An Efficient Agrobacterium-Mediated Transformation System for Poplar

Abstract
:
1. Introduction
2. Results and Discussion
2.1. Optimization of Poplar Regeneration



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Medium Number | 6-BA (mg/L) | TDZ (mg/L) | Mean Number of Main Shoots per Medium (Number of Main Shoots in Each Medium/Number of Explants in Each Medium) | Total Shoot Regeneration in Each Medium (%) | |
|---|---|---|---|---|---|
| 1 | 1.00 | 0.000 | 3.10 | 89.0 | |
| 2 | 1.00 | 0.002 | 3.50 | 93.0 | |
| 3 | 1.00 | 0.004 | 4.80 | 94.0 | |
| 4 | 1.00 | 0.008 | 3.30 | 89.0 | |
| 5 | 1.00 | 0.010 | 2.90 | 97.0 | |
| 6 | 0.80 | 0.000 | 38.8 | 85.0 | |
| 7 | 0.80 | 0.002 | 43.0 | 82.0 | |
| 8 | 0.80 | 0.004 | 45.8 | 78.0 | |
| 9 | 0.80 | 0.008 | 40.7 | 80.0 | |
| 10 | 0.80 | 0.010 | 30.8 | 79.0 | |
| 11 | 0.50 | 0.000 | 56.8 | 80.0 | |
| 12 | 0.50 | 0.002 | 65.2 | 75.0 | |
| 13 | 0.50 | 0.004 | 69.5 | 73.0 | |
| 14 | 0.50 | 0.008 | 60.5 | 76.0 | |
| 15 | 0.50 | 0.010 | 44.4 | 70.0 | |
| 16 | 0.20 | 0.000 | 8.90 | 37.0 | |
| 17 | 0.20 | 0.002 | 11.0 | 29.0 | |
| 18 | 0.20 | 0.004 | 14.0 | 35.0 | |
| 19 | 0.20 | 0.008 | 9.00 | 32.0 | |
| 20 | 0.20 | 0.010 | 6.13 | 26.0 | |
| 21 | 0.00 | 0.000 | 0.30 | 6.00 | |
| 22 | 0.00 | 0.002 | 0.50 | 9.00 | |
| 23 | 0.00 | 0.004 | 0.80 | 13.0 | |
| 24 | 0.00 | 0.008 | 0.50 | 10.0 | |
| 25 | 0.00 | 0.010 | 0.10 | 8.00 |

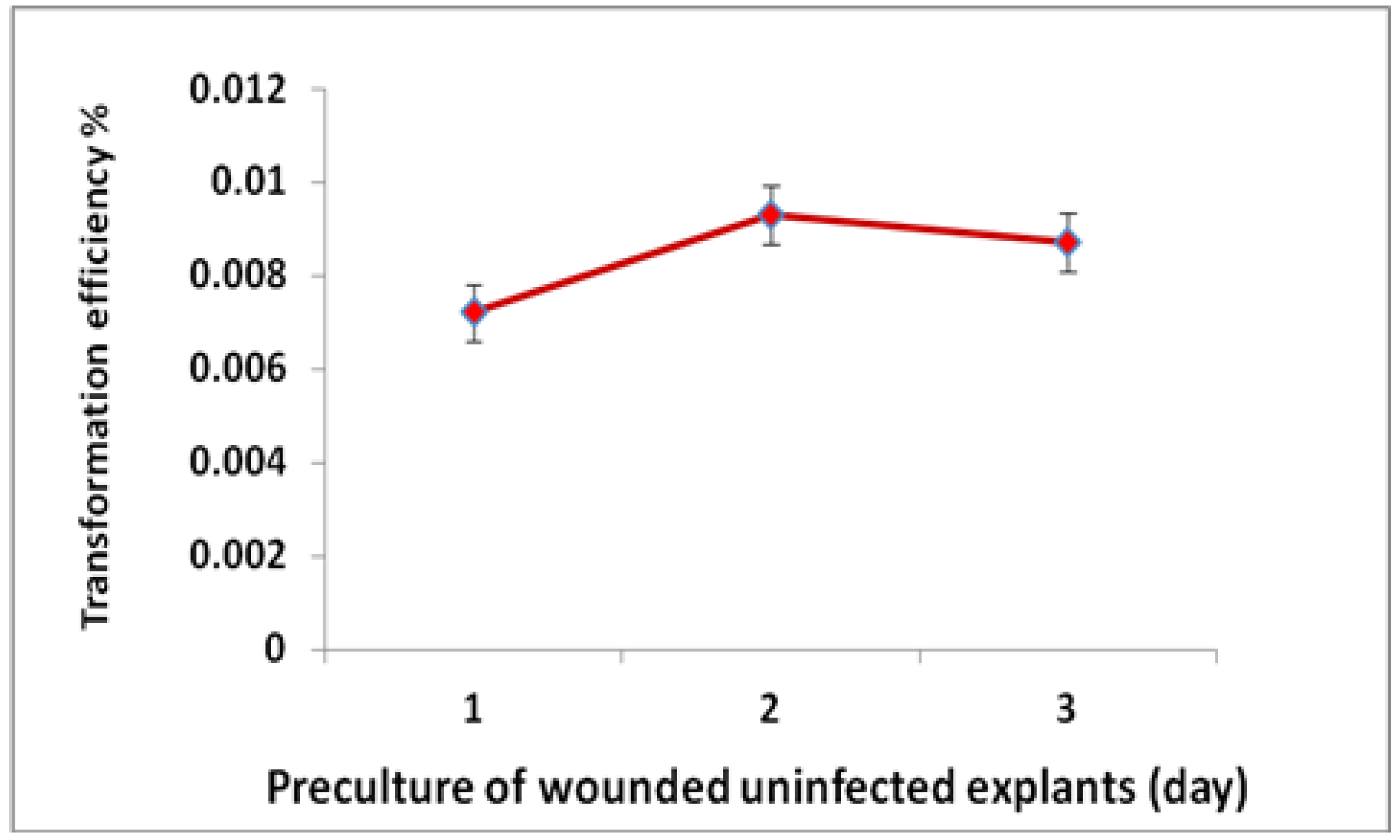
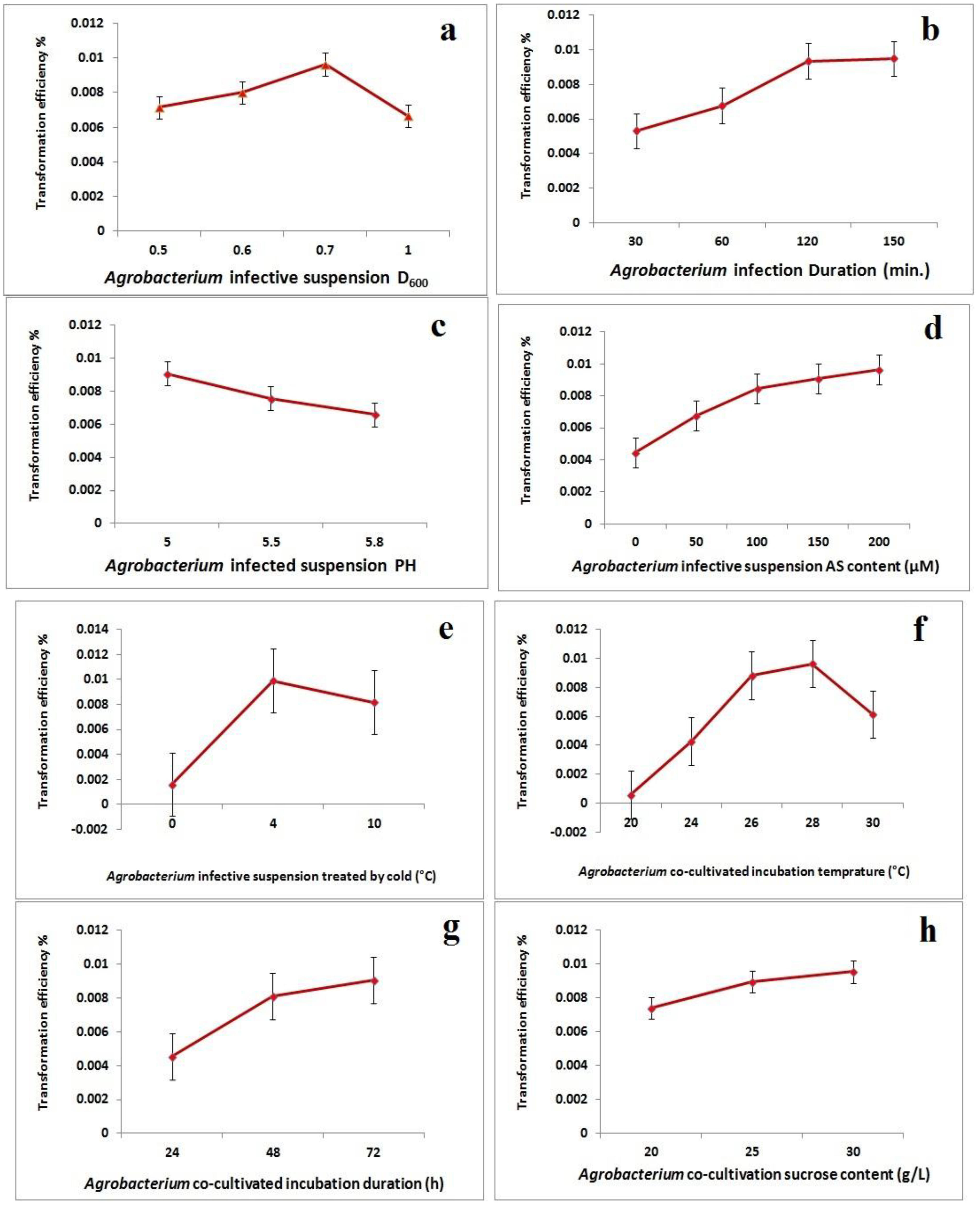
2.2. Improvement of Agrobacterium-Mediated Transformation Efficiency

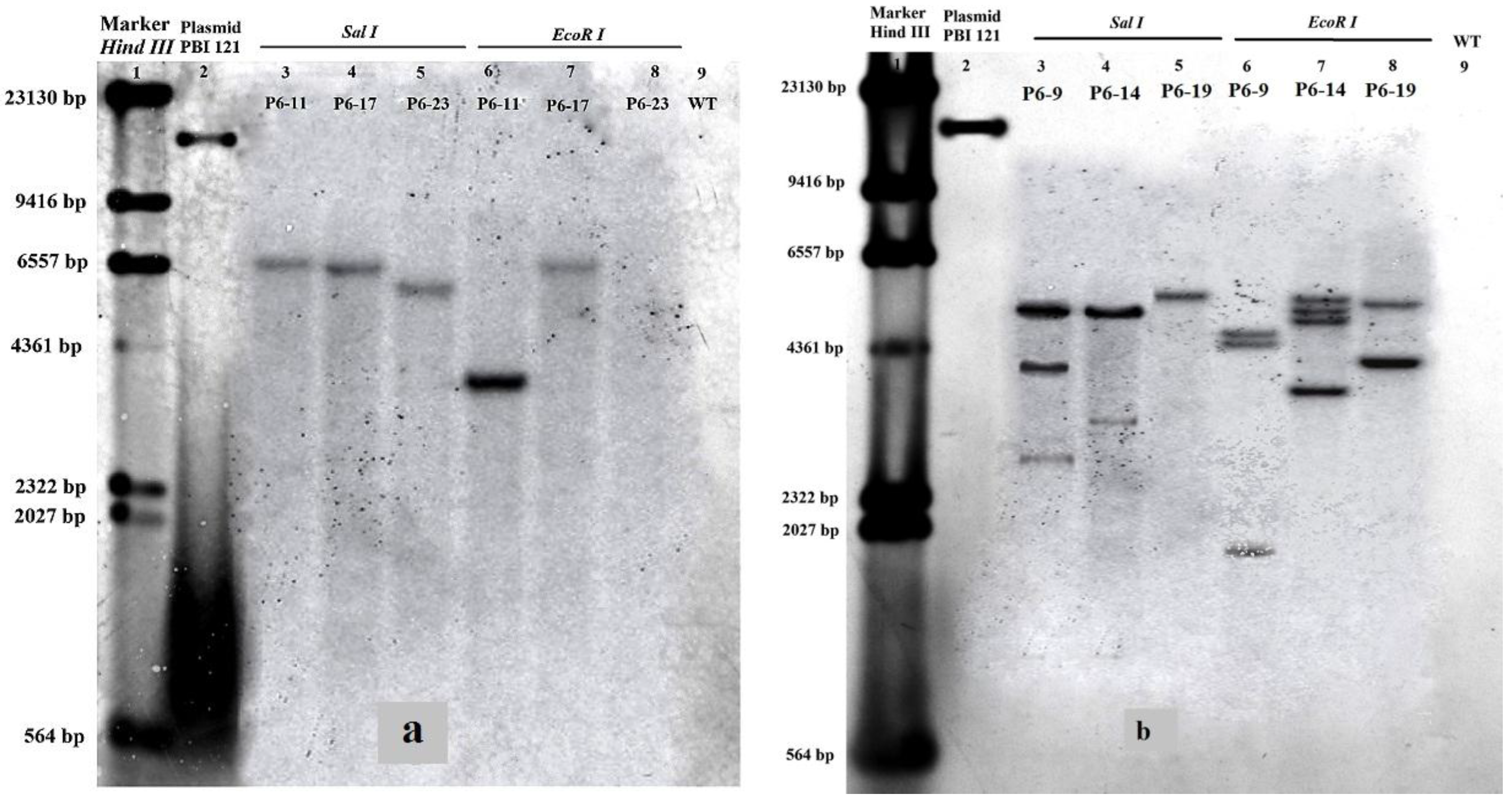
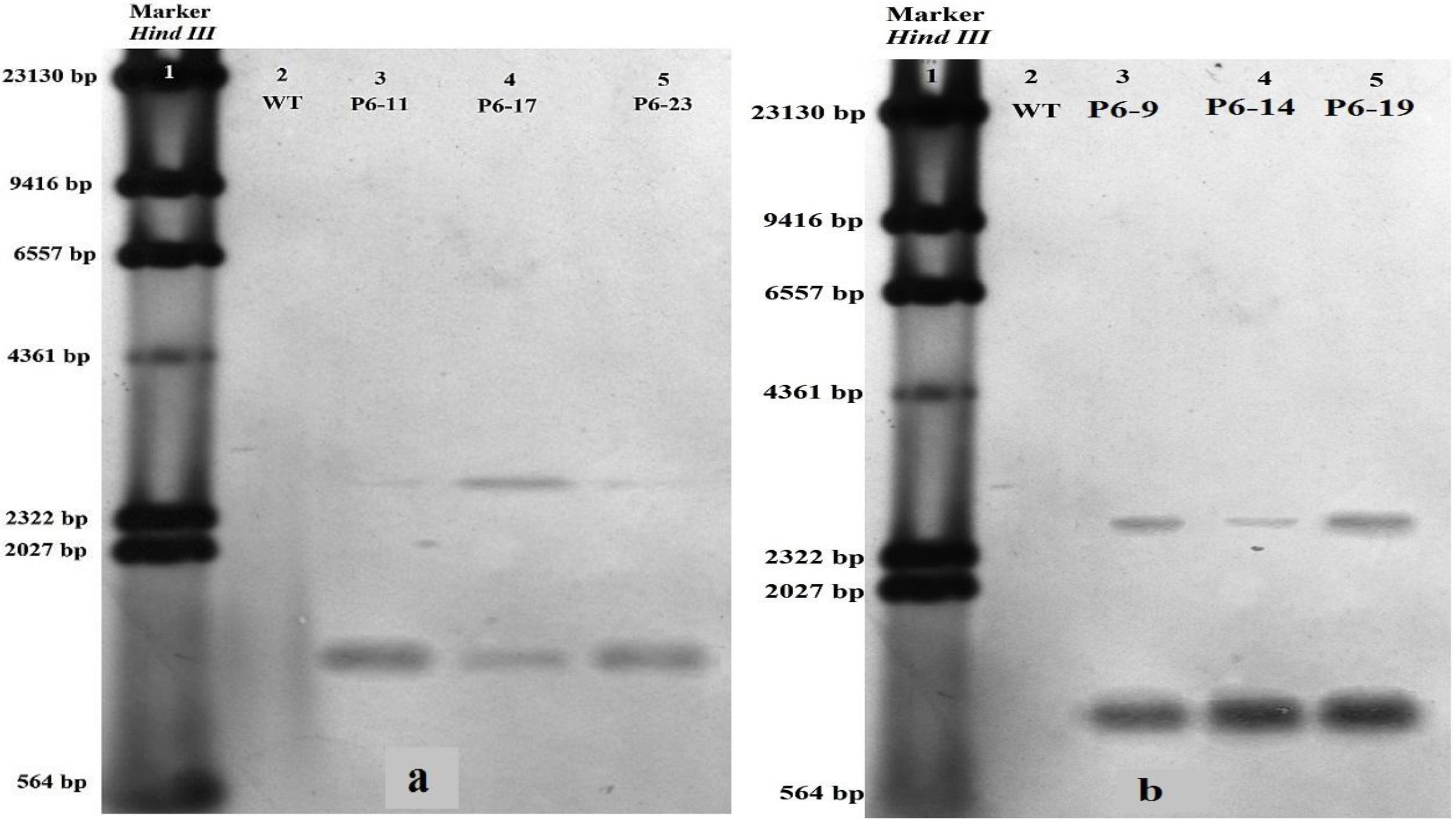
2.3. Southern and Northern Blot Analyses


3. Experimental Section
3.1. Plant Materials and Agrobacterium Strain
3.2. Optimization of Regeneration Medium for Poplar
3.3. Agrobacterium-Mediated Transformation
3.4. Southern and Northern Blot Analyses
3.5. Statistical Analysis
4. Conclusions
Abbreviations
| 6-BA | N-6-benzyladenine |
| AS | acetosyringone |
| NPTII | neomycin phosphotransferase |
| ORF | open reading frame |
| TDZ | thidiazuron |
| Cef | cefotaxime sodium |
| Kan | kanamycin sulfate |
| MS | Murashige and Skoog |
| Rif | rifampicin |
| NAA | naphthaleneacetic acid |
| IAA | indoleacetic acid |
| IBA | indole-3-butyric acid |
| NAM | no apical meristem |
| ATAF1,2 | Arabidopsis transcription activation factor |
| CUC2 | cup-shaped cotyledon |
Acknowledgments
Author Contributions
Conflicts of interest
References
- Confalonieri, M.; Balestrazzi, A.; Bisoffi, S. Genetic transformation of Populus nigra by Agrobacterium tumefaciens. Plant Cell Rep. 1994, 13, 256–261. [Google Scholar]
- Ferreira, S.; Batista, D.; Serrazina, S.; Pais, M.S. Morphogenesis induction and organogenic nodule differentiation in Populus euphratica Oliv. leaf explants. Plant Cell Tissue Organ Cult. 2009, 96, 35–43. [Google Scholar] [CrossRef]
- Gozukirmizi, N.; Bajrovic, K.; Ipekqi, Z. Genotype differences in direct plant regeneration from stem explants of Populus tremula in Turkey. J. For. Res. 1998, 3, 123–126. [Google Scholar] [CrossRef]
- Han, X.; Ma, S.; Kong, X.; Takano, T.; Liu, S. Efficient Agrobacterium-mediated transformation of hybrid poplar Populus davidiana Dode × Populus bollena Lauche. Int. J. Mol. Sci. 2013, 14, 2515–2528. [Google Scholar] [CrossRef]
- Kang, B.; Osburn, L.; Kopsell, D.; Tuskan, G.A.; Cheng, Z.M. Micropropagation of Populus trichocarpa “Nisqually-1”: The genotype deriving the Populus reference genome. Plant Cell Tissue Organ Cult. 2009, 99, 251–257. [Google Scholar] [CrossRef]
- Yadav, R.; Arora, P.; Kumar, D.; Katyal, D.; Dilbaghi, N.; Chaudhury, A. High frequency direct plant regeneration from leaf, internode, and root segments of Eastern Cottonwood (Populus deltoides). Plant Biotechnol. Rep. 2009, 3, 175–182. [Google Scholar] [CrossRef]
- Horsch, R.B.; Fry, J.E.; Hoffmann, N.L.; Eichholtz, D.; Rogers, S.G.; Fraley, R.T. A simple and general method for transferring genes into plants. Science 1985, 227, 1229–1231. [Google Scholar]
- Han, K.H.; Meilan, R.; Ma, C.; Strauss, S.H. An Agrobacterium tumefaciens transformation protocol effective on a variety of cottonwood hybrids (genus Populus). Plant Cell Rep. 2000, 19, 315–320. [Google Scholar]
- Meilan, R.; Ma, C. Poplar (Populus ssp.). In Methods in Molecular Biology, Agrobacterium Protocols, 2nd ed.; Wang, K., Ed.; Humana Press: Totowa, NJ, USA, 2006; pp. 143–151. [Google Scholar]
- Ma, C.P.; Strauss, S.H.; Meilan, R. Agrobacterium-mediated transformation of the genome sequenced poplar clone, Nisqually-1 (Populus trichocarpa). Plant Mol. Biol. Rep. 2004, 22, 1–9. [Google Scholar]
- Yevtushenko, D.P.; Santosh, M. Efficient Agrobacterium-mediated transformation of commercial hybrid poplar Populus nigra L. × P. maximowiczii A. Henry. Plant Cell Rep. 2010, 29, 211–221. [Google Scholar] [CrossRef]
- Husaini, A.M. Pre- and post-agroinfection strategies for efficient leaf disk transformation and regeneration of transgenic strawberry plants. Plant Cell Rep. 2010, 29, 97–110. [Google Scholar] [CrossRef]
- Bartlett, J.G.; Alves, S.C.; Smedley, M.; Snape, J.W.; Harwood, W.A. High-throughput Agrobacterium-mediated barley transformation. Plant Methods 2008, 4, 1–12. [Google Scholar] [CrossRef]
- Kondou, Y.; Higuchi, M.; Takahashi, S. Systematic approaches to using the FOX hunting system to identify useful rice genes. Plant J. 2009, 57, 883–894. [Google Scholar] [CrossRef]
- Nakamura, H.; Hakata, M.; Amano, K.; Miyao, A. A genome-wide gain-of-function analysis of rice genes using the FOX-Hunting system. Plant Mol. Biol. 2007, 65, 357–371. [Google Scholar] [CrossRef]
- Fowler, S.; Thomashow, M.F. Arabidopsis transcriptome profiling indicates that multiple regulatory pathways are activated during cold acclimation in addition to the CBF cold response pathway. Plant Cell 2002, 14, 1675–1690. [Google Scholar] [CrossRef]
- Nakashima, K.; Ito, Y.; Yamaguchi-Shinozaki, K. Transcriptional regulatory networks in response to abiotic stresses in Arabidopsis and grasses. Plant Physiol. 2009, 149, 88–95. [Google Scholar] [CrossRef]
- Nakashima, K.; Takasaki, H.; Mizoi, J.; Shinozaki, K.; Yamaguchi-Shinozaki, K. NAC transcription factors in plant abiotic stress responses. Biochim. Biophys. Acta (BBA) Gene Regul. Mech. 2012, 1819, 97–103. [Google Scholar] [CrossRef]
- Rabbani, M.A.; Maruyama, K.; Abe, H.; Khan, M.A.; Katsura, K.; Ito, Y.; Yoshiwara, K.; Seki, M.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Monitoring expression profiles of rice genes under cold, drought, and high-salinity stresses and abscisic acid application using cDNA microarray and RNA gel-blot analyses. Plant Physiol. 2003, 133, 1755–1767. [Google Scholar] [CrossRef]
- Yamaguchi-Shinozaki, K.; Shinozaki, K. Transcriptional regulatory networks in cellular responses and tolerance to dehydration and cold stresses. Annu. Rev. Plant Biol. 2006, 57, 781–803. [Google Scholar] [CrossRef]
- Holsters, M.; Waele, D.; Depicker, A.; Messens, E.; van Montagu, M.; Schell, J. Transfection and transformation of Agrobacterium tumefaciens. Mol. Gen. Genet. 1978, 163, 181–187. [Google Scholar] [CrossRef]
- Tzfira, T.; Frankmen, L.; Vaidya, M.; Citovsky, V. Site-specific integration of Agrobacterium T-DNA via double-stranded intermediates. Plant Physiol. 2003, 133, 1011–1023. [Google Scholar] [CrossRef]
- Thakur, D.K.; Srivastava, D.K. High-efficiency plant regeneration from leaf explants of male Himalayan poplar (Populus ciliata Wall). In Vitro Cell Dev. Biol. Plant 2006, 42, 144–147. [Google Scholar] [CrossRef]
- Zhan, X.C.; Kawai, S.; Katayama, Y.; Morohoshi, N. A new approach based on the leaf disc method for Agrobacterium mediated transformation and regeneration of aspen. Plant Sci. 1997, 123, 105–112. [Google Scholar] [CrossRef]
- Herrera-Estrella, L.; Simpson, J.; Martinez-Trujillo, M. Transgenic plants: An historical perspective. Methods Mol. Biol. 2005, 286, 3–32. [Google Scholar]
- Hui, P.; Cheng, H.Y.; Chen, C.; Yu, X.W.; Yang, J.N.; Gao, W.R.; Shi, Q.H.; Zhang, H.; Li, J.G.; Ma, H. A NAC transcription factor gene of chickpea (Cicer arietinum), CarNAC3, is involved in drought stress response and various developmental processes. J. Plant Physiol. 2009, 166, 1934–1945. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bioassays with tobacco tissue culture. Plant Physiol. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Horsch, R.B.; Fry, J.; Hoffmann, N.; Neidermeyer, J.; Rogers, S.G.; Fraley, R.T. Leaf disc transformation. In Plant Molecular Biology Manual; Gelvin, S.B., Schilperoort, R.A., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1988; pp. 1–9. [Google Scholar]
- Satchel, S.E.; Messens, E.; van Montagu, M.; Zambryski, P. Identification of the signal molecules produced by wounded plant cells that activate T-DNA transfer in Agrobacterium tumefaciens. Nature 1985, 318, 624–629. [Google Scholar] [CrossRef]
- Doyle, J.J.; Dickson, E.E. Preservation of plant samples for DNA restriction endonuclease analysis. Taxon 1987, 36, 715–722. [Google Scholar] [CrossRef]
- Porebski, L.; Bailey, L.; Baum, B. Modification of a CTAB DNA extraction protocol for plants containing high polysaccharide and polyphenol components. Plant Mol. Biol. Rep. 1997, 15, 8–15. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Movahedi, A.; Zhang, J.; Amirian, R.; Zhuge, Q. An Efficient Agrobacterium-Mediated Transformation System for Poplar. Int. J. Mol. Sci. 2014, 15, 10780-10793. https://doi.org/10.3390/ijms150610780
Movahedi A, Zhang J, Amirian R, Zhuge Q. An Efficient Agrobacterium-Mediated Transformation System for Poplar. International Journal of Molecular Sciences. 2014; 15(6):10780-10793. https://doi.org/10.3390/ijms150610780
Chicago/Turabian StyleMovahedi, Ali, Jiaxin Zhang, Rasoul Amirian, and Qiang Zhuge. 2014. "An Efficient Agrobacterium-Mediated Transformation System for Poplar" International Journal of Molecular Sciences 15, no. 6: 10780-10793. https://doi.org/10.3390/ijms150610780
APA StyleMovahedi, A., Zhang, J., Amirian, R., & Zhuge, Q. (2014). An Efficient Agrobacterium-Mediated Transformation System for Poplar. International Journal of Molecular Sciences, 15(6), 10780-10793. https://doi.org/10.3390/ijms150610780