Albumin Suppresses Human Hepatocellular Carcinoma Proliferation and the Cell Cycle

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
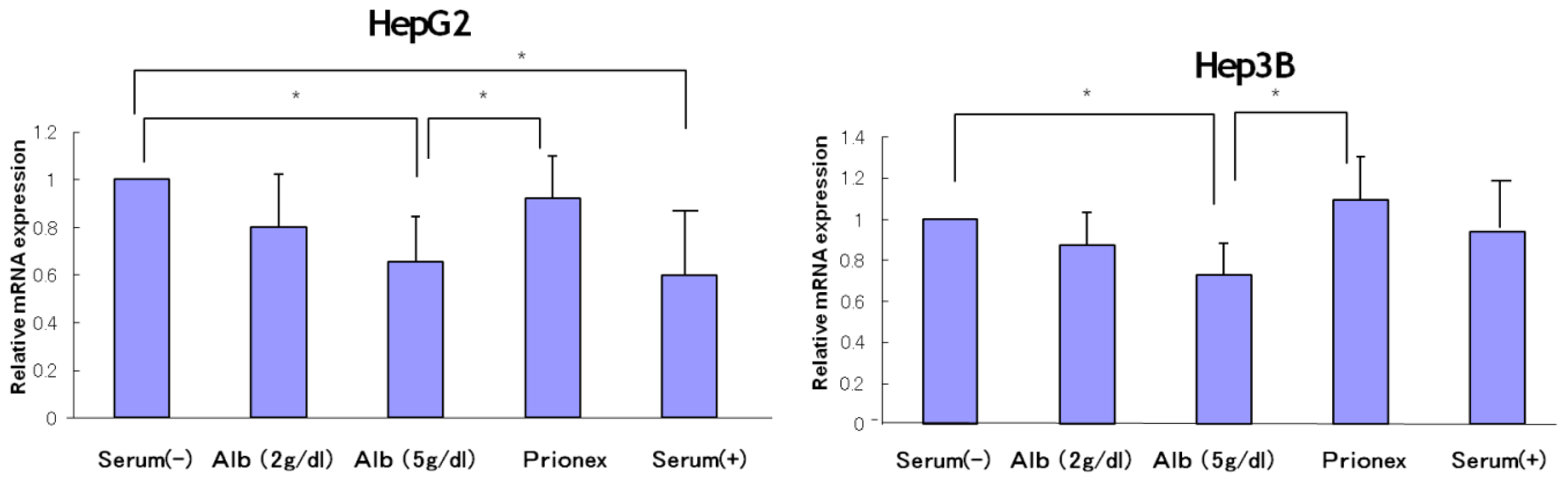
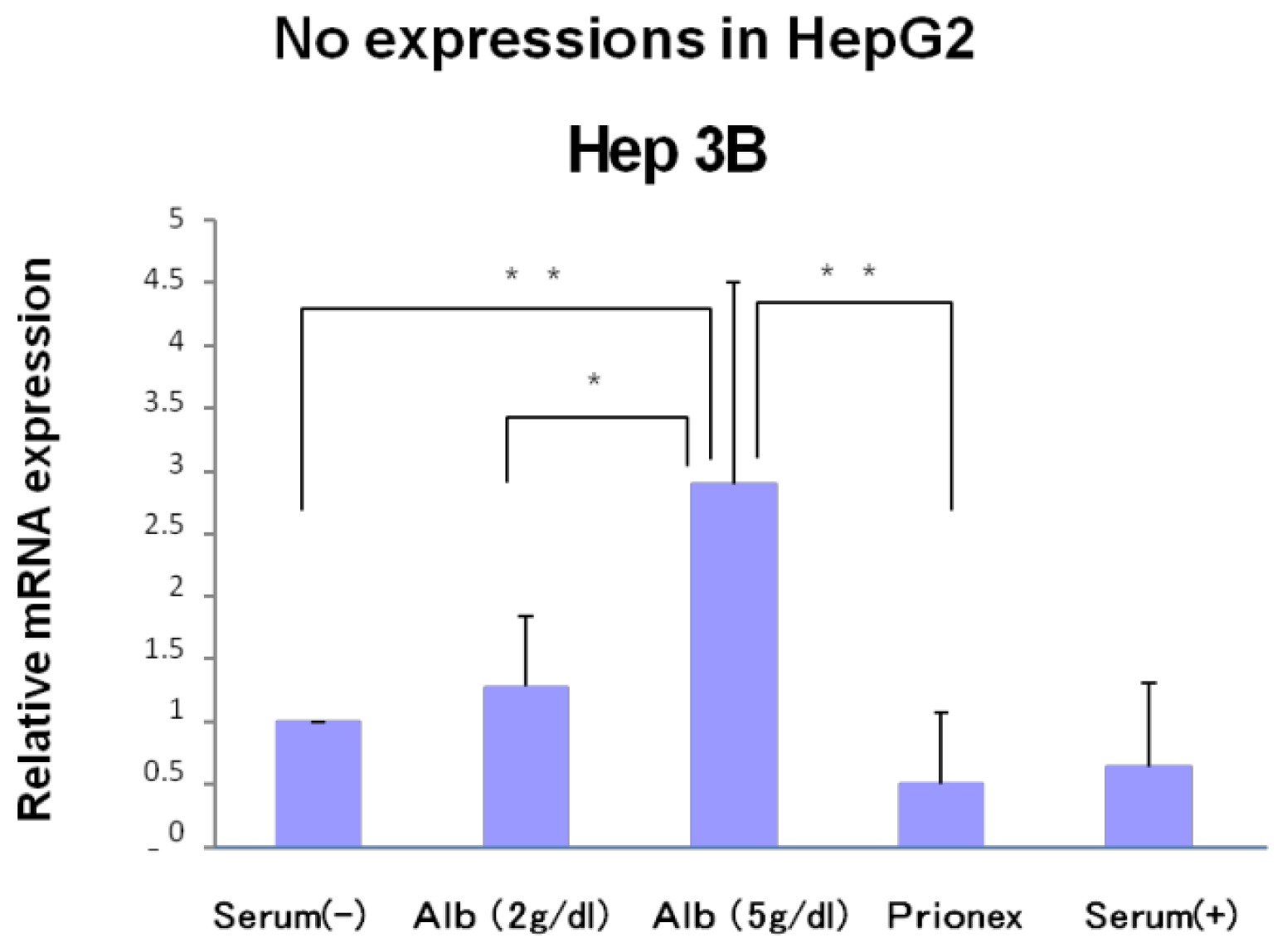
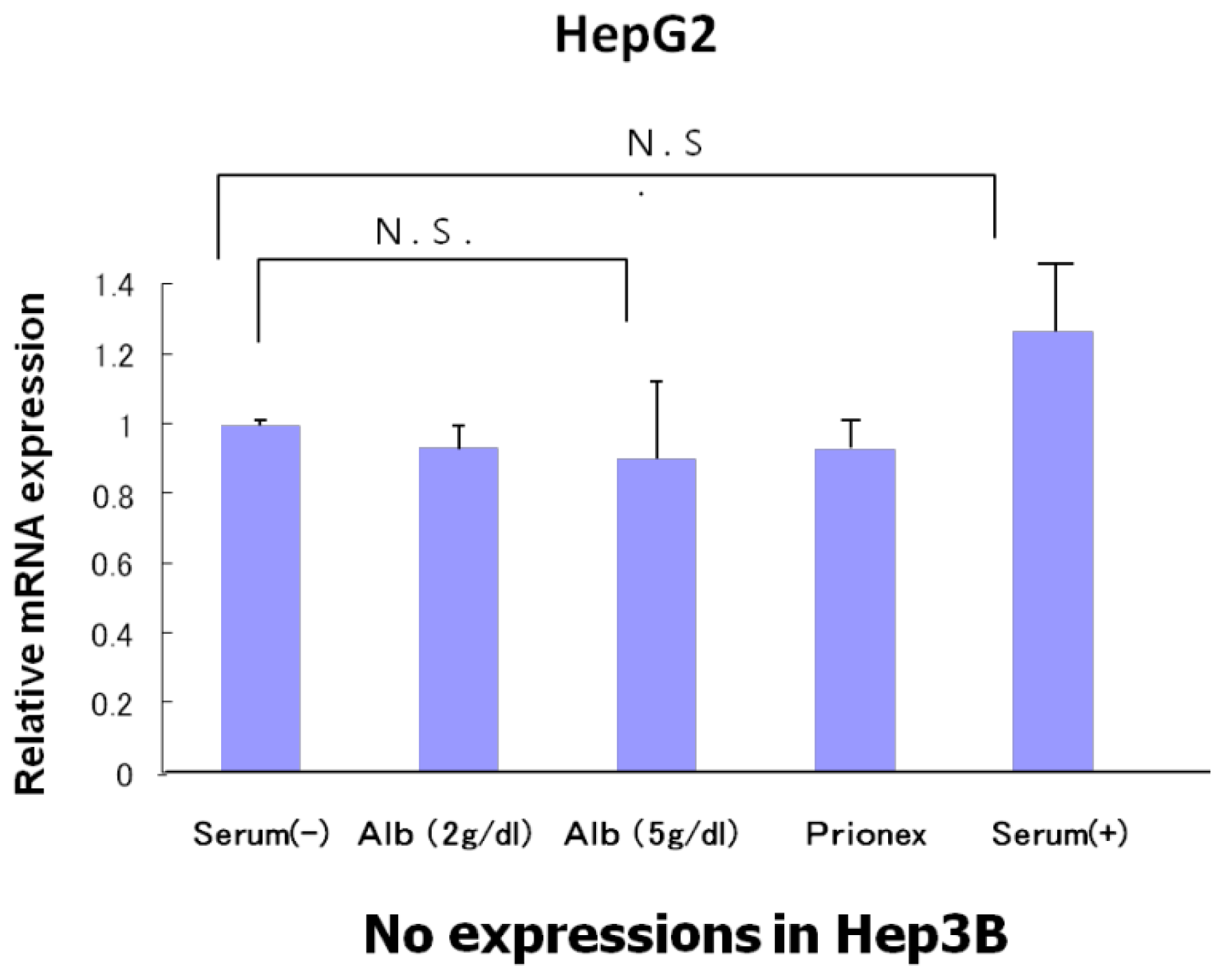
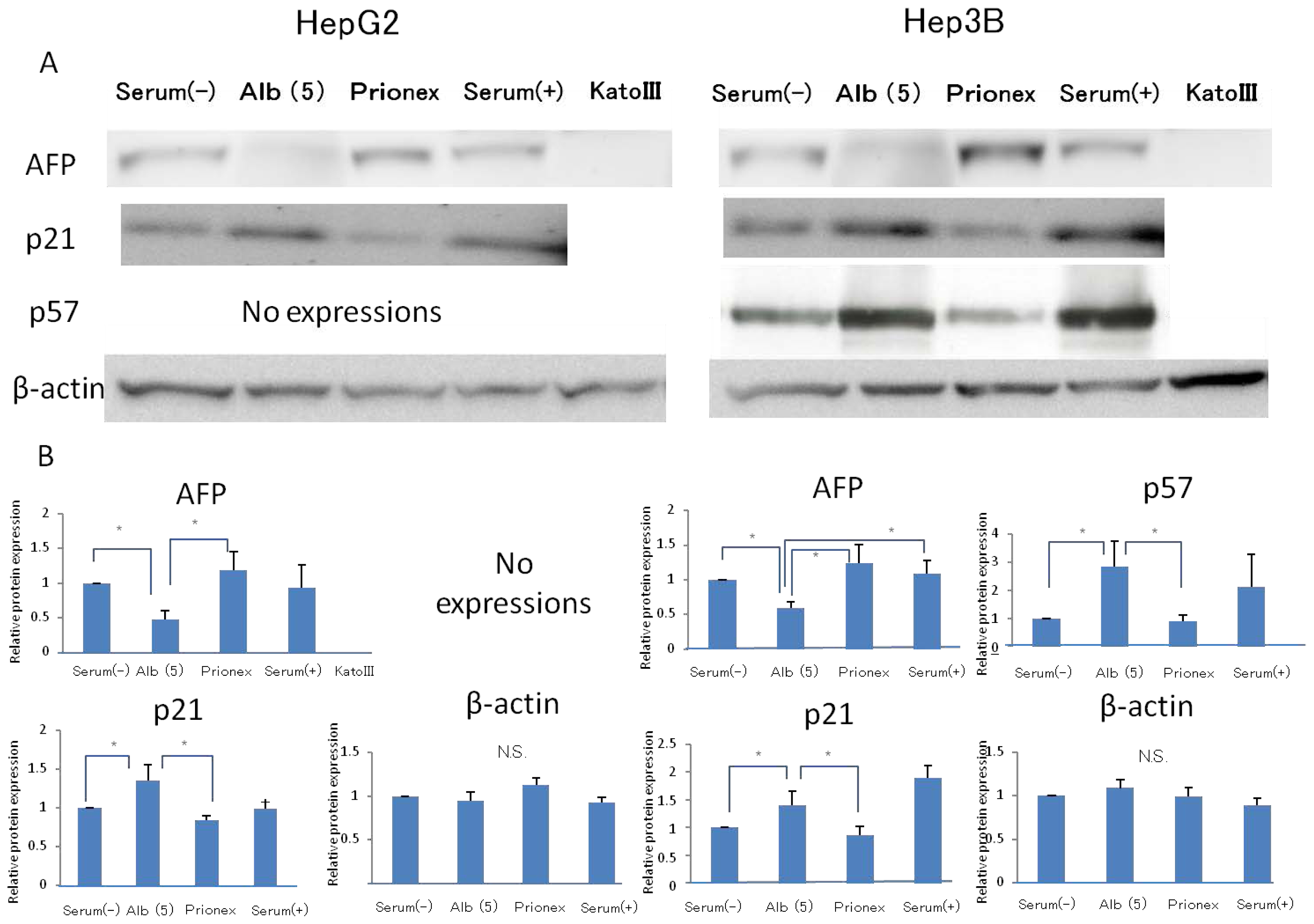
2.1. Effect of Albumin Concentrations on AFP, p21, and p57 mRNA Expression and in HepG2 and Hep3B (Figures 1–4)
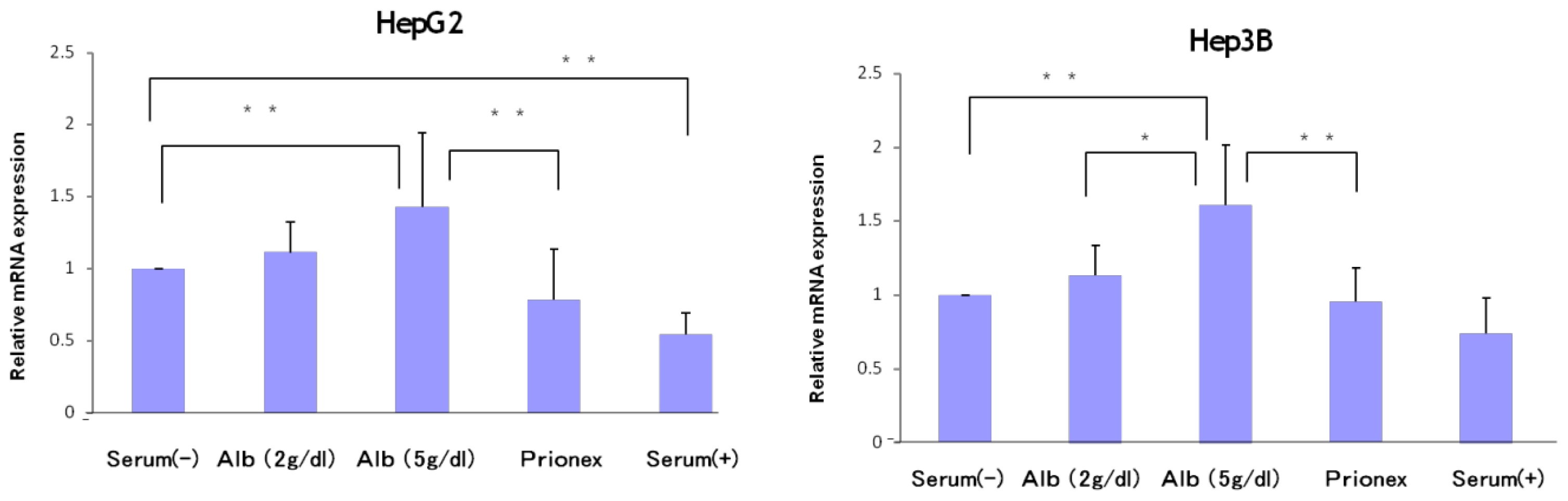
2.2. Effect of 5 g/dL Albumin Concentrations on AFP, p21, and p57 mRNA Expressions in HepG2 and Hep3B (Figure 5)
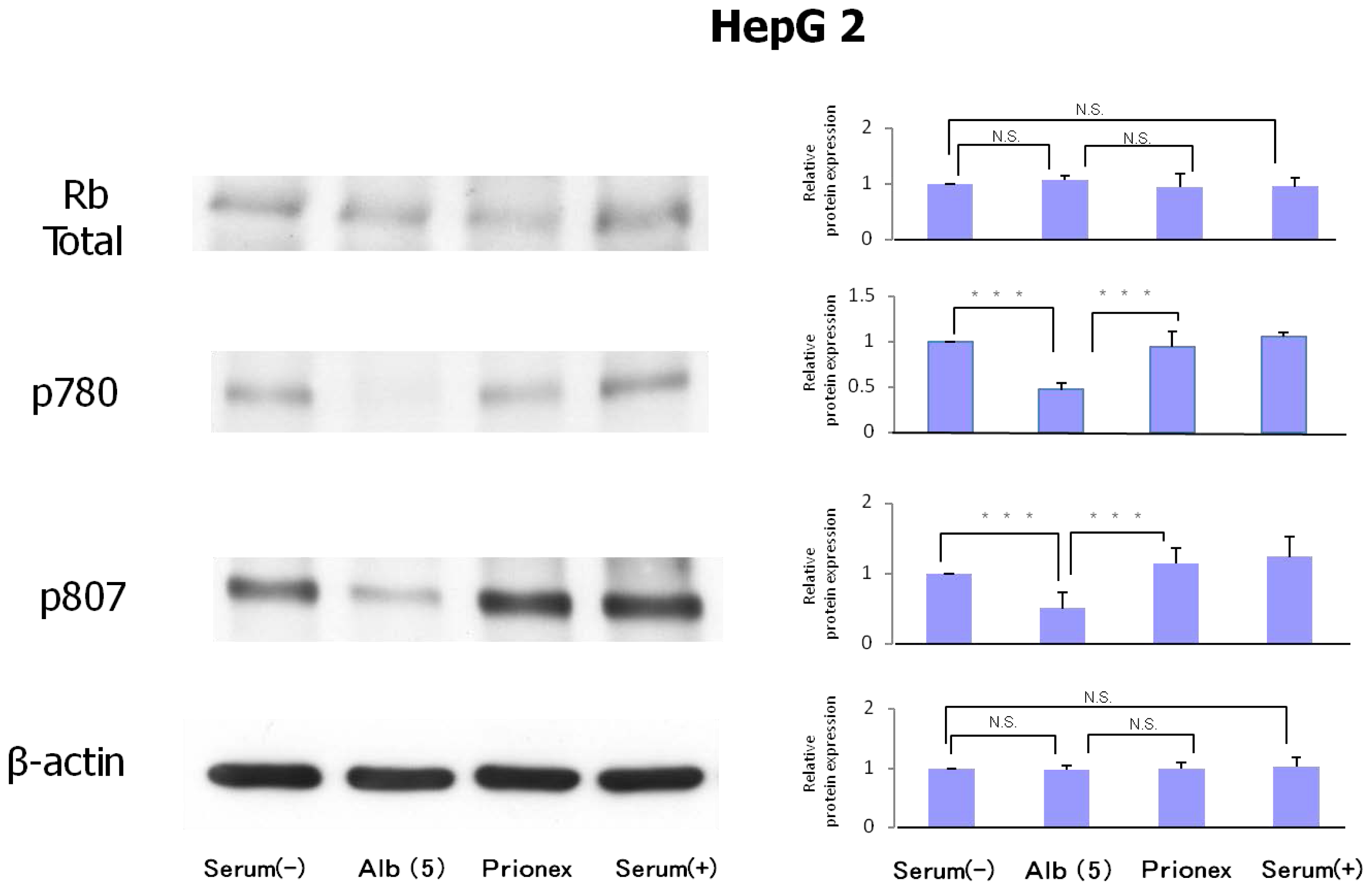
2.3. Protein Expression of Rb and Phosphorylation of p780 and p807 (Figure 6)
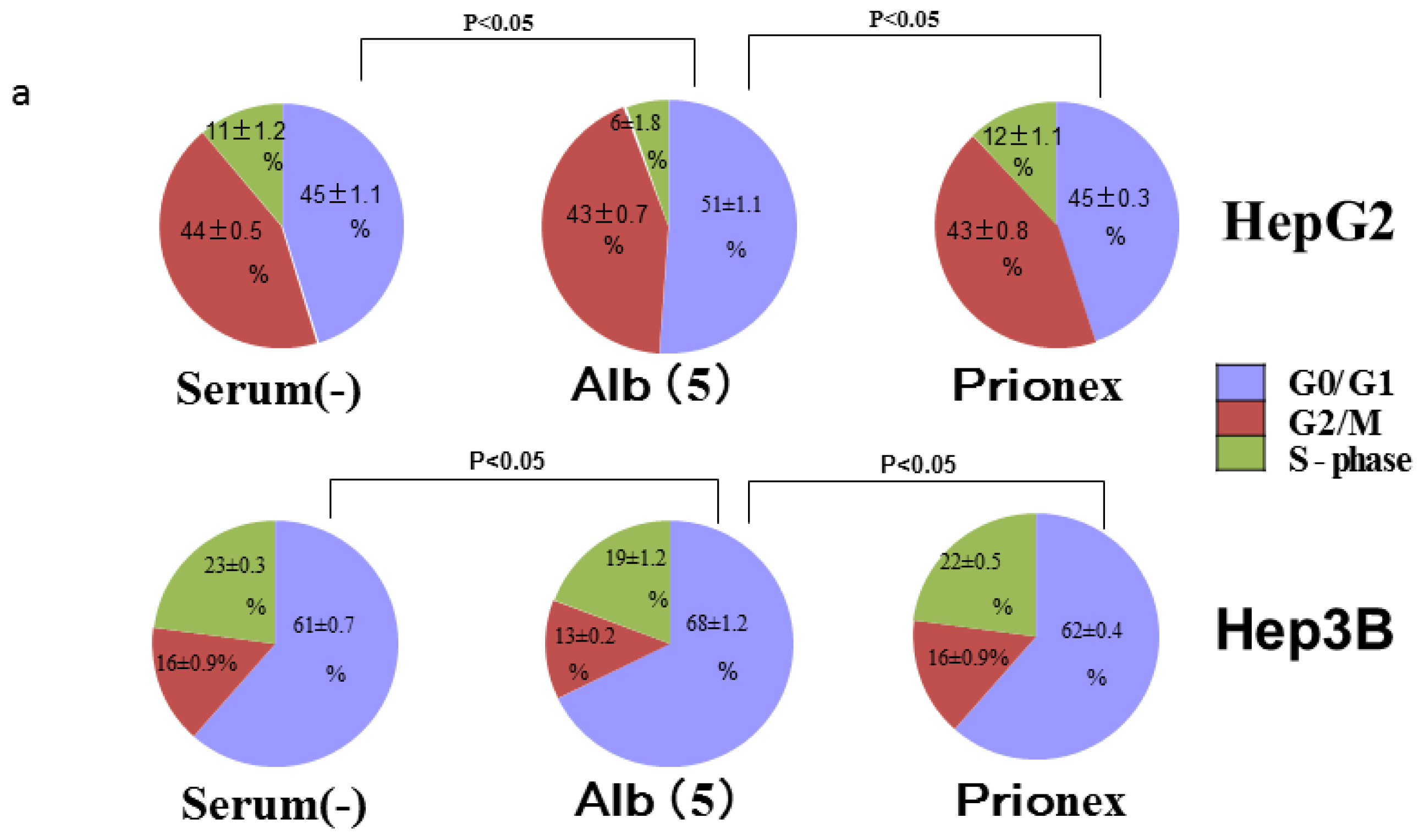
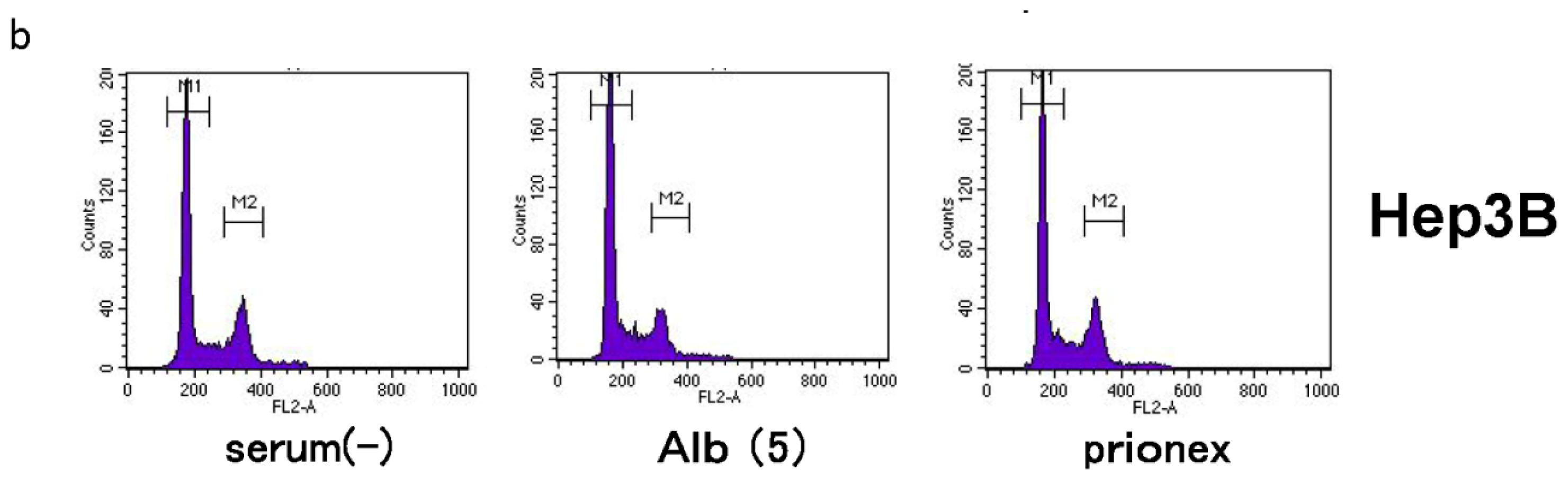
2.4. Albumin Concentration Effect on the Cell Cycle (Figure 7)
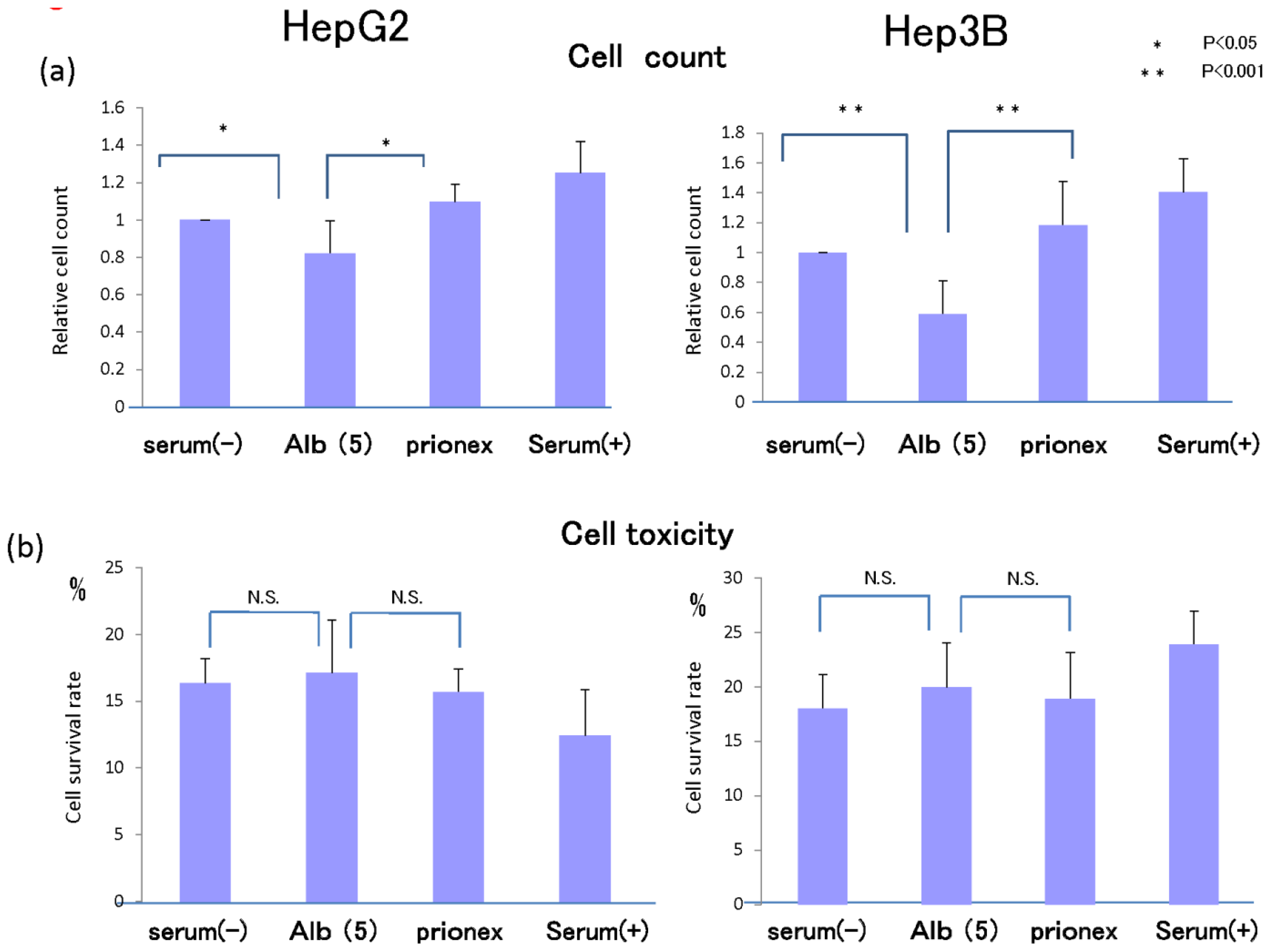
2.5. Cell Proliferation in Vitro and Cell Toxicity Measurement of Albumin (Figure 8)
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Gel Electrophoresis and Western Blotting
4.3. Real-Time Polymerase Chain Reaction (PCR) for Quantitative Assessment of mRNA Expression
4.4. Cell Count and Cell Toxicity Measurement
4.5. Flow Cytometric Analysis
5. Conclusions
Conflicts of Interest
References
- El-Serag, H.B.; Rudolph, K.L. Hepatocellular carcinoma: Epidemiology and molecular carcinogenesis. Gastroenterology 2007, 132, 2557–2576. [Google Scholar]
- Bruix, J.; Sherman, M. Management of hepatocellular carcinoma: An update. Hepatology 2011, 53, 1020–1022. [Google Scholar]
- Mazzaferro, V.; Romito, R.; Schiavo, M.; Mariani, L.; Camerini, T.; Bhoori, S.; Capussotti, L.; Calise, F.; Pellicci, R.; Belli, G.; et al. Prevention of hepatocellular carcinoma recurrence with alpha-interferon after liver resection in HCV cirrhosis. Hepatology 2006, 44, 1543–1554. [Google Scholar]
- Muto, Y.; Sato, S.; Watanabe, A.; Moriwaki, H.; Suzuki, K.; Kato, A.; Kato, M.; Nakamura, T.; Higuchi, K.; Nishiguchi, S.; et al. Effects of oral branched-chain amino acid granules on event-free survival in patients with liver cirrhosis. Clin. Gastroenterol. Hepatol. 2005, 3, 705–713. [Google Scholar]
- Nojiri, S.; Kusakabe, A.; Shinkai, N.; Matsuura, K.; Iio, E.; Miyaki, T.; Joh, T. Factors influencing distant recurrence of hepatocellular carcinoma following combined radiofrequency ablation and transarterial chemoembolization therapy in patients with hepatitis C. Cancer Manag. Res. 2011, 3, 267–272. [Google Scholar]
- Ikeda, K.; Arase, Y.; Saitoh, S.; Kobayashi, M.; Suzuki, Y.; Suzuki, F.; Tsubota, A.; Chayama, K.; Murashima, N.; Kumada, H. Interferon beta prevents recurrence of hepatocellular carcinoma after complete resection or ablation of the primary tumor—A prospective randomized study of hepatitis C virus-related liver cancer. Hepatology 2000, 32, 228–232. [Google Scholar]
- Muto, Y.; Moriwaki, H.; Ninomiya, M.; Adachi, S.; Saito, A.; Takasaki, K.T.; Tanaka, T.; Tsurumi, K.; Okuno, M.; Tomita, E.; et al. Prevention of second primary tumors by an acyclic retinoid polyprenoic acid in patients with hepatocellular carcinoma Hepatoma Prevention Study Group. N. Engl. J. Med. 1996, 334, 1561–1567. [Google Scholar]
- Kubo, S.; Yamamoto, T.; Ikebe, T.; Shuto, T.; Hirohashi, K.; Tanaka, H.; Tsukamoto, T.; Wakasa, K.; Kinoshita, H. Relationship between multicentric occurrence of hepatocellular carcinoma and histology of noncancerous hepatic tissue in patients with chronic hepatitis C. Jpn. J. Cancer Res. 1999, 90, 1076–1080. [Google Scholar]
- Marchesini, G.; Bianchi, G.; Merli, M.; Amodio, P.; Panella, C.; Loguercio, C.; Rossi Fanelli, F.; Abbiati, R. Nutritional supplementation with branched-chain amino acids in advanced cirrhosis: A double-blind randomized trial. Gastroenterology 2003, 124, 1792–1801. [Google Scholar]
- Ijichi, C.; Matsumura, T.; Tsuji, T.; Eto, Y. Branched-chain amino acids promote albumin synthesis in rat primary hepatocytes through the mTOR signal transduction system. Biochem. Biophys. Res. Commun. 2003, 303, 59–64. [Google Scholar]
- Kakazu, E.; Kanno, N.; Ueno, Y.; Shimosegawa, T. Extracellular branched-chain amino acids especially valine regulate maturation and function of monocyte-derived dendritic cells. J. Immunol. 2007, 179, 7137–7146. [Google Scholar]
- Nakamura, I.; Ochiai, K.; Imai, Y.; Moriyasu, F.; Imawari, M. Restoration of innate host defense responses by oral supplementation of branched-chain amino acids in decompensated cirrhotic patients. Hepatol. Res. 2007, 37, 1062–1067. [Google Scholar]
- Tsukishiro, T.; Shimizu, Y.; Higuchi, K.; Watanabe, A. Effect of branched-chain amino acids on the composition and cytolytic activity of liver-associated lymphocytes in rats. J. Gastroenterol. Hepatol. 2000, 15, 849–859. [Google Scholar]
- Saito, Y.; Saito, H.; Nakamura, M.; Wakabayashi, K.; Takagi, T.; Ebinuma, H.; Ishii, H. Effect of the molar ratio of branched-chain to aromatic amino acids on growth and albumin mRNA expression of human liver cancer cell lines in a serum-free medium. Nutr. Cancer 2001, 39, 126–131. [Google Scholar]
- Yoshiji, H.; Noguchi, R.; Kitade, M.; Kaji, K.; Ikenaka, Y.; Namisaki, T.; Yoshii, J.; Yanase, K.; Yamazaki, M.; Tsujimoto, T.; et al. Branched-chain amino acids suppress insulin-resistance-based hepatocarcinogenesis in obese diabetic rats. J. Gastroenterol. 2009, 44, 483–491. [Google Scholar]
- Iwasa, J.; Shimizu, M.; Shiraki, M.; Shirakami, Y.; Sakai, H.; Terakura, Y.; Takai, K.; Tsurumi, H.; Tanaka, T.; Moriwaki, H. Dietary supplementation with branched-chain amino acids suppresses diethylnitrosamine-induced liver tumorigenesis in obese and diabetic C57BL/KsJ-db/db mice. Cancer Sci. 2010, 101, 460–467. [Google Scholar]
- Roulin, D.; Cerantola, Y.; Dormond-Meuwly, A.; Demartines, N.; Dormond, O. Targeting mTORC2 inhibits colon cancer cell proliferation in vitro and tumor formation in vivo. Mol. Cancer 2010, 9, 57. [Google Scholar]
- Ninomiya, S.; Shimizu, M.; Imai, K.; Takai, K.; Shiraki, M.; Hara, T.; Tsurumi, H.; Ishizaki, S.; Moriwaki, H. Possible role of visfatin in hepatoma progression and the effects of branched-chain amino acids on visfatin-induced proliferation in human hepatoma cells. Cancer Prev. Res. (Phila.) 2011, 4, 2092–2100. [Google Scholar]
- Dowling, R.J.; Topisirovic, I.; Fonseca, B.D.; Sonenberg, N. Dissecting the role of mTOR: Lessons from mTOR inhibitors. Biochim. Biophys. Acta 2010, 1804, 433–439. [Google Scholar]
- Kim, R.D.; Stein, G.S.; Chari, R.S. Impact of cell swelling on proliferative signal transduction in the liver. J. Cell. Biochem. 2001, 83, 56–69. [Google Scholar]
- Wasik, M.; Gorski, A.; Nowaczyk, M. An optimal assay of human B cell proliferation in vitro. J. Immunol. Methods 1987, 99, 1–5. [Google Scholar]
- Bedotti, R.; Borghetti, A.F.; Favilla, R. Stimulatory effect of serum albumin on the proliferation of serum-free SV40-transformed Balb/c 3T3 cells. Biochim. Biophys. Acta 1990, 1053, 74–80. [Google Scholar]









© 2014 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Nojiri, S.; Joh, T. Albumin Suppresses Human Hepatocellular Carcinoma Proliferation and the Cell Cycle. Int. J. Mol. Sci. 2014, 15, 5163-5174. https://doi.org/10.3390/ijms15035163
Nojiri S, Joh T. Albumin Suppresses Human Hepatocellular Carcinoma Proliferation and the Cell Cycle. International Journal of Molecular Sciences. 2014; 15(3):5163-5174. https://doi.org/10.3390/ijms15035163
Chicago/Turabian StyleNojiri, Shunsuke, and Takashi Joh. 2014. "Albumin Suppresses Human Hepatocellular Carcinoma Proliferation and the Cell Cycle" International Journal of Molecular Sciences 15, no. 3: 5163-5174. https://doi.org/10.3390/ijms15035163
APA StyleNojiri, S., & Joh, T. (2014). Albumin Suppresses Human Hepatocellular Carcinoma Proliferation and the Cell Cycle. International Journal of Molecular Sciences, 15(3), 5163-5174. https://doi.org/10.3390/ijms15035163