers, R.V.; Bernard, M.P.; Han, Y.; Wang, X. Co-evolution of ligand-receptor pairs. Nature 1994, 368, 251–255. [Google Scholar]
- Zhang, R.; Buczko, E.; Dufau, M.L. Requirement of cysteine residues in exons 1–6 of the extracellular domain of the luteinizing hormone receptor for gonadotropin binding. J. Biol. Chem. 1996, 271, 5755–5760. [Google Scholar]
- Ho, S.C.; Goh, S.S.; Li, S.; Khoo, D.H.; Paterson, M. Effects of mutations involving cysteine residues distal to the S281HCC motif at the C-terminus on the functional characteristics of a truncated ectodomain-only thyrotropin receptor anchored on glycosylphosphatidyl-inositol. Thyroid 2008, 18, 1313–1319. [Google Scholar]
- Urizar, E.; Montanelli, L.; Loy, T.; Bonomi, M.; Swillens, S.; Gales, C.; Bouvier, M.; Smits, G.; Vassart, G.; Costagliola, S. Glycoprotein hormone receptors: Link between receptor homodimerization and negative cooperativity. EMBO J. 2005, 24, 1954–1964. [Google Scholar]
- Allen, M.D.; Neumann, S.; Gershengorn, M.C. Occupancy of both sites on the thyrotropin (TSH) receptor dimer is necessary for phosphoinositide signaling. FASEB J. 2011, 25, 3687–3694. [Google Scholar]
- Parker, S.L.; Kane, J.K.; Parker, M.S.; Berglund, M.M.; Lundell, I.A.; Li, M.D. Cloned neuropeptide Y (NPY) Y1 and pancreatic polypeptide Y4 receptors expressed in Chinese hamster ovary cells show considerable agonist-driven internalization in contrast to the NPY Y2 receptor. Eur. J. Biochem. 2001, 268, 877–886. [Google Scholar]
- Hunyady, L. Molecular mechanisms of angiotensin II receptor internalization. J. Am. Soc. Nephrol. 1999, 10(Suppl 11), S47–S56. [Google Scholar]
- Faussner, A.; Proud, D.; Towns, M.; Bathon, J.M. Influence of the cytosolic carboxyl termini of human B1 and B2 kinin receptors on receptor sequestration ligand internalization and signal transduction. J. Biol. Chem. 1998, 273, 2617–2623. [Google Scholar]
- Nouel, D.; Gaudriault, G.; Houle, M.; Reisine, T.; Vincent, J.P.; Mazella, J.; Beaudet, A. Differential internalization of somatostatin in COS-7 cells transfected with SST1 and SST2 receptor subtypes: A confocal microscopic study using novel fluorescent somatostatin derivatives. Endocrinology 1997, 138, 296–306. [Google Scholar]
- Chu, P.; Murray, S.; Lissin, D.; von Zastrow, M. Delta and kappa opioid receptors are differentially regulated by dynamin-dependent endocytosis when activated by the same alkaloid agonist. J. Biol. Chem. 1997, 272, 27124–27130. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Molecule | Number of aa | aPCN count | % aPCN | bPCN count | % bPCN |
|---|---|---|---|---|---|
| Go | 354 | 2 | 1.98 | 3 | 3.39 |
| Gi1 | 354 | 3 | 3.11 | 4 | 4.24 |
| Gi2 | 355 | 3 | 3.10 | 3 | 3.10 |
| Gi3 | 354 | 3 | 3.11 | 4 | 4.24 |
| Gt1 | 350 | 2 | 2.29 | 5 | 6.29 |
| Gt2 | 354 | 2 | 2.26 | 4 | 5.09 |
| Gt3 | 354 | 2 | 2.26 | 3 | 3.39 |
| Gq | 353 | 4 | 4.82 | 3 | 3.12 |
| Gs | 394 | 2 | 1.52 | 2 | 2.03 |
| Golf | 381 | 1 | 0.787 | 4 | 4.2 |
| β-arrestin-1 | 418 | 4 | 4.07 | 6 | 6.70 |
| β-arrestin-2 | 409 | 4 | 4.16 | 6 | 6.85 |
| Domain | % Sequences with GXXXG | % ILMV in XXX | % FWY in XXX |
|---|---|---|---|
| tm1 | 2.84 | 40 | 0 |
| tm2 | 0.948 | 30 | 0 |
| tm3 | 4.74 | 16.4 | 25.5 |
| tm4 | 7.11 | 38.7 | 9.33 |
| tm5 | 3.79 | 35 | 7.50 |
| tm6 | 10.43 | 46.4 | 6.36 |
| tm7 | 0.474 | 40 | 0 |
| Glycophorins | 100 | 50 | 50 |
| Group | Mean # aa | DE % | HKR % | Homo-acidic % | Homo-basic % | aPCN % | bPCN % |
|---|---|---|---|---|---|---|---|
| G-protein subunits | |||||||
| Gα [17] | 362 | 15 | 15.5 | 27.1 | 27.9 | 2.30 | 3.88 |
| Gβ [5] | 351 | 11.6 | 12.0 | 28.3 | 20.0 | 0.15 | 1.70 |
| Gγ [11] | 71 | 13.9 | 15.5 | 25.3 | 20.8 | 0.54 | 0 |
| A-GPCRs (rhodopsin family) | |||||||
| ic1 [211] | 11.2 | 1.68 | 25.9 | 0.30 | 39.6 | 0 | 17.1 |
| ic2 [211] | 14.8 | 1.69 | 26.0 | 0.50 | 58.1 | 0 | 10.8 |
| ic3 [211] | 42.9 | 5.56 | 27.5 | 5.22 | 62.0 | 0.64 | 13.2 |
| ic3 C-terminal 13 aa [211] | 13.0 | 5.90 | 35.5 | 1.87 | 62.1 | 0.51 | 17.3 |
| ic3 less C-terminal 13 aa [207] | 31.4 | 4.88 | 21.4 | 5.32 | 42.5 | 0.52 | 6.66 |
| ic4 [211] | 56.5 | 9.23 | 18.4 | 12.3 | 44.1 | 1.26 | 4.73 |
| H8 [210] | 14.0 | 6.22 | 24.5 | 1.13 | 41.3 | 0.10 | 6.49 |
| ic4 past H8 [208] | 39.5 | 9.72 | 16.1 | 14.8 | 30.9 | 1.51 | 3.91 |
| Ribosomal proteins | |||||||
| 60S ribosomal proteins [49] | 168 | 7.27 | 28.2 | 8.00 | 66.2 | 0.85 | 18.2 |
© 2014 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Parker, M.S.; Sah, R.; Balasubramaniam, A.; Park, E.A.; Sallee, F.R.; Parker, S.L. Dimers of G-Protein Coupled Receptors as Versatile Storage and Response Units. Int. J. Mol. Sci. 2014, 15, 4856-4877. https://doi.org/10.3390/ijms15034856
Parker MS, Sah R, Balasubramaniam A, Park EA, Sallee FR, Parker SL. Dimers of G-Protein Coupled Receptors as Versatile Storage and Response Units. International Journal of Molecular Sciences. 2014; 15(3):4856-4877. https://doi.org/10.3390/ijms15034856
Chicago/Turabian StyleParker, Michael S., Renu Sah, Ambikaipakan Balasubramaniam, Edwards A. Park, Floyd R. Sallee, and Steven L. Parker. 2014. "Dimers of G-Protein Coupled Receptors as Versatile Storage and Response Units" International Journal of Molecular Sciences 15, no. 3: 4856-4877. https://doi.org/10.3390/ijms15034856
APA StyleParker, M. S., Sah, R., Balasubramaniam, A., Park, E. A., Sallee, F. R., & Parker, S. L. (2014). Dimers of G-Protein Coupled Receptors as Versatile Storage and Response Units. International Journal of Molecular Sciences, 15(3), 4856-4877. https://doi.org/10.3390/ijms15034856