Investigation into Variation of Endogenous Metabolites in Bone Marrow Cells and Plasma in C3H/He Mice Exposed to Benzene


Abstract
:1. Introduction
2. Results and Discussion
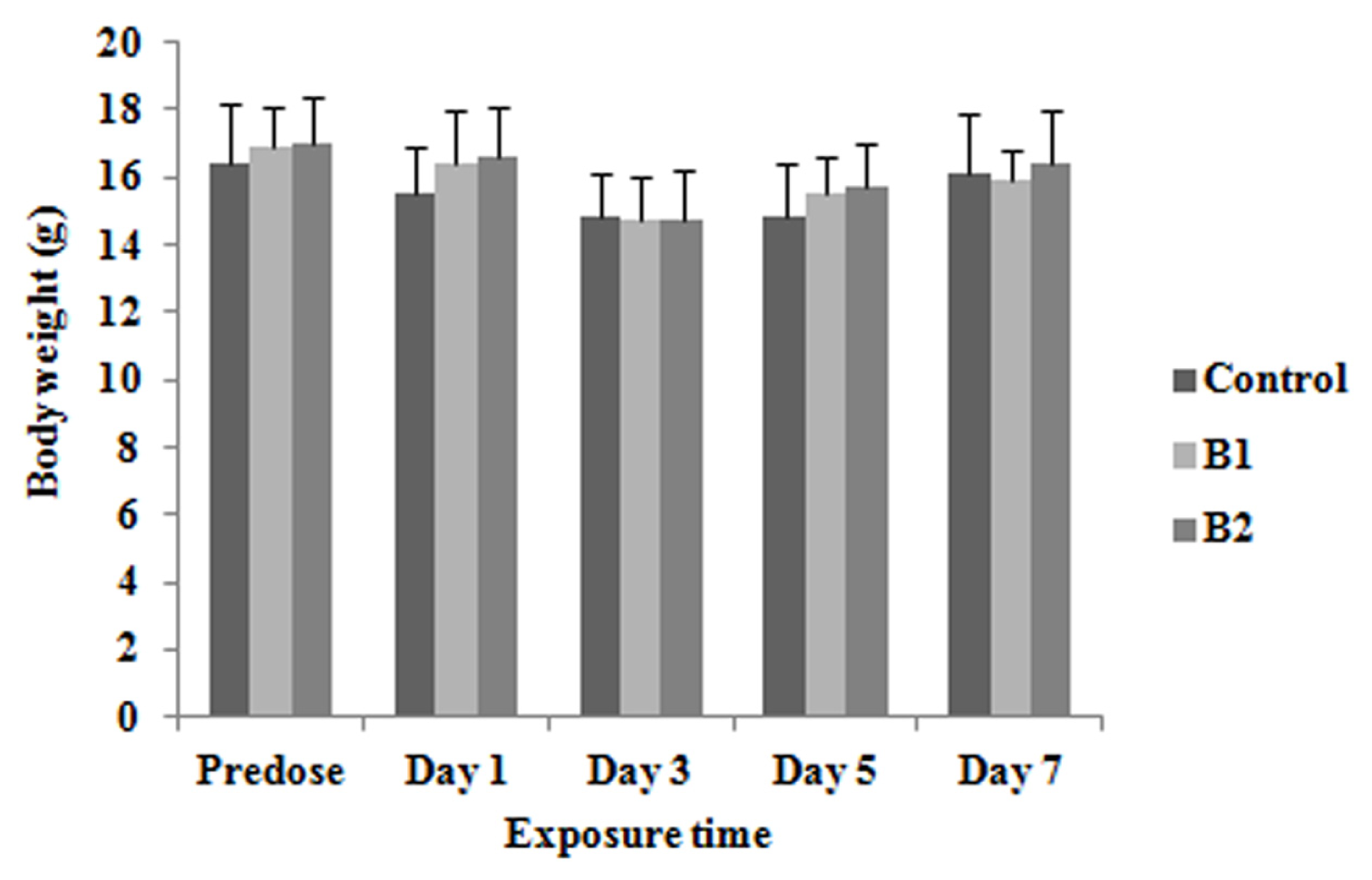
2.1. Body Weights and Relative Organ Weights
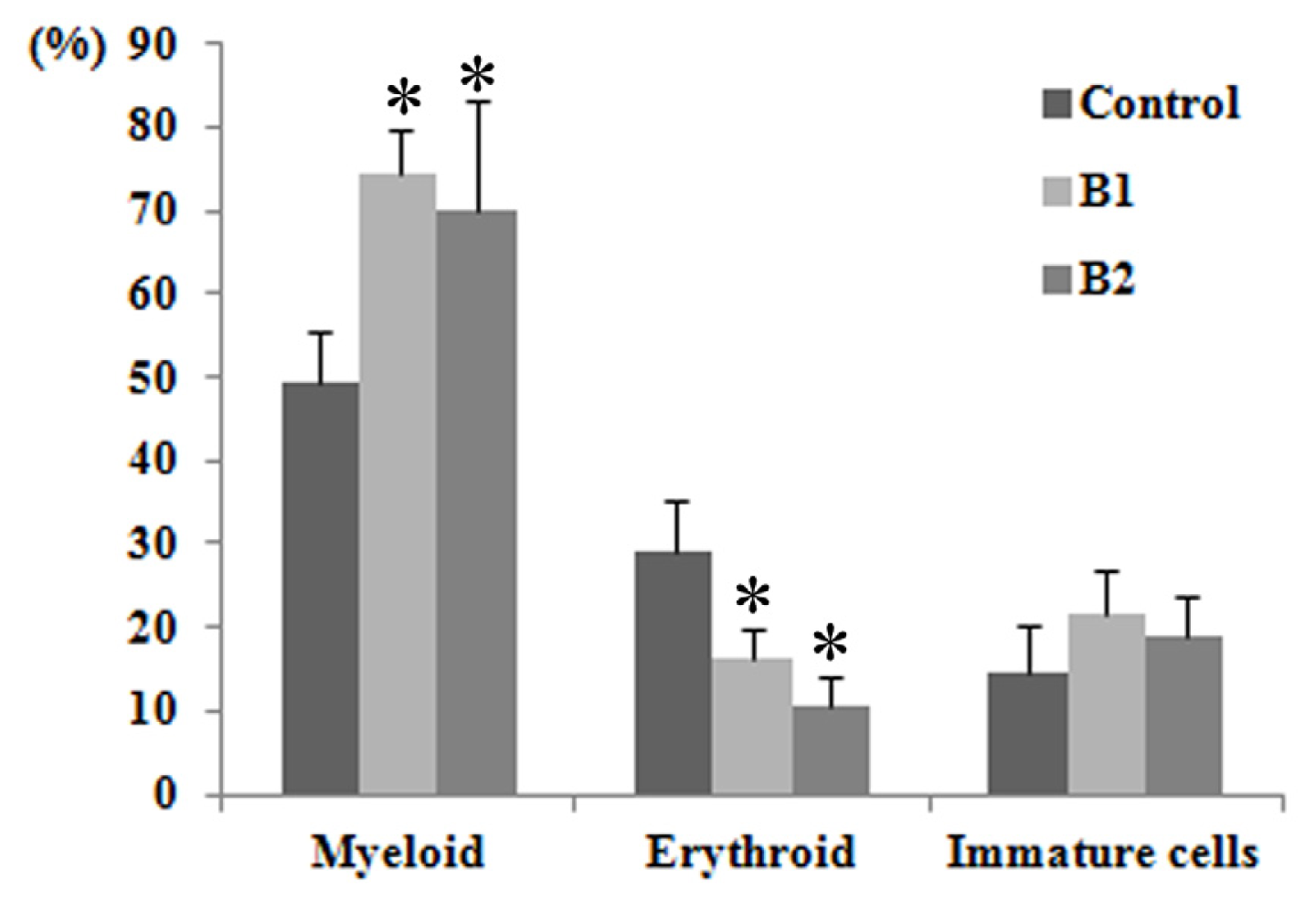
2.2. Blood Parameters and Bone Marrow Smear
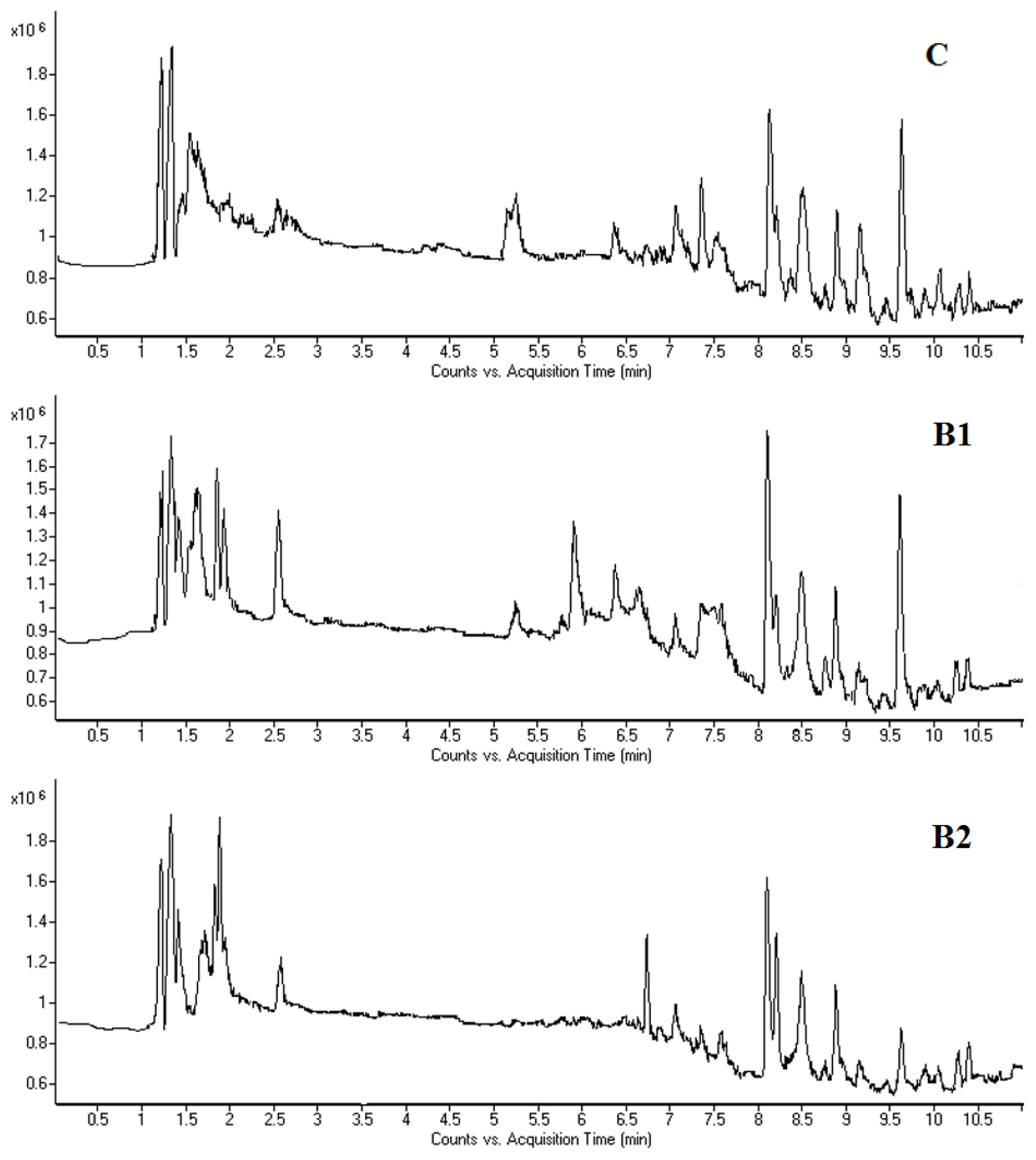
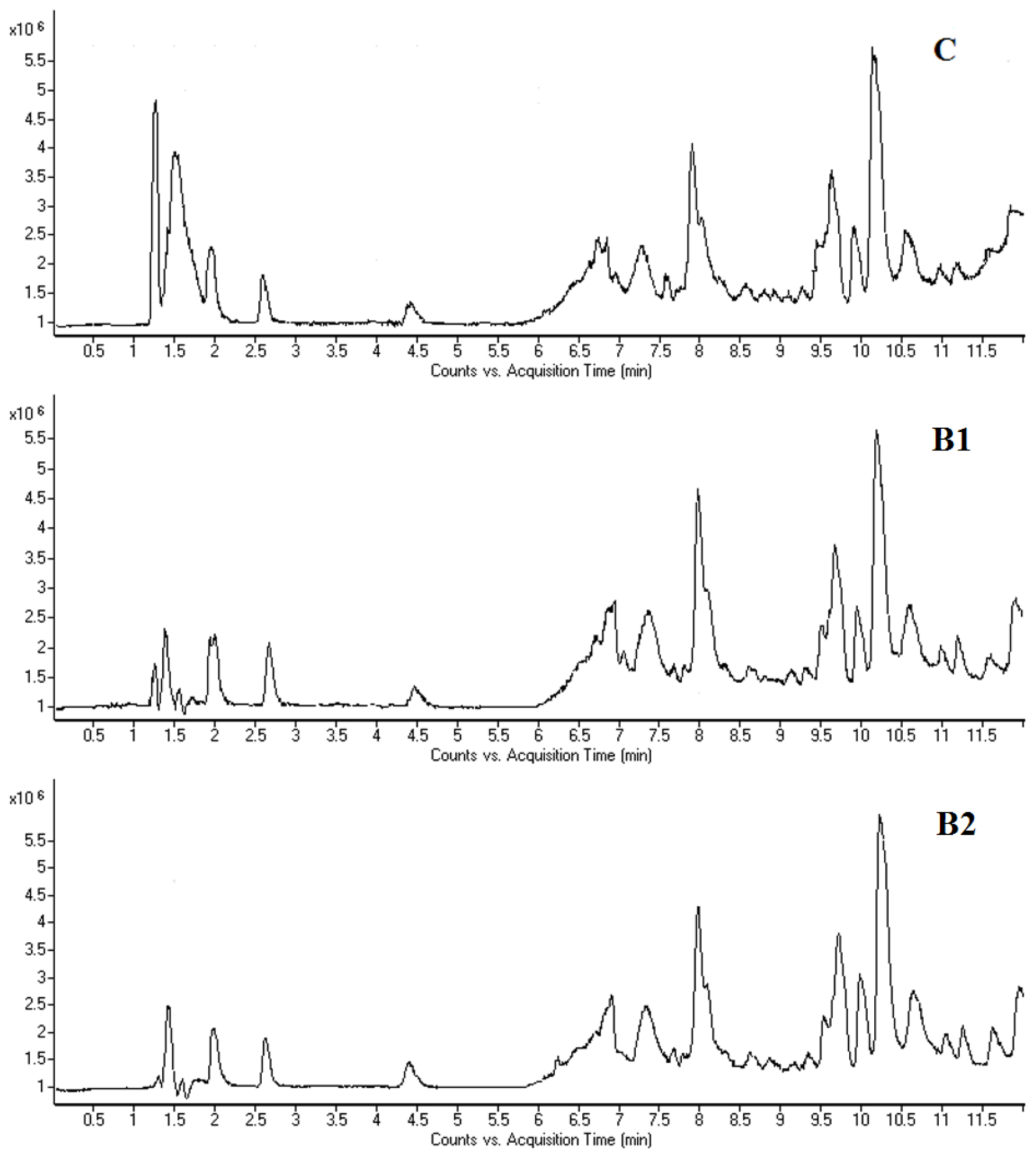
2.3. LC-MS Fingerprinting of Mouse Bone Marrow Cells and Plasma
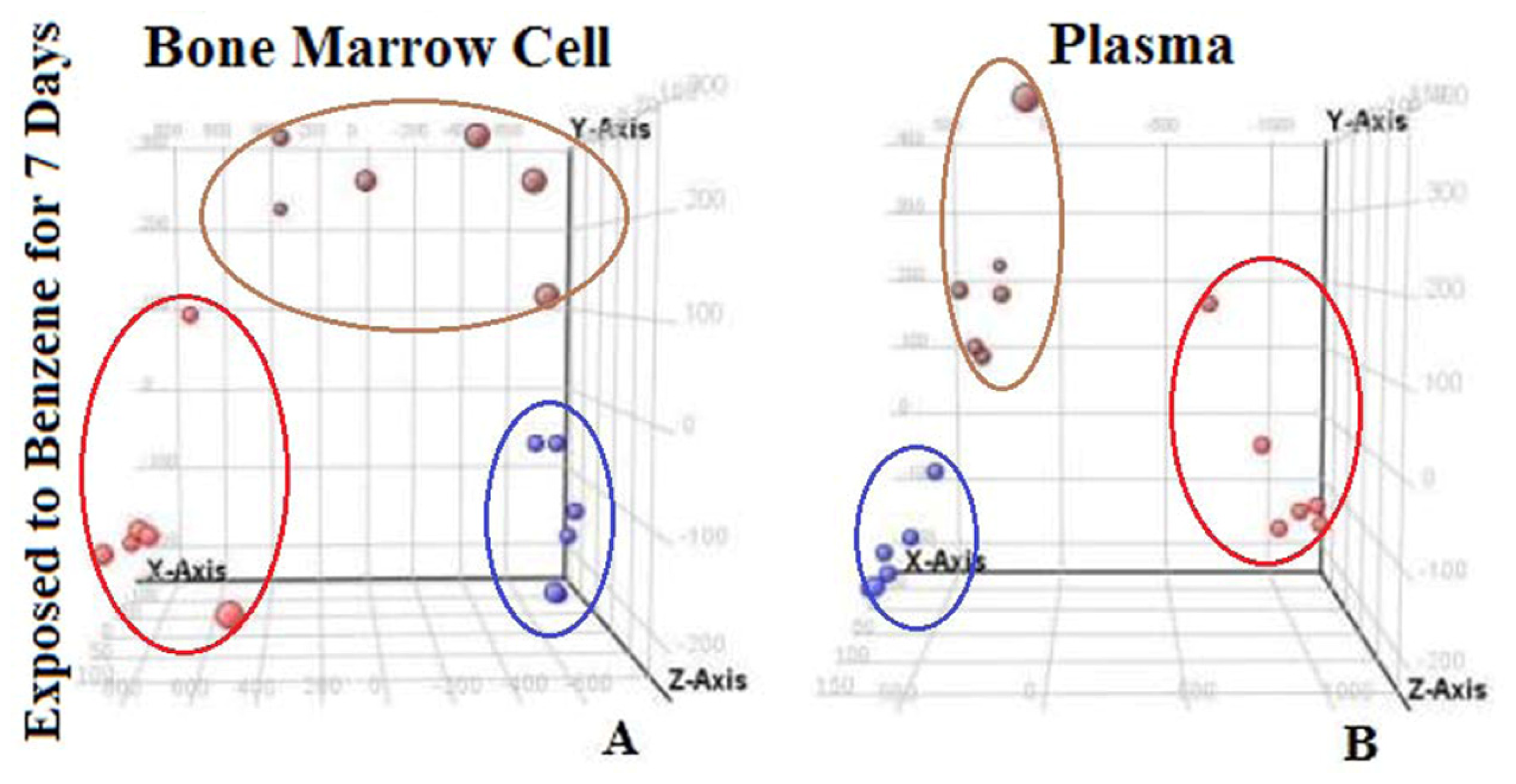
2.4. Principal Component Analysis and Discovery of Changed Endogenous Metabolites
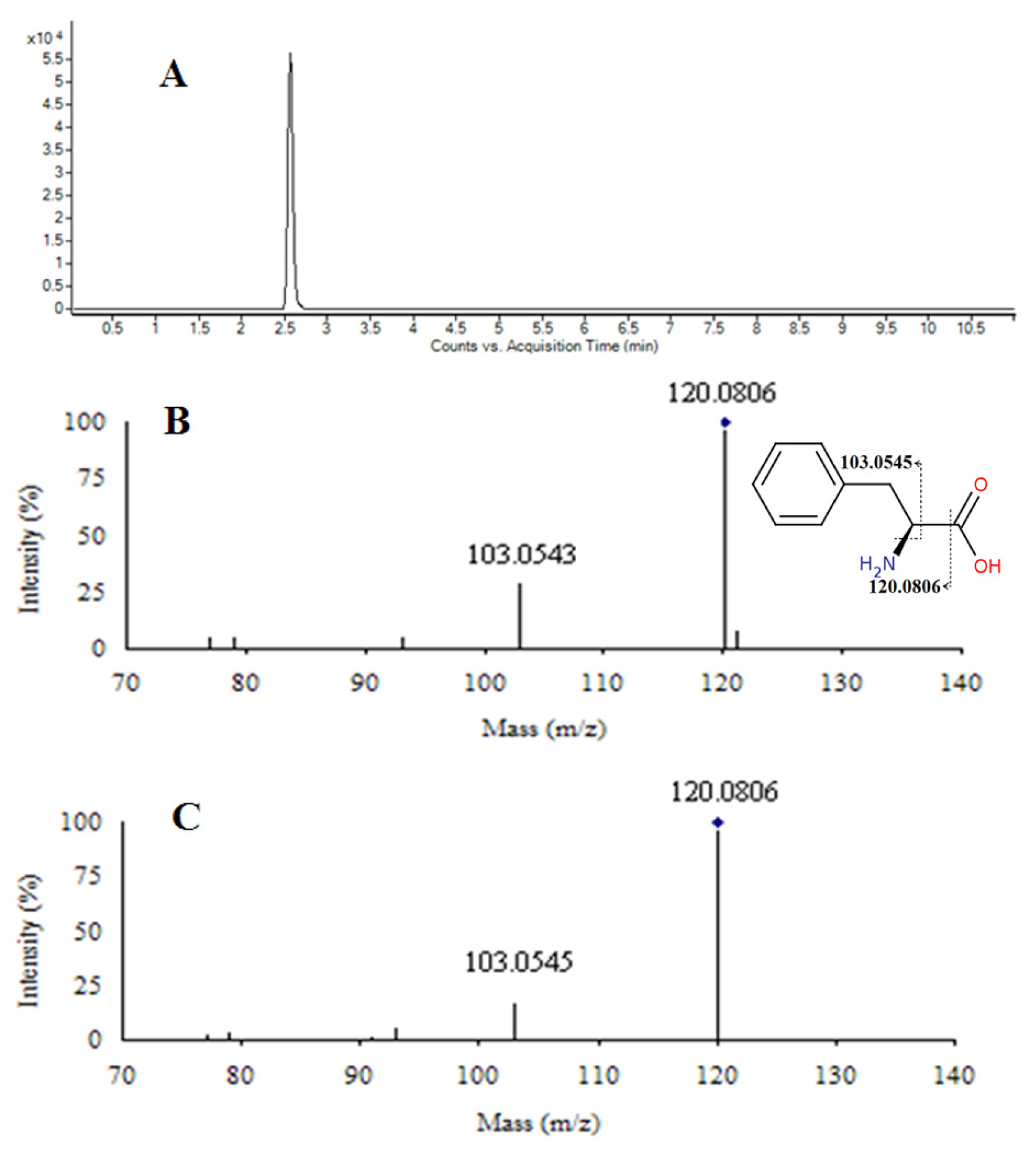
2.5. Identification of Changed Endogenous Metabolites
2.6. Biological Significance of Endogenous Metabolites Alternations in Bone Marrow Cells and Plasma
2.6.1. Significance of Changed Endogenous Metabolites in Bone Marrow Cells Induced by Benzene
2.6.2. Significance of Changed Endogenous Metabolites in Plasma Induced by Benzene
3. Experimental Section
3.1. Chemicals and Reagents
3.2. Ethics Statement
3.3. Animals and Treatments
3.4. Collection of Plasma and Bone Marrow Cell Samples
3.5. Sample Preparation and HPLC/MS Analysis
3.6. Data Processing
3.7. Statistics
4. Conclusions
Acknowledgments
Conflicts of Interest
References
- Shen, M.; Lan, Q.; Zhang, L.; Chanock, S.; Li, G.; Vermeulen, R.; Rappaport, S.M.; Guo, W.; Hayes, R.B.; Linet, M.; et al. Polymorphisms in genes involved in DNA double-strand break repair pathway and susceptibility to benzene-induced hematotoxicity. Carcinogenesis 2006, 27, 2083–2089. [Google Scholar]
- Glass, D.C.; Gray, C.N.; Jolley, D.J.; Gibbons, C.; Sim, M.R.; Fritschi, L.; Adams, G.G.; Bisby, J.A.; Manuell, R. Leukemia risk associated with low-level benzene exposure. Epidemiology 2003, 14, 569–577. [Google Scholar]
- Hayes, R.B.; Songnian, Y.; Dosemeci, M.; Linet, M. Benzene and lymphohematopoietic malignancies in humans. Am. J. Ind. Med 2001, 40, 117–126. [Google Scholar]
- Smith, M.T.; Jones, R.M.; Smith, A.H. Benzene exposure and risk of non-Hodgkin lymphoma. Cancer Epidemiol. Biomark. Prev. 2007, 16, 385–391. [Google Scholar]
- International Agency for Research on Cancer (IARC). Benzene IARC monographs on the evaluation of the carcinogenic risk of chemicals to humans: Some industrial chemicals and dyestuffs. Int. Agency Res. Cancer 1982, 29, 93–148.
- IARC (International Agency for Research on Cancer). Monographs on the Evaluation of Carcinogenic Risks to Humans Overall Evaluations of Carcinogenicity. Available online: http://monographs.iarc.fr/ENG/Monographs/vol71/mono71.pdf (accessed on 3 March 2014).
- Qu, Q.; Shore, R.; Li, G.; Jin, X.; Chen, L.C.; Cohen, B.; Melikian, A.A.; Eastmond, D.; Rappaport, S.M.; Yin, S.; et al. Hematological changes among Chinese workers with a broad range of benzene exposure. Am. J. Ind. Med 2002, 42, 275–285. [Google Scholar]
- Rothman, N.; Li, G.L.; Dosemeci, M.; Bechtold, W.E.; Marti, G.E.; Wang, Y.Z.; Linet, M.; Xi, L.Q.; Lu, W.; Smith, M.T.; et al. Hematotoxicity among Chinese workers heavily exposed to benzene. Am. J. Ind. Med 1996, 29, 236–246. [Google Scholar]
- Snyder, R. Overview of the toxicology of benzene. J. Toxicol. Environ. Health 2000, 61, 339–346. [Google Scholar]
- Wilbur, S.; Wohlers, D.; Paikoff, S.; Keith, L.S.; Faroon, O. ATSDR evaluation of potential for human exposure to benzene. Toxicol. Ind. Health 2008, 24, 399–442. [Google Scholar]
- Subrahmanyam, V.V.; Doane-Setzer, P.; Steinmetz, K.L.; Ross, D.; Smith, M.T. Phenol-induced stimulation of hydroquinone bioactivation in mouse bone marrow in vivo: Possible implications in benzene myelotoxicity. Toxicology 1990, 62, 107–116. [Google Scholar]
- Subrahmanyam, V.V.; Kolachana, P.; Smith, M.T. Hydroxylation of phenol to hydroquinone catalyzed by a human myeloperoxidase-superoxide complex: Possible implications in benzene-induced myelotoxicity. Free Radic. Res. Commun 1991, 15, 285–296. [Google Scholar]
- Zhang, L.; Eastmond, D.A.; Smith, M.T. The nature of chromosomal aberrations detected in humans exposed to benzene. Crit. Rev. Toxicol. 2002, 32, 1–42. [Google Scholar]
- Longacre, S.L.; Kocsis, J.J.; Snyder, R. Influence of strain diffrences in mice on the metabolism and toxicity of benzene. Toxicol. Appl. Pharmacol. 1981, 60, 398–409. [Google Scholar]
- Billet, S.; Paget, V.; Garcon, G.; Heutte, N.; Andre, V.; Shirali, P.; Sichel, F. Benzene-induced mutational pattern in the tumour suppressor gene Tp53 analysed by use of a functional assay the functional analysis of separated alleles in yeast in human lung cells. Arch. Toxicol. 2010, 84, 99–107. [Google Scholar]
- McHale, C.M.; Zhang, L.; Lan, Q.; Vermeulen, R.; Li, G.; Hubbard, A.E.; Porter, K.E.; Thomas, R.; Portier, C.J.; Shen, M.; et al. Global gene expression profling of a population exposed to a range of benzene levels. Environ. Health Perspect 2011, 119, 628–634. [Google Scholar]
- Kolachana, P.; Subrahmanyam, V.V.; Meyer, K.B.; Zhang, L.; Smith, M.T. Benzene and its phenolic metabolites produce oxidative DNA damage in HL60 cells in vitro and in the bone marrow in vivo. Cancer Res. 1993, 53, 1023–1026. [Google Scholar]
- Zhang, L.; McHale, C.M.; Rothman, N.; Li, G.; Ji, Z.; Vermeulen, R.; Hubbard, A.E.; Ren, X.; Shen, M.; Rappaport, S.M.; et al. Systems biology of human benzene exposure. Chem. Biol. Interact 2010, 184, 86–93. [Google Scholar]
- Snyder, R. Benzene and leukemia. Crit. Rev. Toxicol 2002, 32, 155–210. [Google Scholar]
- Wang, I.M.; Stone, D.J.; Nickle, D.; Loboda, A.; Puig, O.; Roberts, C. Systems biology approach for new target and biomarker identification. Curr. Top. Microbiol. Immunol 2013, 363, 169–199. [Google Scholar]
- Gavaghan, C.L.; Wilson, I.D.; Nicholson, J.K. Physiological variation in metabolic phenotyping and functional genomic studies: Use of orthogonal signal correction and PLS-DA. FEBS Lett 2002, 530, 191–196. [Google Scholar]
- O’Connell, T.M.; Watkins, P.B. The application metabonomics to predict drug-induced liver injury. Clin. Pharmacol. Ther. 2010, 88, 394–399. [Google Scholar]
- Robertson, D.G. Metabonomics in toxicology: A review. Toxicol. Sci 2005, 85, 809–822. [Google Scholar]
- Cano, K.E.; Li, L.; Bhatia, R.; Bhatia, R.; Forman, S.J.; Chen, Y. NMR-based metabolomic analysis of the molecular pathogenesis of therapy-related myelodysplasia/acute myeloid leukemia. J. Proteome Res. 2011, 10, 2873–2881. [Google Scholar]
- Wang, Y.; Gao, D.; Chen, Z.; Li, S.; Gao, C.; Cao, D.; Liu, F.; Liu, H.; Jiang, Y. Acridone derivative 8a induces oxidative stress-mediated apoptosis in CCRF-CEM leukemia cells: Application of metabolomics in mechanistic studies of antitumor agents. PLoS One 2013, 8, e63572. [Google Scholar]
- Huang, Z.; Lin, L.; Cao, Y.; Chen, Y.; Yan, X.; Xing, J.; Hang, W. Bladder cancer determination via two metabolites: A biomarker pattern approach. Mol. Cell. Proteomics 2011, 10. [Google Scholar] [CrossRef]
- Theodoridis, G.A.; Gika, H.G.; Want, E.J.; Wilson, I.D. Liquid chromatography–mass spectrometry based global metabolite profling: A review. Anal. Chim. Acta 2012, 711, 7–16. [Google Scholar]
- Sun, R.; Zhang, J.; Xiong, M.; Chen, Y.; Yin, L.; Pu, Y. Metabonomics biomarkers for subacute toxicity screening for benzene exposure in mice. J. Toxicol. Environ. Health A 2012, 75, 1163–1173. [Google Scholar]
- Coen, M.; Holmes, E.; Lindon, J.C.; Nicholson, J.K. NMR-based metabolic profiling and metabonomic approaches to problems in molecular toxicology. Chem. Res. Toxicol. 2008, 21, 9–27. [Google Scholar]
- Wang, Y.; Zhang, L.; Chen, W.L.; Wang, J.H.; Li, N.; Li, J.M.; Mi, J.Q.; Zhang, W.N.; Li, Y.; Wu, S.F.; et al. Rapid diagnosis and prognosis of de novo acute myeloid leukemia by serum metabonomic analysis. J. Proteome Res 2013, 12, 4393–4401. [Google Scholar]
- Levi, M.M.; Vink, R.; de Jonge, E. Management of bleeding disorders by prohemostatic therapy. Int. J. Hematol. 2002, 76, 139–144. [Google Scholar]
- Yang, X.J. The diverse superfamily of lysine acetyltransferases and their roles in leukemia and other diseases. Nucleic Acids Res 2004, 32, 959–976. [Google Scholar]
- Koves, T.R.; Ussher, J.R.; Noland, R.C.; Slentz, D.; Mosedale, M.; Ilkayeva, O.; Bain, J.; Stevens, R.; Dyck, J.R.; Newgard, C.B.; et al. Mitochondrial overload and incomplete fatty acid oxidation contribute to skeletal muscle insulin resistance. Cell Metab. 2008, 7, 45–56. [Google Scholar]
- Ito, K.; Carracedo, A.; Weiss, D.; Arai, F.; Ala, U.; Avigan, D.E.; Schafer, Z.T.; Evans, R.M.; Suda, T.; Lee, C.H.; et al. A PML–PPAR-δ pathway for fatty acid oxidation regulates hematopoietic stem cell maintenance. Nat. Med 2012, 18, 1350–1358. [Google Scholar]
- Hough, L.B. Genomics meets histamine receptors: New subtypes new receptors. Mol. Pharmacol. 2001, 59, 415–419. [Google Scholar]
- Kletke, O.; Sergeeva, O.A.; Lorenz, P.; Oberland, S.; Meier, J.C.; Hatt, H.; Gisselmann, G. New insights in endogenous modulation of ligand-gated ion channels: Histamine is an inverse agonist at strychnine sensitive glycine receptors. Eur. J. Pharmacol 2013, 710, 59–66. [Google Scholar]
- Mellqvist, U.H.; Hansson, M.; Brune, M.; Dahlgren, C.; Hermodsson, S.; Hellstrand, K. Natural killer cell dysfunction and apoptosis induced by chronic myelogenous leukemia cells: Role of reactive oxygen species and regulation by histamine. Blood 2000, 96, 1961–1968. [Google Scholar]
- Brune, M.; Castaigne, S.; Catalano, J.; Gehlsen, K.; Ho, A.D.; Hofmann, W.K.; Hogge, D.E.; Nilsson, B.; Or, R.; Romero, A.I.; et al. Improved leukemia-free survival after postconsolidation immunotherapy with histamine dihydrochloride and interleukin-2 in acute myeloid leukemia: Results of a randomized phase 3 trial. Blood 2006, 108, 88–96. [Google Scholar]
- Smits, E.L.; Berneman, Z.N.; van Tendeloo, V.F. Immunotherapy of acute myeloid leukemia: Current approaches. Oncologist 2009, 14, 240–252. [Google Scholar]
- Aurelius, J.; Martner, A.; Brune, M.; Palmqvist, L.; Hansson, M.; Hellstrand, K.; Thoren, F.B. Remission maintenance in acute myeloid leukemia: Impact of functional histamine H2 receptors expressed by leukemic cells. Haematologica 2012, 97, 1904–1908. [Google Scholar]
- Subrahmanyam, V.V.; Ross, D.; Eastmond, D.A.; Smith, M.T. Potential role of free radicals in benzene-induced myelotoxicity and leukemia. Free Radic. Biol. Med 1991, 11, 495–515. [Google Scholar]
- Hiraku, Y.; Kawanishi, S. Oxidative DNA damage and apoptosis induced by benzene metabolites. Cancer Res. 1996, 56, 5172–5178. [Google Scholar]
- Mutomba, M.C.; Yuan, H.; Konyavko, M.; Adachi, S.; Yokoyama, C.B.; Esser, V.; McGarry, J.D.; Babior, B.M.; Gottlieb, R.A. Regulation of the activity of caspases by l-carnitine and palmitoylcarnitine. FEBS Lett. 2000, 478, 19–25. [Google Scholar]
- Ibuki, Y.; Goto, R. Dysregulation of apoptosis by benzene metabolites and their relationships with carcinogenesis. Biochim. Biophys. Acta 2004, 1690, 11–21. [Google Scholar]
- Allen, K.R.; Degg, T.J.; Anthoney, D.A.; Fitzroy-Smith, D. Monitoring the treatment of carcinoid disease using blood serotonin and plasma 5–hydroxyindoleacetic acid: Three case examples. Ann. Clin. Biochem. 2007, 44, 300–307. [Google Scholar]
- Kirillova, V.V.; Nigmatullina, R.R.; Dzhordzhikiya, R.K.; Kudrin, V.S.; Klodt, P.M. Increased concentrations of serotonin and 5-hydroxyindoleacetic acid in blood plasma from patients with pulmonary hypertension due to mitral valve disease. Bull. Exp. Biol. Med 2009, 147, 408–410. [Google Scholar]
- Peng, C.T.; Wu, K.H.; Lan, S.J.; Tsai, J.J.; Tsai, F.J.; Tsai, C.H. Amino acid concentrations in cerebrospinal fluid in children with acute lymphoblastic leukemia undergoing chemotherapy. Eur. J. Cancer 2005, 41, 1158–1163. [Google Scholar]
- Tunek, A.; Olofsson, T.; Berlin, M. Toxic effects of benzene and benzene metabolites on granulopoietic stem cells and bone marrow cellularity in mice. Toxicol. Appl. Pharmacol. 1981, 59, 149–156. [Google Scholar]
- Velasco Lezama, R.; Barrera Escorcia, E.; Muñoz Torres, A.; Tapia Aguilar, R.; González Ramírez, C.; García Lorenzana, M.; Ortiz Monroy, V.; Betancourt Rule, M. A model for the induction of aplastic anemia by subcutaneous administration of benzene in mice. Toxiology 2001, 162, 179–191. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Relative liver weight | Relative spleen weight | Relative lung weight | Relative kidney weight |
|---|---|---|---|---|
| Control | 6.01 ± 0.32 | 0.36 ± 0.05 | 0.69 ± 0.10 | 1.63 ± 0.14 |
| Benzene 1 | 6.38 ± 0.31 | 0.27 ± 0.09 * | 0.67 ± 0.08 | 1.65 ± 0.15 |
| Benzene 2 | 6.54 ± 0.48 | 0.25 ± 0.07 * | 0.56 ± 0.02 * | 1.69 ± 0.06 |
| Group | WBC (109/L) | RBC (1012/L) | Hgb (g/L) | Pit (109/L) |
|---|---|---|---|---|
| Control | 4.05 ± 0.65 | 7.98 ± 0.39 | 137.20 ± 5.76 | 364.25 ± 60.50 |
| Benzene 1 | 3.87 ± 1.06 | 7.30 ± 0.14 * | 126.67 ± 3.50 * | 322.00 ± 107.53 |
| Benzene 2 | 4.48 ± 0.96 | 7.32 ± 0.42 * | 127.60 ± 7.50 * | 282.2 ± 50.01 |
| Compartment | m/z | RT (Retention time) | Trend a | p-value | Metabolites | Related pathway |
|---|---|---|---|---|---|---|
| Bone Marrow Cell | 204.123 | 1.54 | ↓ | 9.30 × 10−4 | l-Acetylcarnitine | Oxidation of Fatty Acids |
| 165.0546 | 1.681 | ↑ | 5.14 × 10−3 | p-Coumaric acid | Unknown | |
| 182.081 | 1.668 | ↑ | 3.97 × 10−4 | l-tyrosine | Tyrosine/Phenylalanine and Tyrosine Metabolism, Catecholamine Biosynthesis | |
| 166.0863 | 2.548 | ↑ | 1.62 × 10−3 | l-Phenylalanine | Phenylalanine and Tyrosine Metabolism | |
| 147.1168 | 9.205 | ↓ | 1.65 × 10−4 | Lysine | Lysine Degradation, Biotin Metabolism, Carnitine Synthesis | |
| Plasma | 192.0644 | 1.2775 | ↓ | 1.92 × 10−6 | 5-Hydroxyindoleacetic acid | Tryptophan metabolism |
| 112.0869 | 1.3 | ↓ | 2.06 × 10−4 | Histamine | Histidine metabolism | |
| 156.0765 | 1.321 | ↑ | 1.08 × 10−5 | l-Histidine | Histidine Metabolism, Ammonia Recycling Transcription/Translation | |
| 126.1026 | 1.322 | ↓ | 1.32 × 10−4 | N-Methylhistamine | Histidine metabolism | |
| 204.123 | 1.58 | ↓ | 6.01 × 10−5 | l-Acetylcarnitine | Oxidation of Fatty Acids | |
| 130.0499 | 2.177 | ↑ | 2.88 × 10−6 | Pyrrolidonecarboxylic acid | Gamma-glutamyl cycle | |
| 400.3421 | 9.195 | ↓ | 3.11 × 10−7 | Palmitoylcarnitine | Fatty acid Metabolism | |
© 2014 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Sun, R.; Zhang, J.; Yin, L.; Pu, Y. Investigation into Variation of Endogenous Metabolites in Bone Marrow Cells and Plasma in C3H/He Mice Exposed to Benzene. Int. J. Mol. Sci. 2014, 15, 4994-5010. https://doi.org/10.3390/ijms15034994
Sun R, Zhang J, Yin L, Pu Y. Investigation into Variation of Endogenous Metabolites in Bone Marrow Cells and Plasma in C3H/He Mice Exposed to Benzene. International Journal of Molecular Sciences. 2014; 15(3):4994-5010. https://doi.org/10.3390/ijms15034994
Chicago/Turabian StyleSun, Rongli, Juan Zhang, Lihong Yin, and Yuepu Pu. 2014. "Investigation into Variation of Endogenous Metabolites in Bone Marrow Cells and Plasma in C3H/He Mice Exposed to Benzene" International Journal of Molecular Sciences 15, no. 3: 4994-5010. https://doi.org/10.3390/ijms15034994
APA StyleSun, R., Zhang, J., Yin, L., & Pu, Y. (2014). Investigation into Variation of Endogenous Metabolites in Bone Marrow Cells and Plasma in C3H/He Mice Exposed to Benzene. International Journal of Molecular Sciences, 15(3), 4994-5010. https://doi.org/10.3390/ijms15034994