Contribution of Chitinase A’s C-Terminal Vacuolar Sorting Determinant to the Study of Soluble Protein Compartmentation



{kind=link}
Abstract
:1. Introduction
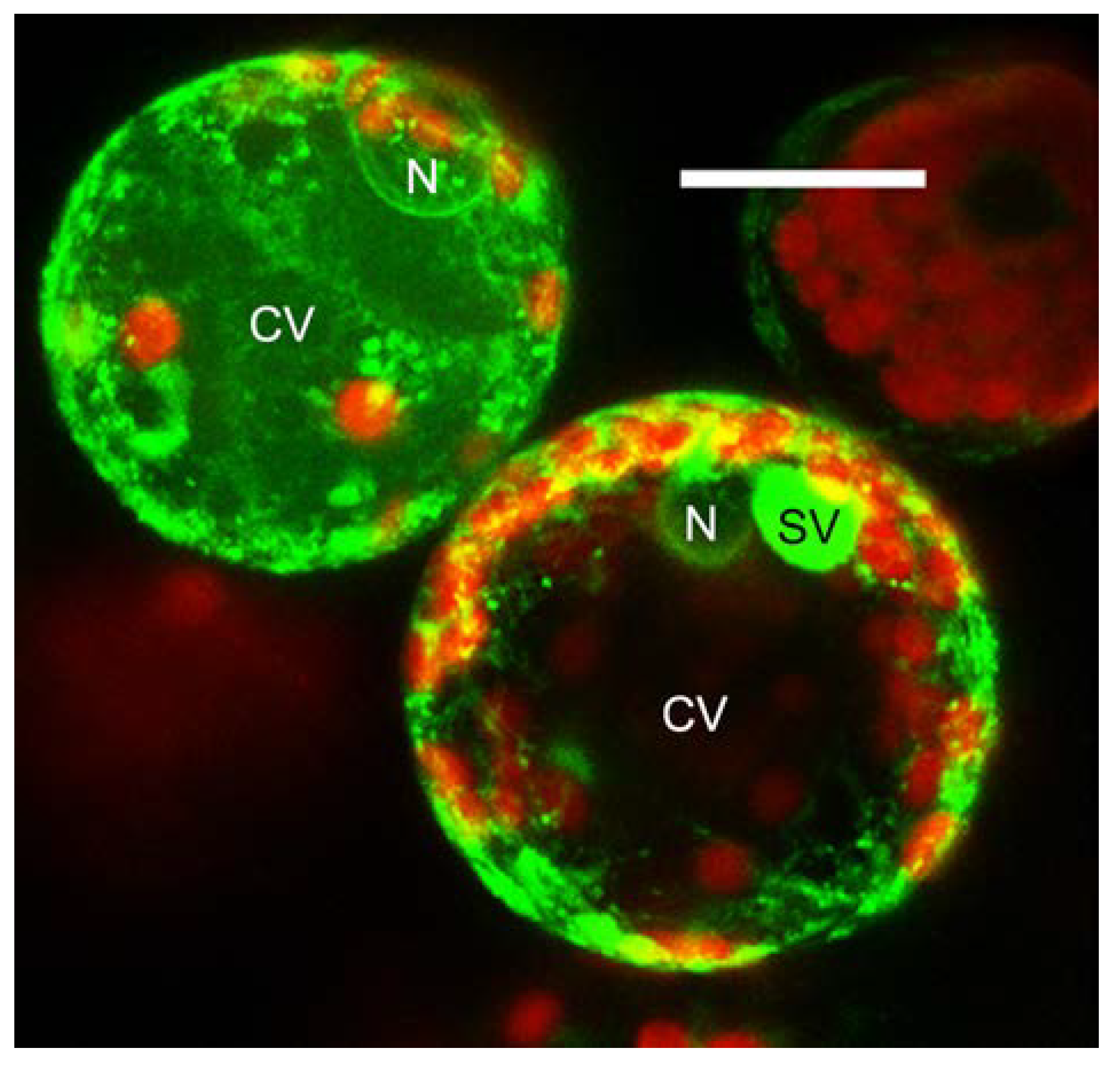
2. Chitinase Sorting, a Hidden Path
3. Conclusions

Acknowledgments
Author Contributions
Conflicts of Interest
References
- Thomas, K.; Aalbers, M.; Bannon, G.A.; Bartels, M.; Dearman, R.J.; Esdaile, D.J.; Fu, T.J.; Glatt, C.M.; Hadfield, N.; Hatzos, C.; et al. A multi-laboratory evaluation of a common in vitro pepsin digestion assay protocol used in assessing the safety of novel proteins. Regul. Toxicol. Pharmacol. 2004, 39, 87–98. [Google Scholar]
- Helenius, A.; Aebi, M. Roles of N-linked glycans in the endoplasmic reticulum. Annu. Rev. Biochem. 2004, 73, 1019–1049. [Google Scholar] [CrossRef]
- Huet, J.; Rucktooa, P.; Clantin, B.; Azarkan, M.; Looze, Y.; Villeret, V.; Wintjens, R. X-ray structure of papaya chitinase reveals the substrate binding mode of glycosyl hydrolase family 19 chitinases. Biochemistry 2008, 47, 8283–8291. [Google Scholar] [CrossRef]
- Ohnishi, T.; Juffer, A.H.; Tamoi, M.; Skriver, K.; Fukamizo, T. 26 kDa endochitinase from barley seeds: An interaction of the ionizable side chains essential for catalysis. J. Biochem. 2005, 138, 553–562. [Google Scholar] [CrossRef]
- Kezuka, Y.; Kojima, M.; Mizuno, R.; Suzuki, K.; Watanabe, T.; Nonaka, T. Structure of full-length class I chitinase from rice revealed by X-ray crystallography and small-angle X-ray scattering. Proteins 2010, 78, 2295–2305. [Google Scholar] [CrossRef]
- Neuhaus, J.-M. Plant chitinases (PR-3, PR-4, PR-8, PR-11). In Pathogenesis-Related Proteins in Plants; Datta, S.K., Muthukrishnan, S., Eds.; The CRC Press: Boca Raton, FL, USA, 1999; pp. 77–105. [Google Scholar]
- Neuhaus, J.-M.; Pietrzak, M.; Boller, T. Mutation analysis of the C-terminal vacuolar targeting peptide of tobacco chitinase: Low specificity of the sorting system, and gradual transition between intracellular retention and secretion into the extracellular space. Plant J. 1994, 5, 45–54. [Google Scholar]
- Di Sansebastiano, G.P.; Paris, N.; Marc-Martin, S.; Neuhaus, J.-M. Specific accumulation of GFP in a non-acidic vacuolar compartment via a C-terminal propeptide-mediated sorting pathway. Plant J. 1998, 15, 449–457. [Google Scholar] [CrossRef]
- Di Sansebastiano, G.; Rehman Reiaz, U.L.; Neuhaus, J.-M. Rat β-glucuronidase as a reporter protein for the analysis of the plant secretory pathway. Plant Biosyst. 2007, 141, 329–336. [Google Scholar] [CrossRef]
- Dombrowski, J.E.; Schroeder, M.R.; Bednarek, S.Y.; Raikhel, N.V. Determination of the functional elements within the vacuolar targeting signal of barley lectin. Plant Cell 1993, 5, 587–596. [Google Scholar] [CrossRef]
- Melchers, L.S.; Sela-Buurlage, M.B.; Vloemans, S.A.; Woloshuk, C.P.; van Roekel, J.S.C.; Pen, J.; van den Elzen, P.J.M.; Cornelissen, B.J.C. Extracellular targeting of the vacuolar tobacco proteins-AP24, chitinase and β-1,3-glucanase in transgenic plants. Plant Mol. Biol. 1993, 21, 583–593. [Google Scholar]
- Neuhaus, J.M.; Sticher, L.; Meins, F.; Boller, T. A short C-terminal sequence is necessary and sufficient for the targeting of chitinases to the plant vacuole. Proc. Natl. Acad. Sci. USA 1991, 88, 10362–10366. [Google Scholar] [CrossRef]
- Sticher, L.; Hinz, U.; Meyer, A.D.; Meins, F. Intracellular transport and processing of a tobacco vacuolar β-1,3-glucanase. Planta 1992, 188, 559–565. [Google Scholar]
- Saalbach, G.; Jung, R.; Kunze, G.; Saalbach, I.; Adler, K.; Müntz, K. Different legumin protein domains act as vacuolar targeting signals. Plant Cell 1991, 3, 695–708. [Google Scholar] [CrossRef]
- Higgins, T.J.V.; Chandler, P.M.; Randall, P.J.; Spencer, D.; Beach, L.R.; Blagrove, R.J.; Kortt, A.A.; Inglis, A.S. Gene structure, protein structure, and regulation of the synthesis of a sulfur-rich protein in pea seeds. J. Biol. Chem. 1986, 261, 11124–11130. [Google Scholar]
- Frigerio, L.; de Virgilio, M.; Prada, A.; Faoro, F.; Vitale, A. Sorting of phaseolin to the vacuole is saturable and requires a short C-terminal peptide. Plant Cell 1998, 10, 1031–1042. [Google Scholar] [CrossRef]
- Stigliano, E.; Faraco, M.; Neuhaus, J.-M.; Montefusco, A.; Dalessandro, G.; Piro, G.; di Sansebastiano, G.P. Two glycosylated vacuolar GFPs are new markers for ER-to-vacuole sorting. Plant Physiol. Biochem. 2013, 73, 337–343. [Google Scholar] [CrossRef]
- Di Sansebastiano, G.P.; Paris, N.; Marc-Martin, S.; Neuhaus, J.-M. Regeneration of a lytic central vacuole and of neutral peripheral vacuoles can be visualized by green fluorescent proteins targeted to either type of vacuoles. Plant Physiol. 2001, 126, 78–86. [Google Scholar] [CrossRef]
- Flückiger, R.; de Caroli, M.; Piro, G.; Dalessandro, G.; Neuhaus, J.-M.; di Sansebastiano, G.P. Vacuolar system distribution in Arabidopsis tissues, visualized using GFP fusion proteins. J. Exp. Bot. 2003, 54, 1577–1584. [Google Scholar] [CrossRef]
- Xiang, L.; Etxeberria, E.; van den Ende, W. Vacuolar protein sorting mechanisms in plants. FEBS J. 2013, 280, 979–993. [Google Scholar] [CrossRef]
- Marty, F. Plant vacuoles. Plant Cell 1999, 11, 587–600. [Google Scholar] [CrossRef]
- Di Sansebastiano, G.; Renna, L.; Gigante, M.; de Caroli, M.; Piro, G.; Dalessandro, G. Green fluorescent protein reveals variability in vacuoles of tree plant species. Biol. Plant. 2007, 51, 49–55. [Google Scholar] [CrossRef]
- Faraco, M.; di Sansebastiano, G.P.; Spelt, K.; Koes, R.E.; Quattrocchio, F.M. One protoplast is not the other! Plant Physiol 2011, 156, 474–478. [Google Scholar]
- Irani, N.G.; Grotewold, E. Light-induced morphological alteration in anthocyanin-accumulating vacuoles of maize cells. BMC Plant Biol. 2005, 5, 7. [Google Scholar] [CrossRef] [Green Version]
- Epimashko, S.; Meckel, T.; Fischer-Schliebs, E.; Lüttge, U.; Thiel, G. Two functionally different vacuoles for static and dynamic purposes in one plant mesophyll leaf cell. Plant J. 2004, 37, 294–300. [Google Scholar] [CrossRef]
- Pedrazzini, E.; Komarova, N.Y.; Rentsch, D.; Vitale, A. Traffic routes and signals for the tonoplast. Traffic 2013, 14, 622–628. [Google Scholar] [CrossRef]
- Robinson, D.G. Response to Rogers letter. Plant Physiol. 2008, 146, 1026. [Google Scholar] [CrossRef]
- Jolliffe, N.A.; Craddock, C.P.; Frigerio, L. Pathways for protein transport to seed storage vacuoles. Biochem. Soc. Trans. 2005, 33, 1016–1018. [Google Scholar] [CrossRef]
- Zouhar, J.; Rojo, E. Plant vacuoles: Where did they come from and where are they heading? Curr. Opin. Plant Biol. 2009, 12, 677–684. [Google Scholar] [CrossRef]
- Hatsugai, N.; Hara-Nishimura, I. Two vacuole-mediated defense strategies in plants. Plant Signal. Behav. 2010, 5, 1568–1570. [Google Scholar] [CrossRef]
- De Marchis, F.; Bellucci, M.; Pompa, A. Unconventional pathways of secretory plant proteins from the endoplasmic reticulum to the vacuole bypassing the Golgi complex. Plant Signal. Behav. 2013, 8, e25129. [Google Scholar] [CrossRef]
- Viotti, C.; Krüger, F.; Krebs, M.; Neubert, C.; Fink, F.; Lupanga, U.; Scheuring, D.; Boutté, Y.; Frescatada-Rosa, M.; Wolfenstetter, S.; et al. The endoplasmic reticulum is the main membrane source for biogenesis of the lytic vacuole in Arabidopsis. Plant Cell 2013, 25, 3434–3449. [Google Scholar] [CrossRef]
- Matile, P.; Moor, H. Vacuolation: Origin and development of the lysosomal apparatus in root-tip cells. Planta 1968, 80, 159–175. [Google Scholar] [CrossRef]
- Mesquita, J.F. Electron microscope study of the origin and development of the vacuoles in root-tip cells of Lupinusalbus L. J. Ultrastruct. Res. 1969, 26, 242–250. [Google Scholar] [CrossRef]
- Poustka, F.; Irani, N.G.; Feller, A.; Lu, Y.; Pourcel, L.; Frame, K.; Grotewold, E. A trafficking pathway for anthocyanins overlaps with the endoplasmic reticulum-to-vacuole protein-sorting route in Arabidopsis and contributes to the formation of vacuolar inclusions. Plant Physiol. 2007, 145, 1323–1335. [Google Scholar] [CrossRef]
- Robinson, D.G.; Jiang, L.; Schumacher, K. The endosomal system of plants: Charting new and familiar territories. Plant Physiol. 2008, 147, 1482–1492. [Google Scholar] [CrossRef]
- Geldner, N.; Anders, N.; Wolters, H.; Keicher, J.; Kornberger, W.; Muller, P.; Delbarre, A.; Ueda, T.; Nakano, A.; Jürgens, G. The Arabidopsis GNOM ARF–GEF mediates endosomal recycling, auxin transport, and auxin-dependent plant growth. Cell 2003, 112, 219–230. [Google Scholar] [CrossRef]
- Di Sansebastiano, G.P.; Fornaciari, S.; Barozzi, F.; Piro, G.; Arru, L. New insights on plant cell elongation: A role for acetylcholine. Int. J. Mol. Sci. 2014, 15, 4565–4582. [Google Scholar] [CrossRef]
- Zouhar, J.; Rojo, E.; Bassham, D.C. AtVPS45 is a positive regulator of the SYP41/SYP61/VTI12 SNARE complex involved in trafficking of vacuolar cargo. Plant Physiol. 2009, 149, 1668–1678. [Google Scholar] [CrossRef]
- Sanmartin, M.; Ordonez, A.; Sohn, E.J.; Robert, S.; Sanchez-Serrano, J.J.; Surpin, M.A.; Raikhel, N.V.; Rojo, E. Divergent functions of VTI12 and VTI11 in trafficking to storage and lytic vacuoles in Arabidopsis. Proc. Natl. Acad. Sci. USA 2007, 104, 3645–3650. [Google Scholar] [CrossRef]
- Viotti, C. ER and vacuoles: Never been closer. Front. Plant Sci. 2014, 5, 20. [Google Scholar] [CrossRef]
- Ebine, K.; Okatani, Y.; Uemura, T.; Goh, T.; Shoda, K.; Niihama, M.; Morita, M.T.; Spitzer, C.; Otegui, M.S.; Nakano, A.; et al. A SNARE complex unique to seed plants is required for protein storage vacuole biogenesis and seed development of Arabidopsis thaliana. Plant Cell 2008, 20, 3006–3021. [Google Scholar] [CrossRef]
- De Benedictis, M.; Bleve, G.; Faraco, M.; Stigliano, E.; Grieco, F.; Piro, G.; Dalessandro, G.; di Sansebastiano, G.P. AtSYP51/52 functions diverge in the post-Golgi traffic and differently affect vacuolar sorting. Mol. Plant 2013, 6, 916–930. [Google Scholar] [CrossRef]
- Faraco, M.; Latino, A.; de Benedictis, M.; Piro, G.; di Sansebastiano, G.P. Atsyp51 and Atsyp52 differently affect sorting of GFPChi and AleuGFP vacuolar markers. J. Plant Biochem. Physiol. 2013, 1, 2. [Google Scholar]
- Di Sansebastiano, G.P. Defining new SNARE functions: The i-SNARE. Front. Plant Sci. 2013, 4, 99. [Google Scholar] [CrossRef]
- Kang, B.H.; Nielsen, E.; Preuss, M.L.; Mastronarde, D.; Staehelin, L.A. Electron tomography of RabA4b-and PI-4Kβ1-labeled trans Golgi network compartments in Arabidopsis. Traffic 2011, 12, 313–329. [Google Scholar] [CrossRef]
- Cao, X.; Rogers, S.W.; Butler, J.; Beevers, L.; Rogers, J.C. Structural requirements for ligand binding by a probable plant vacuolar sorting receptor. Plant Cell 2000, 12, 493–506. [Google Scholar] [CrossRef]
- Ahmed, S.U.; Bar-Peled, M.; Raikhel, N.V. Cloning and subcellular location of an Arabidopsis receptor-like protein that shares common features with protein-sorting receptors of eukaryotic cells. Plant Physiol. 1997, 114, 325–336. [Google Scholar]
- Jiang, L.; Phillips, T.E.; Rogers, S.W.; Rogers, J.C. Biogenesis of the protein storage vacuole crystalloid. J. Cell Biol. 2000, 150, 755–770. [Google Scholar] [CrossRef]
- Park, M.; Kim, S.J.; Vitale, A.; Hwang, I. Identification of the protein storage vacuole and protein targeting to the vacuole in leaf cells of three plant species. Plant Physiol. 2004, 134, 625–639. [Google Scholar] [CrossRef]
- Park, M.; Lee, D.; Lee, G.J.; Hwang, I. AtRMR1 functions as a cargo receptor for protein trafficking to the protein storage vacuole. J. Cell Biol. 2005, 170, 757–767. [Google Scholar] [CrossRef]
- Shen, Y.; Wang, J.; Ding, Y.; Lo, S.W.; Gouzerh, G.; Neuhaus, J.-M.; Jiang, L. The rice RMR1 associates with a distinct prevacuolar compartment for the protein storage vacuole pathway. Mol. Plant 2011, 4, 854–868. [Google Scholar] [CrossRef]
- Wang, H.; Rogers, J.C.; Jian, L. Plant RMR proteins: Unique vacuolar sorting receptors that couple ligand sorting with membrane internalization. FEBS J. 2011, 278, 59–68. [Google Scholar] [CrossRef]
- Castelli, S.; Vitale, A. The phaseolin vacuolar sorting signal promotes transient, strong membrane association and aggregation of the bean storage protein in transgenic tobacco. J. Exp. Bot. 2005, 56, 1379–1387. [Google Scholar] [CrossRef]
- Stigliano, E.; University of Neuchâtel, Neuchâtel, Switzerland. Unpublished work, 2014.
- Scabone, C.M.; Frigerio, L.; Petruccelli, S. A fluorescent reporter protein containing AtRMR1 domains is targeted to the storage and central vacuoles in Arabidopsis thaliana and tobacco leaf cells. Plant Cell Rep. 2011, 30, 1823–1833. [Google Scholar]
- Hara-Hishimura, I.; Takeuchi, Y.; Inoue, K.; Nishimura, M. Vesicle transport and processing of the precursor to 2S albumin in pumpkin. Plant J. 1993, 4, 793–800. [Google Scholar]
- Batistic, O.; Kudla, J. Plant calcineurin B-like proteins and their interacting protein kinases. Biochim. Biophys. Acta 2009, 1793, 985–992. [Google Scholar] [CrossRef]
- Levanony, H.; Rubln, R.; Altschuler, Y.; Gallli, G. Evidente for a novel route of wheat storage proteins to vacuoles. J. Cell Biol. 1992, 119, 1117–1128. [Google Scholar] [CrossRef]
- Takahashi, H.; Saito, Y.; Kitagawa, T.; Morita, S.; Masumura, T.; Tanaka, K. A novel vesicle derived directly from endoplasmic reticulum is involved in the transport of vacuolar storage proteins in rice endosperm. Plant Cell Physiol. 2005, 46, 245–249. [Google Scholar] [CrossRef]
- Reyes, F.C.; Chung, T.; Holding, D.; Jung, R.; Vierstra, R.; Otegui, M.S. Delivery of prolamins to the protein storage vacuole in maize aleurone cells. Plant Cell 2011, 23, 769–784. [Google Scholar] [CrossRef]
- Rivera-Serrano, E.E.; Rodriguez-Welsh, M.F.; Hicks, G.R.; Rojas-Pierce, M. A small molecule inhibitor partitions two distinct pathways for trafficking of tonoplast intrinsic proteins in Arabidopsis. PLoS One 2012, 7, e44735. [Google Scholar]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Stigliano, E.; Di Sansebastiano, G.-P.; Neuhaus, J.-M. Contribution of Chitinase A’s C-Terminal Vacuolar Sorting Determinant to the Study of Soluble Protein Compartmentation. Int. J. Mol. Sci. 2014, 15, 11030-11039. https://doi.org/10.3390/ijms150611030
Stigliano E, Di Sansebastiano G-P, Neuhaus J-M. Contribution of Chitinase A’s C-Terminal Vacuolar Sorting Determinant to the Study of Soluble Protein Compartmentation. International Journal of Molecular Sciences. 2014; 15(6):11030-11039. https://doi.org/10.3390/ijms150611030
Chicago/Turabian StyleStigliano, Egidio, Gian-Pietro Di Sansebastiano, and Jean-Marc Neuhaus. 2014. "Contribution of Chitinase A’s C-Terminal Vacuolar Sorting Determinant to the Study of Soluble Protein Compartmentation" International Journal of Molecular Sciences 15, no. 6: 11030-11039. https://doi.org/10.3390/ijms150611030
APA StyleStigliano, E., Di Sansebastiano, G. -P., & Neuhaus, J. -M. (2014). Contribution of Chitinase A’s C-Terminal Vacuolar Sorting Determinant to the Study of Soluble Protein Compartmentation. International Journal of Molecular Sciences, 15(6), 11030-11039. https://doi.org/10.3390/ijms150611030