Development of an Intergeneric Conjugal Transfer System for Xinaomycins-Producing Streptomyces noursei Xinao-4

Abstract
:1. Introduction
2. Results and Discussion
2.1. Antibiotic Tolerance of S. noursei Xinao-4
2.2. Optimization of Solid Media for Exconjugants Regeneration

2.3. Effect of Heat-Shock on the Conjugation Efficiency

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Temperature (°C) | Viability (%) | Conjugation Frequency |
|---|---|---|
| Room temperature | 100 | (4.87 ± 0.31) × 10−5 |
| 40 | 95.5 | (8.0 ± 0.56) × 10−5 |
| 45 | 69.87 | (1.96 ± 0.29) × 10−4 |
| 50 | 56.12 | (3.47 ± 0.21) × 10−4 |
| 55 | 17.97 | (7.1 ± 0.45) × 10−5 |

2.4. Effect of Donor-to-Recipient Ratio on Conjugation Efficiency
| Donor Numbers (×107) | Conjugation Frequency |
|---|---|
| 1 | (6.59 ± 0.29) × 10−5 |
| 10 | (5.5 ± 0.27) × 10−4 |
| 100 | (2.3± 0.24) × 10−3 |
| 1000 | (4.4 ± 0.2) × 10−4 |
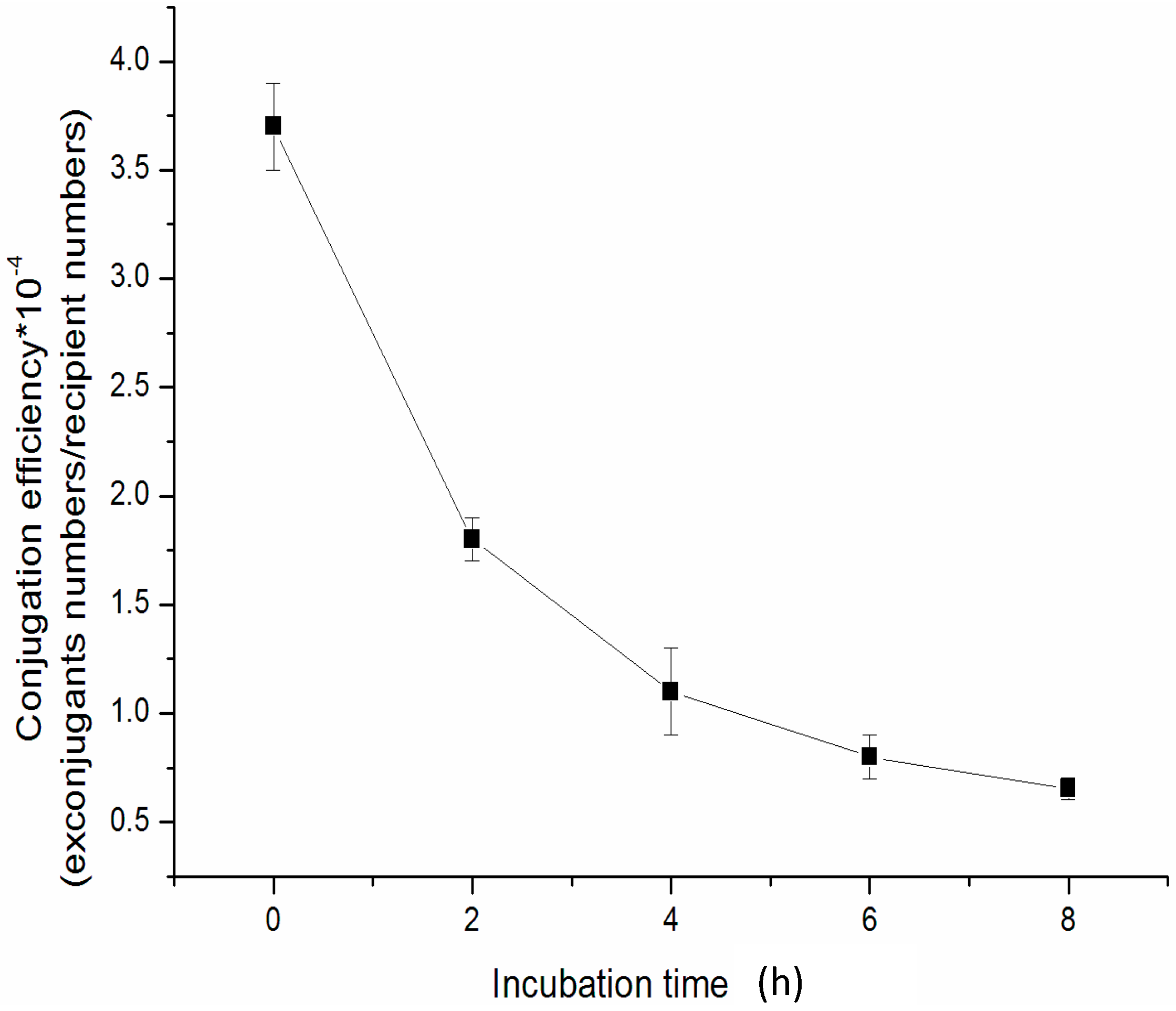
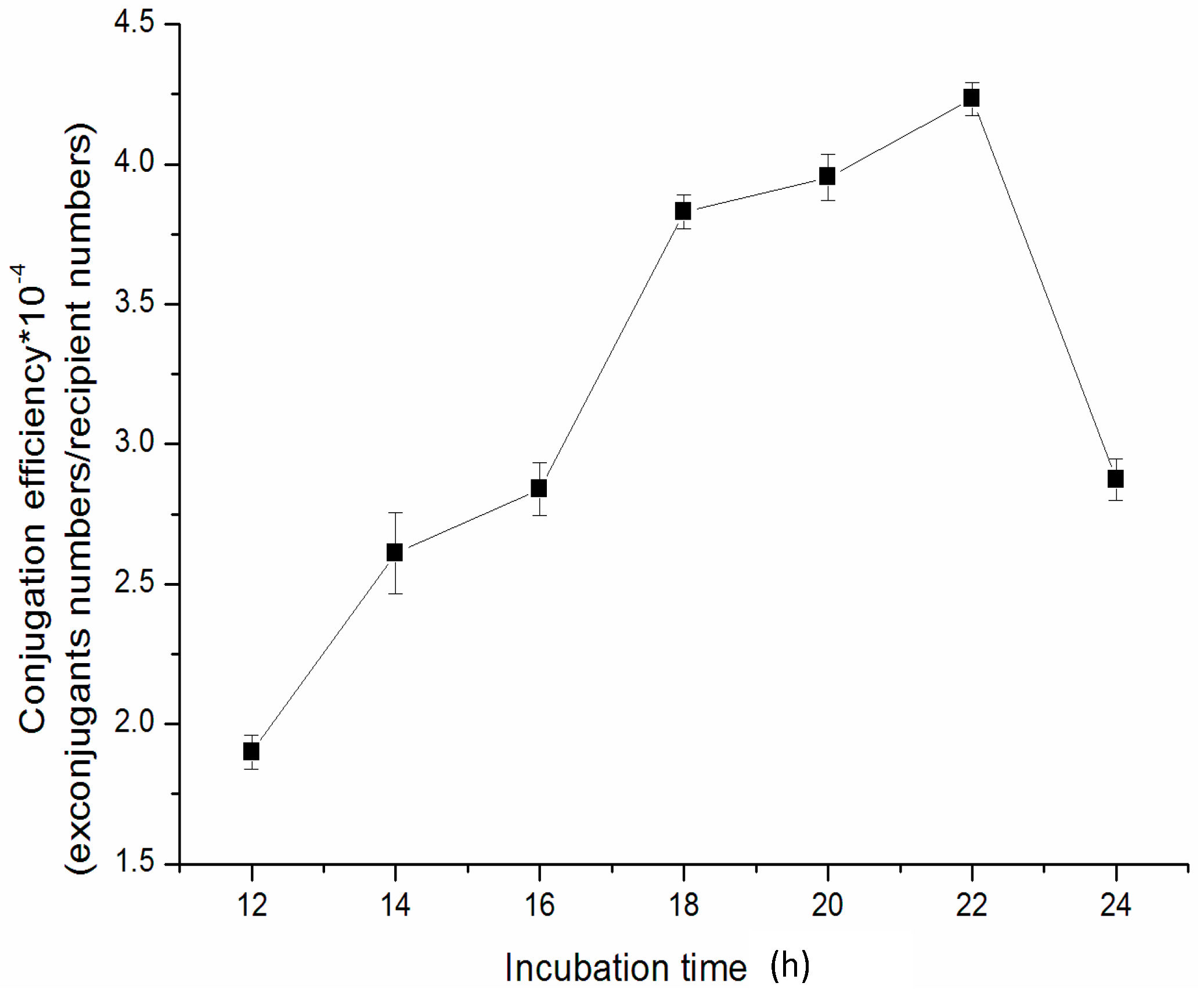
2.5. The Effect of Incubation Time of Mix-Culture on the Conjugation Efficiency

2.6. Conjugal Transfers of Integrated and Self-Replicative Plasmid into S. noursei Xinao-4
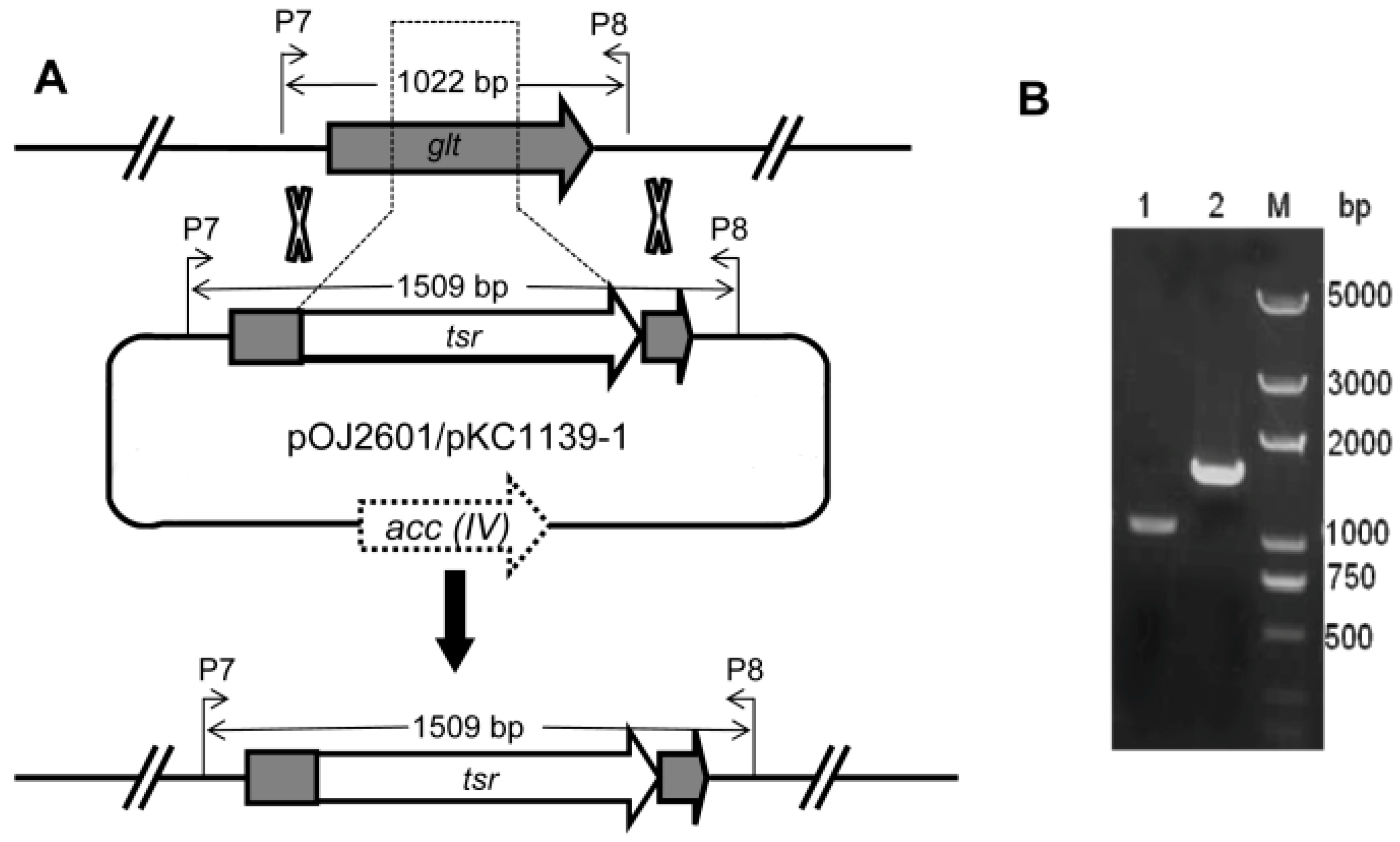
2.7. Validation of Conjugation System: A Case Study

3. Experimental Section
3.1. Strains and Plasmids
3.2. Media and Culture Condition
3.3. Construction of Plasmids for Gene Deletion
3.4. Intergeneric Conjugation Procedure and Analysis of Exconjugants
3.5. Analysis of Exconjugants
3.6. Fermentations and Bioassay of Xinaomycins
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Phornphisutthimas, S.; Sudtachat, N.; Bunyoo, C.; Chotewutmontri, P.; Panijpan, B.; Thamchaipenet, A. Development of an intergeneric conjugal transfer system for rimocidin-producing Streptomyces rimosus. Lett. Appl. Microbiol. 2010, 50, 530–536. [Google Scholar] [CrossRef]
- Hopwood, D.A. Forty years of genetics with Streptomyces: From in vivo through in vitro to in silico. Microbiology 1999, 145, 2183–2202. [Google Scholar]
- Li, L.; Xu, Z.; Xu, X.; Wu, J.; Zhang, Y.; He, X.; Zabriskie, T.M.; Deng, Z. The mildiomycin biosynthesis: Initial steps for sequential generation of 5-hydroxymethylcytidine 5'-monophosphate and 5-hydroxymethylcytosine in Streptoverticillium rimofaciens ZJU5119. ChemBioChem 2008, 9, 1286–1294. [Google Scholar] [CrossRef]
- Isono, K. Nucleoside antibiotics: Structure, biological activity, and biosynthesis. J. Antibiot. 1988, 41, 1711–1739. [Google Scholar] [CrossRef]
- Bormann, C.; Möhrle, V.; Bruntner, C. Cloning and heterologous expression of the entire set of structural genes for nikkomycin synthesis from Streptomyces tendae Tü901 in Streptomyces lividans. J. Bacteriol. 1996, 178, 1216–1218. [Google Scholar]
- Wu, J.; Li, L.; Deng, Z.; Zabriskie, T.M.; He, X. Analysis of the mildiomycin biosynthesis gene cluster in Streptoverticillum rimofaciens ZJU5119 and characterization of MilC, a hydroxymethylcytosyl-glucuronic acid synthase. ChemBioChem 2012, 13, 1613–1621. [Google Scholar]
- Niu, G.; Li, L.; Wei, J.; Tan, H. Cloning, heterologous expression, and characterization of the gene cluster required for gougerotin biosynthesis. Chem. Biol. 2013, 20, 34–44. [Google Scholar] [CrossRef]
- Cone, M.C.; Yin, X.; Grochowski, L.L.; Parker, M.R.; Zabriskie, T.M. The blasticidin S biosynthesis gene cluster from Streptomyces griseochromogenes: Sequence analysis, organization, and initial characterization. ChemBioChem 2003, 4, 821–828. [Google Scholar] [CrossRef]
- Tercero, J.A.; Espinosa, J.C.; Lacalle, R.A.; Jiménez, A. The biosynthetic pathway of the aminonucleoside antibiotic puromycin, as deduced from the molecular analysis of the pur cluster of Streptomyces alboniger. J. Biol. Chem. 1996, 271, 1579–1590. [Google Scholar]
- Chen, W.; Huang, T.; He, X.; Meng, Q.; You, D.; Bai, L.; Li, J.; Wu, M.; Li, R.; Xie, Z. Characterization of the polyoxin biosynthetic gene cluster from Streptomyces cacaoi and engineered production of polyoxin H. J. Biol. Chem. 2009, 284, 10627–10638. [Google Scholar]
- Fernandez-Moreno, M.A.; Vallin, C.; Malpartida, F. Streptothricin biosynthesis is catalyzed by enzymes related to nonribosomal peptide bond formation. J. Bacteriol. 1997, 179, 6929–6936. [Google Scholar]
- Ning, T.B.; Zhang, T.; Zhou, J.Y.; Yang, J.; Hong, T. Extraction methods and physicochemical properties of fermentation broth of xinaomycin. Hunan Agric. Sci. 2011, 13, 23–25. [Google Scholar]
- Gu, G.J.; Cui, L.K.; Wang, S.H.; Li, F.X.; Lin, X.M. Toxicity test of xinaomycin to seven plant pathogenic fungi in laboratory. Agrochemicals 2013, 52, 293–294. [Google Scholar]
- Li, F.G.; Yang, F.; Feng, Z.Q.; Lu, Q. Field efficacy of 4% xinaomycin AS on tomato mosaix virus disease. J. Henan Agric. Sci. 2013, 42, 86–88. [Google Scholar]
- Feng, Z.Q.; Lu, Q. Field efficacy of new bio-fungicide xinaomycin on tobacco mosaic virus. Mod. Agrochem. 2011, 10, 50–52. [Google Scholar]
- Zhou, J.Y.; Hong, T.; Juan, Z.; Li, Z.D.; Yang, J.; Xiao, L.; Liu, D. Antimicrobial Compound and Preparation Method and Application. CN Patent 200910060121.4, 9 February 2011. [Google Scholar]
- Matsushima, P.; Baltz, R.H. Efficient plasmid transformation of Streptomyces ambofaciens and Streptomyces fradiae protoplasts. J. Bacteriol. 1985, 163, 180–185. [Google Scholar]
- Voeykova, T.; Emelyanova, L.; Tabakov, V.; Mkrtumyan, N. Transfer of plasmid pTO1 from Escherichia coli to various representatives of the order Actinomycetales by intergeneric conjugation. FEMS Microbiol. Lett. 1998, 162, 47–52. [Google Scholar]
- Mazodier, P.; Petter, R.; Thompson, C. Intergeneric conjugation between Escherichia coli and Streptomyces species. J. Bacteriol. 1989, 171, 3583–3585. [Google Scholar]
- Bierman, M.; Logan, R.; O’Brien, K.; Seno, E.T.; Nagaraja Rao, R.; Schoner, B.E. Plasmid cloning vectors for the conjugal transfer of DNA from Escherichia coli to Streptomyces spp. Gene 1992, 116, 43–49. [Google Scholar] [CrossRef]
- Flett, F.; Mersinias, V.; Smith, C.P. High efficiency intergeneric conjugal transfer of plasmid DNA from Escherichia coli to methyl DNA-restricting Streptomycetes. FEMS Microbiol. Lett. 1997, 155, 223–229. [Google Scholar]
- Kieser, T.; Bibb, M.J.; Chater, K.F.; Butter, M.J.; Hopwood, D.A. Practical Streptomyces Genetics: A Laboratory Manual; John Innes Foundation: Norwich, UK, 2000. [Google Scholar]
- Gust, B.; Challis, G.L.; Fowler, K.; Kieser, T.; Chater, K.F. PCR-targeted Streptomyces gene replacement identifies a protein domain needed for biosynthesis of the sesquiterpene soil odor geosmin. Proc. Natl. Acad. Sci. USA 2003, 100, 1541–1546. [Google Scholar] [CrossRef]
- Du, L.; Liu, R.-H.; Ying, L.; Zhao, G.-R. An efficient intergeneric conjugation of DNA from Escherichia coli to mycelia of the lincomycin-producer Streptomyces lincolnensis. Int. J. Mol. Sci. 2012, 13, 4797–4806. [Google Scholar] [CrossRef]
- Nikodinovic, J.; Barrow, K.D.; Chuck, J.-A. High frequency transformation of the amphotericin-producing bacterium Streptomyces nodosus. J. Microbiol. Methods 2003, 55, 273–277. [Google Scholar] [CrossRef]
- Guan, D.; Pettis, G. Intergeneric conjugal gene transfer from Escherichia coli to the sweet potato pathogen Streptomyces ipomoeae. Lett. Appl. Microbiol. 2009, 49, 67–72. [Google Scholar] [CrossRef]
- Jin, Z.; Jin, X.; Jin, Q. Conjugal transferring of resistance gene ptr for improvement of pristinamycin-producing Streptomyces pristinaespiralis. Appl. Biochem. Biotechnol. 2010, 160, 1853–1864. [Google Scholar] [CrossRef]
- Park, H.Y.; Jang, B.Y.; Hwang, Y.I.; Choi, S.U. Construction of intergeneric conjugal transfer for molecular genetic studies of Streptomyces acidiscabies producing thaxtomin. J. Korean Soc. Appl. Biol. Chem. 2012, 55, 265–269. [Google Scholar]
- Bailey, C.R.; Winstanley, D.J. Inhibition of restriction in Streptomyces clavuligerus by heat treatment. J. Gen. Microbiol. 1986, 132, 2945–2947. [Google Scholar]
- Engel, P. Plasmid transformation of Streptomyces tendae after heat attenuation of restriction. Appl. Environ. Microbiol. 1987, 53, 1–3. [Google Scholar]
- Ma, Z.; Liu, J.; Bechthold, A.; Tao, L.; Shentu, X.; Bian, Y.; Yu, X. Development of intergeneric conjugal gene transfer system in Streptomyces diastatochromogenes 1628 and its application for improvement of toyocamycin production. Curr. Microbiol. 2013, 68, 180–185. [Google Scholar]
- Choi, S.U.; Lee, C.K.; Hwang, Y.I.; Kinoshita, H.; Nihira, T. Intergeneric conjugal transfer of plasmid DNA from Escherichia coli to Kitasatospora setae, a bafilomycin b1 producer. Arch. Microbiol. 2004, 181, 294–298. [Google Scholar] [CrossRef]
- Enríquez, L.L.; Mendes, M.V.; Antón, N.; Tunca, S.; Guerra, S.M.; Martín, J.F.; Aparicio, J.F. An efficient gene transfer system for the pimaricin producer Streptomyces natalensis. FEMS Microbiol. Lett. 2006, 257, 312–318. [Google Scholar]
- Choi, S.H.; Kim, H.S.; Yoon, Y.J.; Kim, D.M.; Lee, E.Y. Glycosyltransferase and its application to glycodiversification of natural products. J. Ind. Eng. Chem. 2012, 18, 1208–1212. [Google Scholar] [CrossRef]
- Chen, R.; Zhang, H.; Zhang, G.; Li, S.; Zhang, G.; Zhu, Y.; Liu, J.; Zhang, C. Characterizing amosamine biosynthesis in amicetin reveals AmiG as a reversible retaining glycosyltransferase. J. Am. Chem. Soc. 2013, 135, 12152–12155. [Google Scholar] [CrossRef]
- Wei, Y.Y. Gene Transformation of Streptomyces ahygroscopicus, Cloning and Function of Cytosinpeptidemycin Biosynthetic Genes. Ph.D. Thesis, Shenyang University, Shenyang, China, 15 June 2009. [Google Scholar]
- Zhang, G.; Zhang, H.; Li, S.; Xiao, J.; Zhang, G.; Zhu, Y.; Niu, S.; Ju, J.; Zhang, C. Characterization of the amicetin biosynthesis gene cluster from Streptomyces vinaceusdrappus NRRL 2363 implicates two alternative strategies for amide bond formation. Appl. Environ. Microbiol. 2012, 78, 2393–2401. [Google Scholar]
- Hanahan, D. Studies on transformation of Escherichia coli with plasmids. J. Mol. Biol. 1983, 166, 557–580. [Google Scholar] [CrossRef]
- MacNeil, D.J.; Gewain, K.M.; Ruby, C.L.; Dezeny, G.; Gibbons, P.H.; MacNeil, T. Analysis of Streptomyces avermitilis genes required for avermectin biosynthesis utilizing a novel integration vector. Gene 1992, 111, 61–68. [Google Scholar] [CrossRef]
- Paget, M.S.; Chamberlin, L.; Atrih, A.; Foster, S.J.; Buttner, M.J. Evidence that the extracytoplasmic function sigma factor sigma ςE is required for normal cell wall structure in Streptomyces coelicolor A3(2). J. Bacteriol. 1999, 181, 204–211. [Google Scholar]
- Vara, J.; Lewandowska-Skarbek, M.; Wang, Y.-G.; Donadio, S.; Hutchinson, C. Cloning of genes governing the deoxysugar portion of the erythromycin biosynthesis pathway in Saccharopolyspora erythraea (Streptomyces erythreus). J. Bacteriol. 1989, 171, 5872–5881. [Google Scholar]
- Sambrook, J.; Russell, D. Molecular Cloning: A Laboratory Manual, 3rd ed.; Cold Spring Harbor Laboratory Press: New York, NY, USA, 2001. [Google Scholar]
- Hobbs, G.; Frazer, C.M.; Gardner, D.C.; Cullum, J.A.; Oliver, S.G. Dispersed growth of Streptomyces in liquid culture. Appl. Microbiol. Biotechnol. 1989, 31, 272–277. [Google Scholar]
- Shirling, E.B.; Gottlieb, D. Method for characterization of Streptomyces species. Int. J. Syst. Evolut. Microbiol. 1966, 16, 313–340. [Google Scholar]
- Xiao, Y.; Li, S.M.; Ma, L.; Zhang, G.Y.; Ju, J.H.; Zhang, C.S. Genetic manipulation system for tiacumicin producer Dactylosporangium aurantiacum NRRL 18085. Acta Microbiol. Sin. 2010, 50, 1014–1022. [Google Scholar]
- Ikeda, H.; Kotaki, H.; Omura, S. Genetic studies of avermectin biosynthesis in Streptomyces avermitilis. J. Bacteriol. 1987, 169, 5615–5621. [Google Scholar]
- Shevchuk, N.A.; Bryksin, A.V.; Nusinovich, Y.A.; Cabello, F.C.; Sutherland, M.; Ladisch, S. Construction of long DNA molecules using long PCR-based fusion of several fragments simultaneously. Nucleic Acids Res. 2004, 32, e19. [Google Scholar] [CrossRef]
- Sioud, S.; Aigle, B.; Karray-Rebai, I.; Smaoui, S.; Bejar, S.; Mellouli, L. Integrative gene cloning and expression system for Streptomyces sp. Us 24 and Streptomyces sp. Tn 58 bioactive molecule producing strains. Biol. Med. Res. Int. 2009, 2009, 464986:1–464986:10. [Google Scholar]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Sun, F.-H.; Luo, D.; Shu, D.; Zhong, J.; Tan, H. Development of an Intergeneric Conjugal Transfer System for Xinaomycins-Producing Streptomyces noursei Xinao-4. Int. J. Mol. Sci. 2014, 15, 12217-12230. https://doi.org/10.3390/ijms150712217
Sun F-H, Luo D, Shu D, Zhong J, Tan H. Development of an Intergeneric Conjugal Transfer System for Xinaomycins-Producing Streptomyces noursei Xinao-4. International Journal of Molecular Sciences. 2014; 15(7):12217-12230. https://doi.org/10.3390/ijms150712217
Chicago/Turabian StyleSun, Feng-Hui, Di Luo, Dan Shu, Juan Zhong, and Hong Tan. 2014. "Development of an Intergeneric Conjugal Transfer System for Xinaomycins-Producing Streptomyces noursei Xinao-4" International Journal of Molecular Sciences 15, no. 7: 12217-12230. https://doi.org/10.3390/ijms150712217
APA StyleSun, F. -H., Luo, D., Shu, D., Zhong, J., & Tan, H. (2014). Development of an Intergeneric Conjugal Transfer System for Xinaomycins-Producing Streptomyces noursei Xinao-4. International Journal of Molecular Sciences, 15(7), 12217-12230. https://doi.org/10.3390/ijms150712217