S-(−)-10,11-Dihydroxyfarnesoic Acid Methyl Ester Inhibits Melanin Synthesis in Murine Melanocyte Cells



Abstract
:1. Introduction
2. Results and Discussion
2.1. Isolation, Purification, and Structural Determination of dhFAME


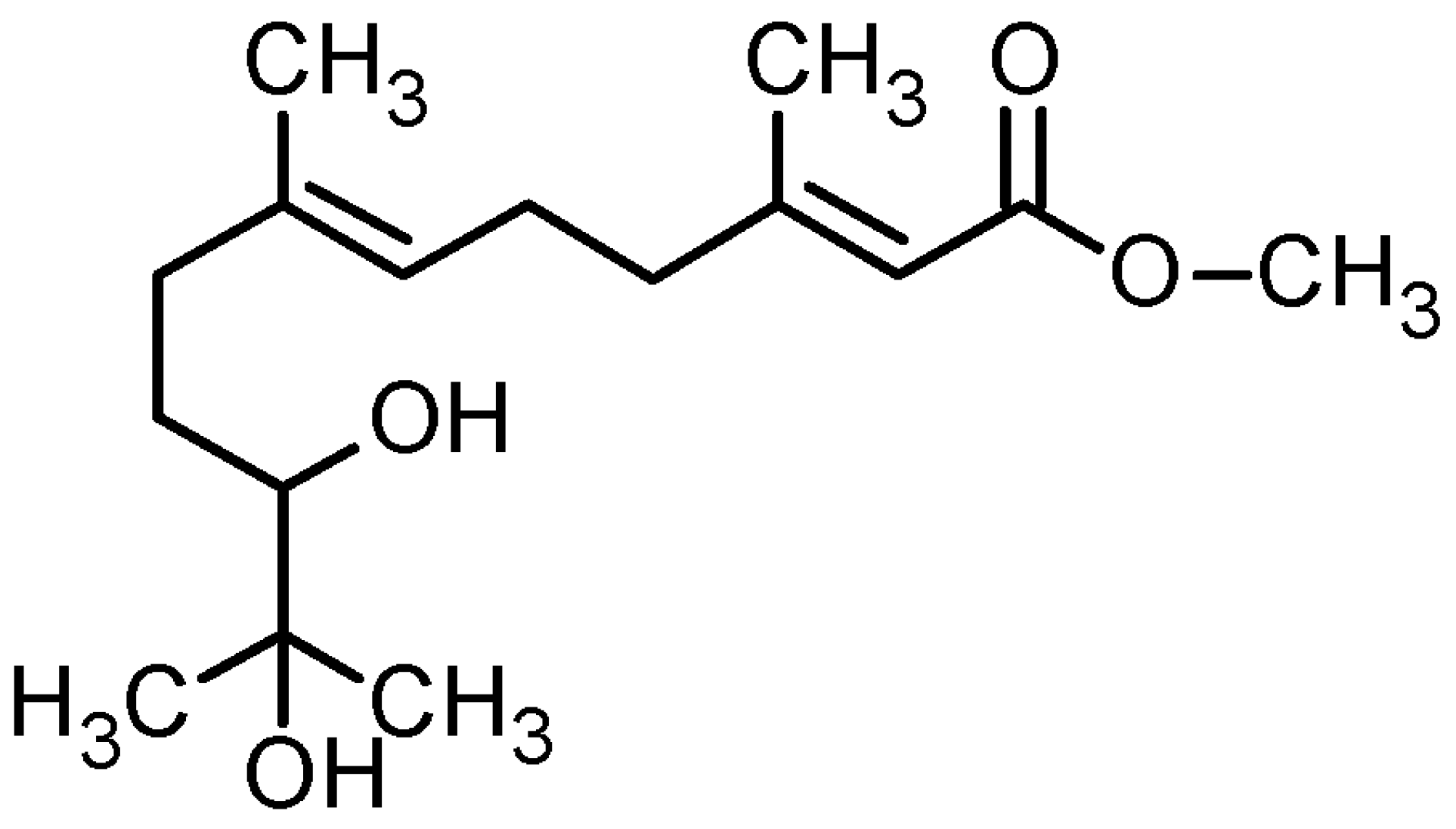{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Position | δH (ppm a) Multi ( J in Hz) | δC (ppm a) | ||
|---|---|---|---|---|
| 1 | 167.0 | C | ||
| 2 | 5.70 | br s | 115.2 | CH |
| 3 | 160.3 | C | ||
| 4 | 2.24 | m | 40.4 | CH2 |
| 5 | 2.24 | m | 25.7 | CH2 |
| 6 | 5.17 | br t (6.5) | 123.3 | CH |
| 7 | 136.3 | C | ||
| 8 | 2.25 | m | 36.6 | CH2 |
| 2.02 | m | |||
| 9 | 1.61 | m | 29.9 | CH2 |
| 1.30 | m | |||
| 10 | 3.20 | br ddd | 77.6 | CH2 |
| 11 | 72.3 | C | ||
| 12 b | 1.11 | s | 25.2 | CH3 |
| 13 b | 1.09 | s | 23.7 | CH3 |
| 14 | 1.64 | br s | 15.3 | CH3 |
| 15 | 2.17 | br d (1.0) | 18.0 | CH3 |
| OCH3 | 3.65 | s | 50.4 | CH3 |
| 10-OH | 2.82 | br d (5.0) | ||
| 11-OH | 2.66 | br s | ||
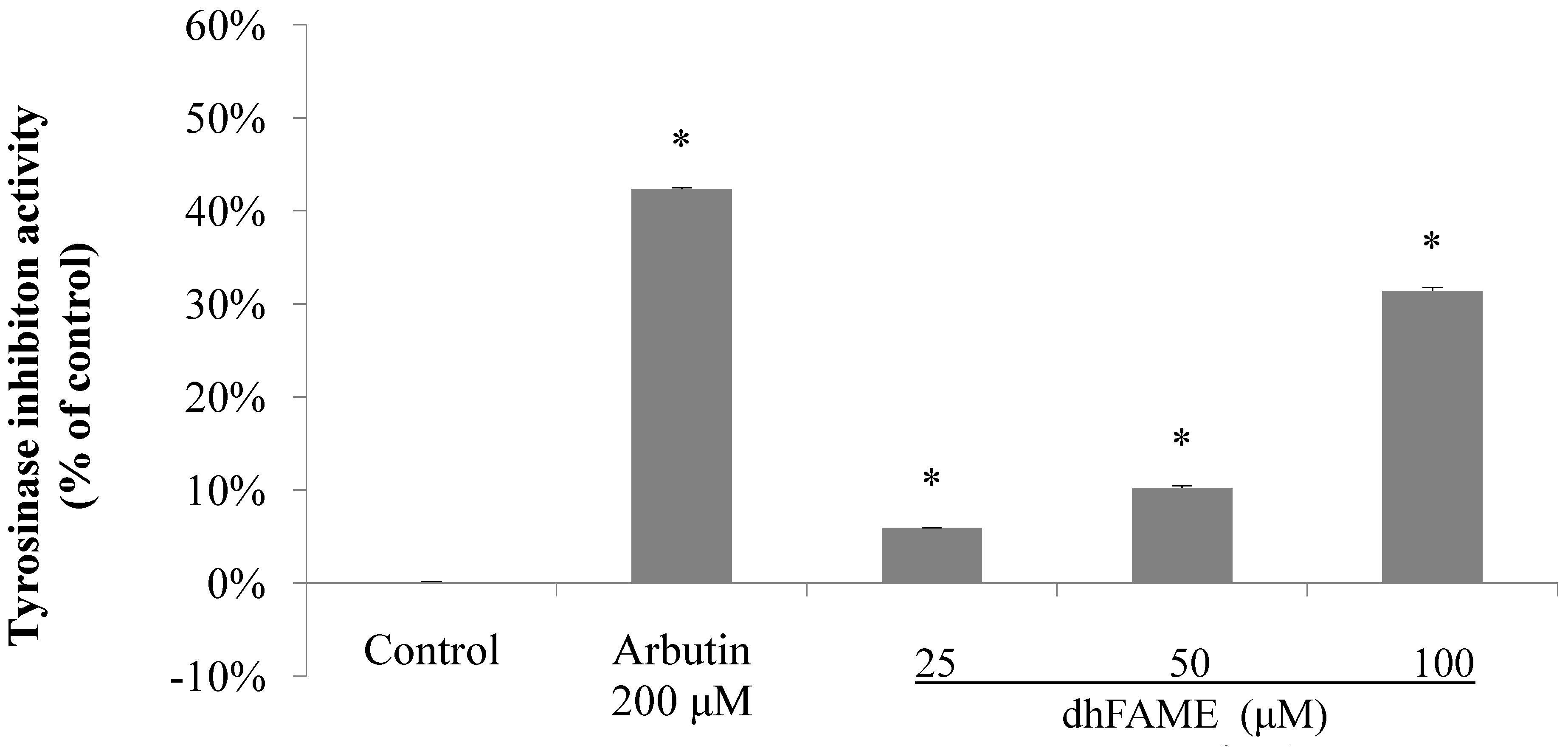
2.2. Effects of S-(−)-10,11-Dihydroxyfarnesoic Acid Methyl Ester (dhFAME) on Melanin Biosynthesis in Streptomyces Bikiniensis and on Tyrosinase Activity

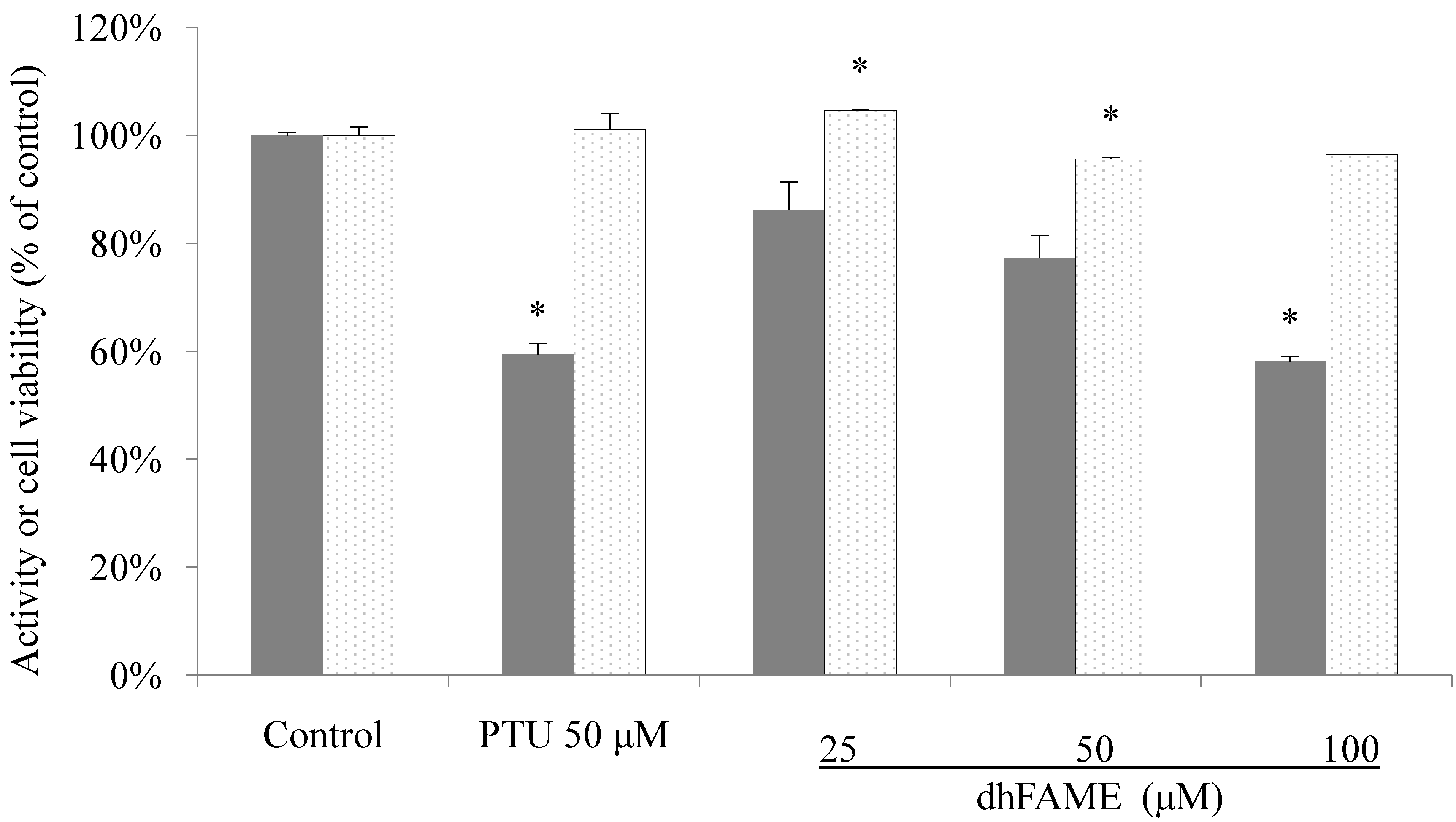
2.3. Effect of dhFAME on Cell Viability and Melanin Content

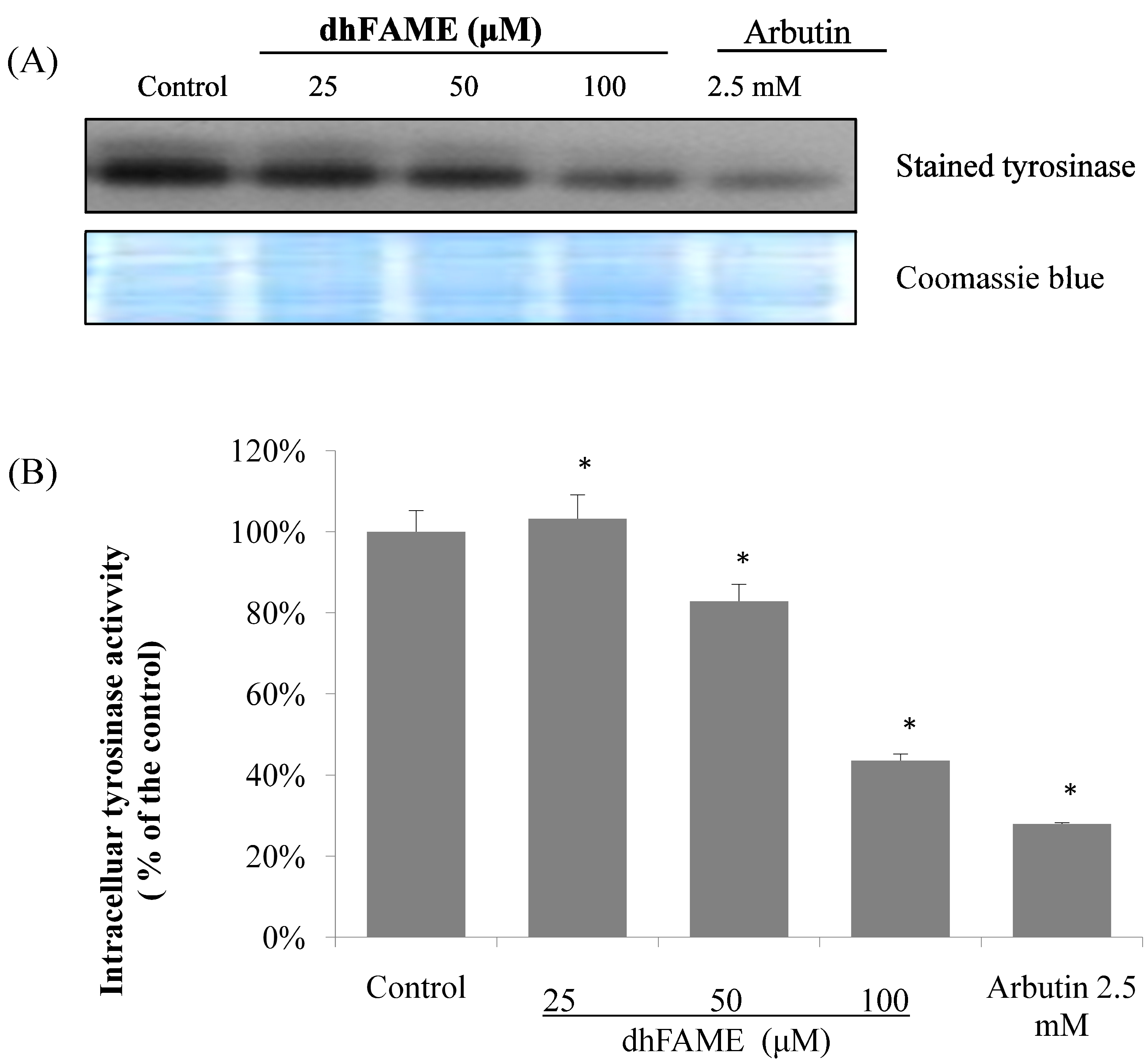
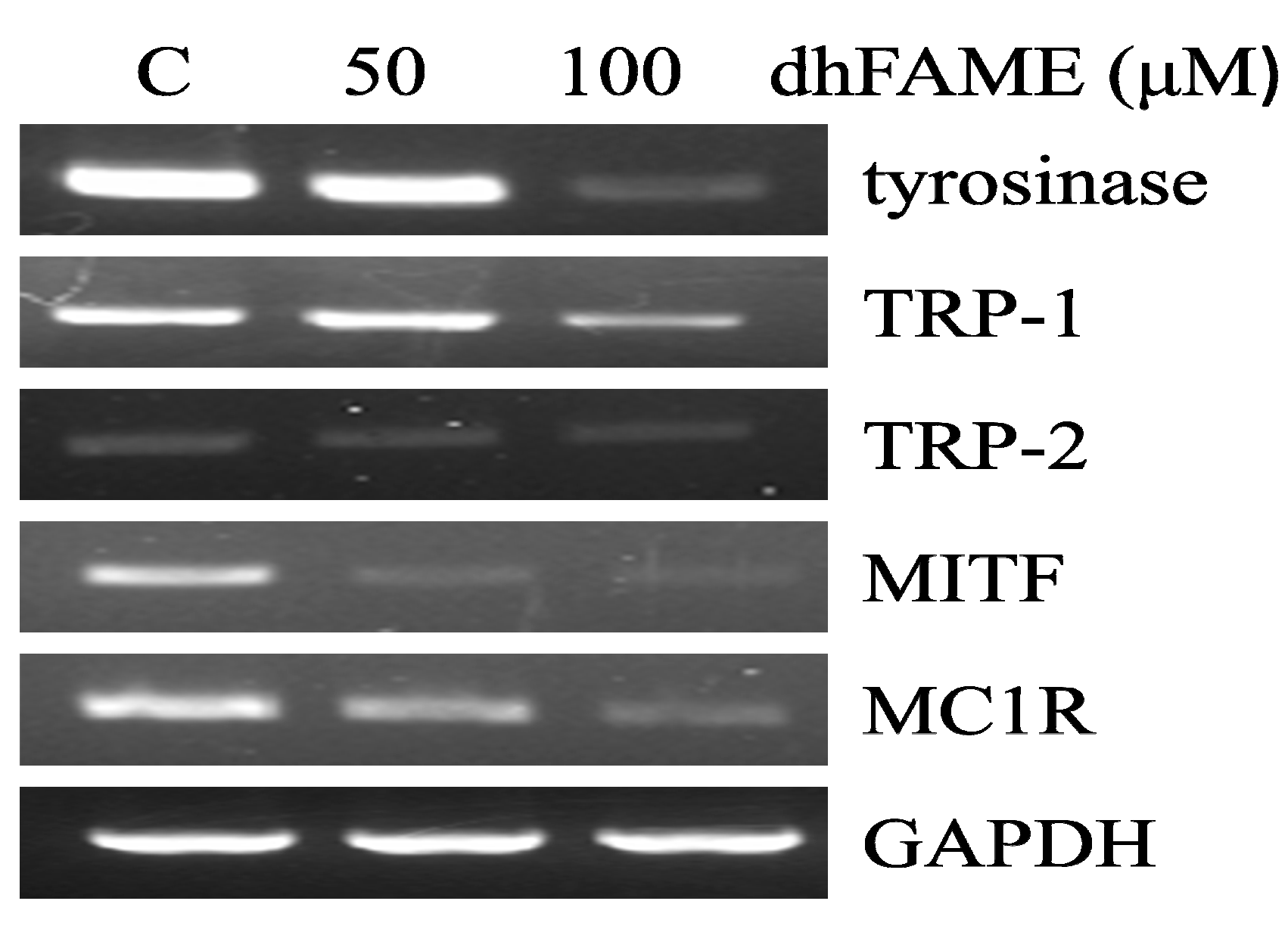
2.4. Effects of dhFAME on Zymography, Tyrosinase Assay and the Expression of Melanogenesis-Related Genes



3. Experimental Section
3.1. Materials
3.2. Purification and Structure Determination of S-(−)-10,11-Dihyroxyfarnesic Acid Methyl Ester (dhFAME)
3.3. Measurement of Tyrosinase Activity
3.4. Cell Culture
3.5. Cell Viability Assay
3.6. Melanization Inhibition Assay on Melan-a Cells
3.7. Analysis of Tyrosinase Activity by Zymography
3.8. Reverse-Transcription Polymerase Chain Reaction (RT-PCR) Analysis of mRNA Expression
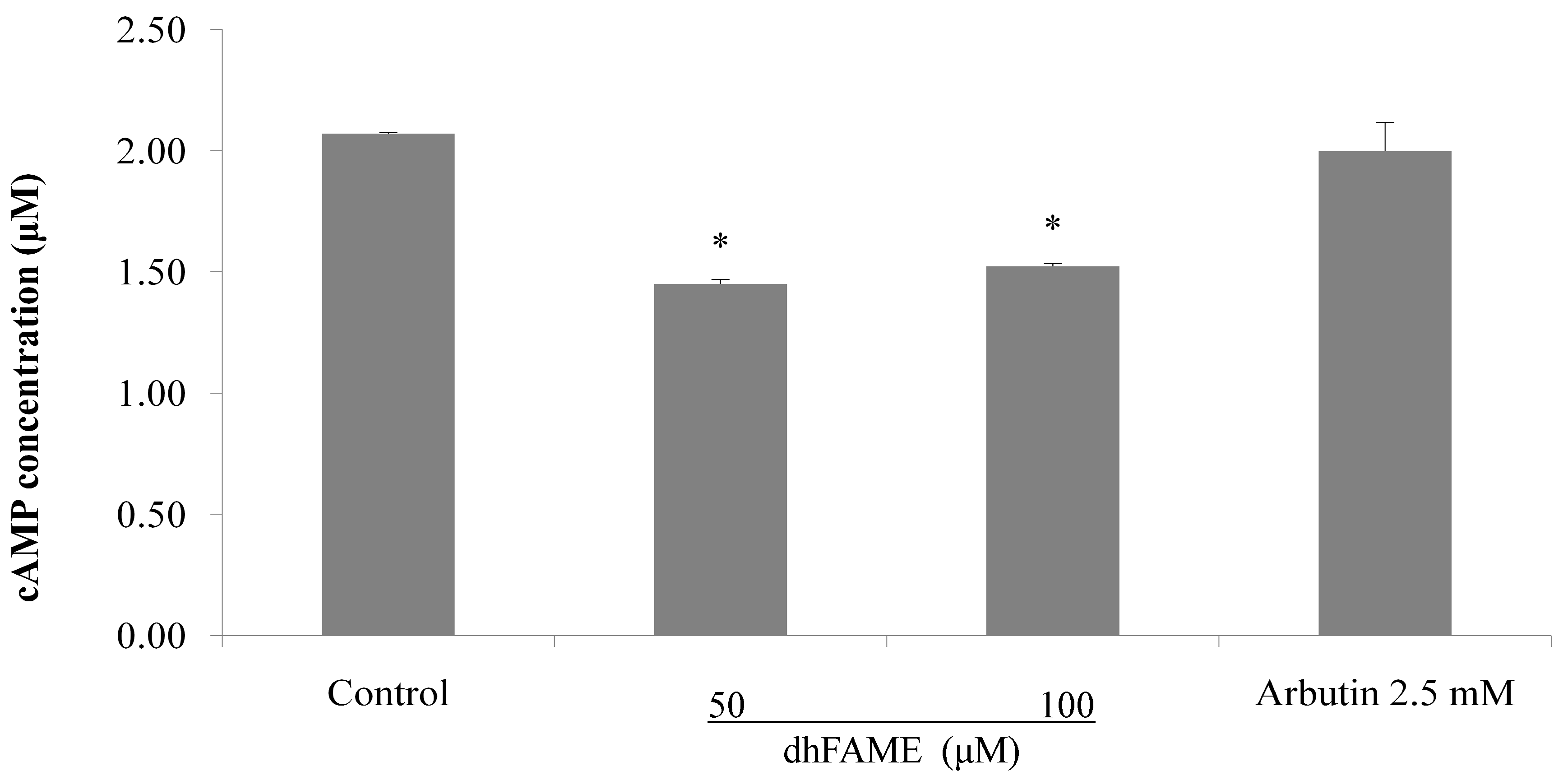
3.9. Measurement of cAMP
3.10. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Mårs, U.; Larsson, B.S. New thioureas and related substances intended for melanoma targeting. Pigment. Cell Res. 1995, 8, 194–201. [Google Scholar] [CrossRef]
- Smit, N.; Vicanova, J.; Pavel, S. The hunt for natural skin whitening agents. Int. J. Mol. Sci. 2009, 10, 5326–5349. [Google Scholar] [CrossRef]
- Ando, H.; Matsui, M.S.; Ichihashi, M. Quasi-drugs developed in Japan for the prevention or treatment of hyperpigmentary disorders. Int. J. Mol. Sci. 2010, 11, 2566–2575. [Google Scholar] [CrossRef]
- Callender, V.D.; St Surin-Lord, S.; Davis, E.C.; Maclin, M. Postinflammatory hyperpigmentation: Etiologic and therapeutic considerations. Am. J. Clin. Dermatol. 2011, 12, 87–99. [Google Scholar] [CrossRef]
- Hearing, V.J. Determination of melanin synthetic pathways. J. Investig. Dermatol. 2011, 131, E8–E11. [Google Scholar] [CrossRef]
- Melnikova, V.O.; Ananthaswamy, H.N. Cellular and molecular events leading to the development of skin cancer. Mutat. Res. 2005, 571, 91–106. [Google Scholar] [CrossRef]
- Nam, S.H.; Yoon, C.S.; Jeon, J.Y; Lee, S.H.; Lee, K.G.; Yeo, J.H.; Hwang, J.S. Mass Producing Method Of S-(−)-10,11-Dihydroxyfarnesoic Acid Methyl Ester For Suppressing Agent Of Melanogenesis. Korea Patent #10-1258699, 22 April 2013. [Google Scholar]
- Nam, S.H.; Yoon, C.S.; Jeon, J.Y.; Lee, S.H.; Lee, K.G.; Yeo, J.H.; Hwang, J.S. Composition for Suppressing Production of Melanogenesis. Korea Patent #10-1239631, 27 March 2013. [Google Scholar]
- Lamel, S.A.; Rahvar, M.; Maibach, H.I. Postinflammatory hyperpigmentation secondary to external insult: An overview of the quantitative analysis of pigmentation. Cutan. Ocul. Toxicol. 2013, 32, 67–71. [Google Scholar]
- Lin, J.Y.; Fisher, D.E. Melanocytebiology and skin pigmentation. Nature 2007, 445, 843–850. [Google Scholar] [CrossRef]
- Wu, Y.; Matsui, M.S.; Chen, J.Z.; Jin, X.; Shu, C.M.; Jin, G.Y.; Dong, G.H.; Wang, Y.K.; Gao, X.H.; Chen, H.D.; et al. Antioxidants add protection to a broad-spectrum sunscreen. Clin. Exp. Dermatol. 2011, 36, 178–187. [Google Scholar] [CrossRef]
- Sláma, K. Insect juvenile hormone analogues. Annu. Rev. Biochem. 2011, 40, 1079–1102. [Google Scholar] [CrossRef]
- Khalil, S.M.; Anspaugh, D.D.; Michael, R.R. Role of juvenile hormone esterase and epoxide hydrolase in reproduction of the cotton bollworm, Helicoverpa zea. J. Insect. Physiol. 2006, 52, 669–678. [Google Scholar]
- Mishima, Y.; Ohyama, Y.; Shibata, Y.; Seto, H.; Hatae, S. Inhibitory action of kojic acid on melanogenesis and its therapeutic effect for various human hyper-pigmentation disorders. Skin Res. 1994, 36, 134–150. [Google Scholar]
- Takizawa, T.; Mitsumori, K.; Tamura, T.; Nasu, M.; Ueda, M.; Imai, T.; Hirose, M. Hepatocellular tumor induction in heterozygous p53-deficient CBA mice by a 26-week dietary administration of kojic acid. Toxicol. Sci. 2003, 73, 287–293. [Google Scholar] [CrossRef]
- Son, H.U.; Lee, S.H. Evaluation of eye irritation by S-(−)-10,11-dihydroxyfarnesic acid methyl ester secreted by Beauveria. bassiana CS1029. Exp. Ther. Med. 2013, 6, 909–912. [Google Scholar]
- Oh, M.J.; Hamid, M.A.; Ngadiran, S.; Seo, Y.K.; Sarmidi, M.R.; Park, C.S. Ficus deltoidea (Mas cotek) extract exerted anti-melanogenic activity by preventing tyrosinase activity in vitro and by suppressing tyrosinase gene expression in B16F1 melanoma cells. Arch. Dermatol. Res. 2011, 303, 161–170. [Google Scholar] [CrossRef] [Green Version]
- Image J Software 1.48 version. Available online: http://imagej.nih.gov/ij/download.html (accessed on 31 January 2014).
- Videira, I.F.; Moura, D.F.; Magina, S. Mechanisms regulating melanogenesis. Bras. Dermatol. 2013, 88, 76–83. [Google Scholar] [CrossRef]
- Robinson, S.; Dixon, S.; August, S.; Diffey, B.; Wakamatsu, K.; Ito, S.; Friedmann, P.S.; Healy, E. Protection against UVR involves MC1R-mediated non-pigmentary and pigmentary mechanisms in vivo. J. Investig. Dermatol. 2010, 130, 1904–1913. [Google Scholar]
- Gunn, T.M.; Silvius, D.; Bagher, P.; Sun, K.; Walker, K.K. MGRN1-dependent pigment-type switching requires its ubiquitination activity but not its interaction with TSG101 or NEDD4. Pigment. Cell Melanoma Res. 2013, 26, 263–268. [Google Scholar] [CrossRef]
- Lee, A.Y.; Noh, M.S. The regulation of epidermal melanogenesis via cAMP and/or PKC signaling pathways: Insights for the development. Arch. Pharm. Res. 2013, 36, 792–801. [Google Scholar] [CrossRef]
- Park, H.Y.; Lee, J.; Gonzalez, S.; Middelkamp-Hup, M.A.; Kapasi, S.; Peterson, S.; Gilchrest, B.A. Topical application of a protein kinase C inhibitor reduces skin and hair pigmentation. J. Investig. Dermatol. 2004, 122, 159–166. [Google Scholar] [CrossRef]
- Lee, C.S.; Jang, W.H.; Park, M.; Jung, K.; Baek, H.S.; Joo, Y.H.; Park, Y.H.; Lim, K.M. A novel adamantyl benzylbenzamide derivative, AP736, suppresses melanogenesis through the inhibition of cAMP-PKA-CREB-activated microphthalmia-associated transcription factor and tyrosinase expression. Exp. Dermatol. 2013, 22, 762–764. [Google Scholar] [CrossRef]
- Guo, H.; Yang, K.; Deng, F.; Xing, Y.; Li, Y.; Lian, X.; Yang, T. Wnt3a inhibits proliferation but promotes melanogenesis of Melan-a cells. Int. J. Mol. Med. 2012, 30, 636–642. [Google Scholar]
- Ahn, S.J.; Koketsu, M.; Ishihara, H.; Lee, S.M.; Ha, S.K.; Lee, K.H.; Kang, T.H.; Kima, S.Y. Regulation of melanin synthesis by selenium-containing carbohydrates. Chem. Pharm. Bull. 2006, 54, 281–286. [Google Scholar]
- Elias, P.M.; Menon, G.; Wetzel, B.K.; Williams, J.W. Evidence that stress to the epidermal barrier influenced the development of pigmentation in humans. Pigment. Cell Melanoma Res. 2009, 22, 420–434. [Google Scholar] [CrossRef]
- Sato, K.; Takahashi, H.; Iraha, R.; Toriyama, M. Down-regulation of tyrosinase expression by acetylsalicylic acid in murine B16 melanoma. Biol. Pharm. Bull. 2008, 31, 33–37. [Google Scholar] [CrossRef]
- Ishihara, Y.; Oka, M.; Tsunakawa, M.; Tomita, K.; Hatori, M.; Yamamoto, H.; Kamei, H.; Miyaki, T.; Konishi, M.; Oki, T. Melanostatin, a new melanin synthesis inhibitor. Production, isolation, chemical properties, structure and biological activity. J. Antibiot. 1991, 44, 25–32. [Google Scholar]
- Goh, M.J.; Lee, H.K.; Cheng, L.; Kong, D.Y.; Yeon, J.H.; He, Q.Q.; Cho, J.C.; Na, Y.J. Depigmentation effect of kadsuralignan F on Melan-a murine melanocytes and human skin equivalents. Int. J. Mol. Sci. 2013, 14, 1655–1666. [Google Scholar]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Baek, S.-H.; Ahn, J.-W.; Nam, S.-H.; Yoon, C.-S.; Shin, J.-C.; Lee, S.-H. S-(−)-10,11-Dihydroxyfarnesoic Acid Methyl Ester Inhibits Melanin Synthesis in Murine Melanocyte Cells. Int. J. Mol. Sci. 2014, 15, 12750-12763. https://doi.org/10.3390/ijms150712750
Baek S-H, Ahn J-W, Nam S-H, Yoon C-S, Shin J-C, Lee S-H. S-(−)-10,11-Dihydroxyfarnesoic Acid Methyl Ester Inhibits Melanin Synthesis in Murine Melanocyte Cells. International Journal of Molecular Sciences. 2014; 15(7):12750-12763. https://doi.org/10.3390/ijms150712750
Chicago/Turabian StyleBaek, Seung-Hwa, Jun-Won Ahn, Sung-Hee Nam, Cheol-Sik Yoon, Jae-Cheon Shin, and Sang-Han Lee. 2014. "S-(−)-10,11-Dihydroxyfarnesoic Acid Methyl Ester Inhibits Melanin Synthesis in Murine Melanocyte Cells" International Journal of Molecular Sciences 15, no. 7: 12750-12763. https://doi.org/10.3390/ijms150712750
APA StyleBaek, S.-H., Ahn, J.-W., Nam, S.-H., Yoon, C.-S., Shin, J.-C., & Lee, S.-H. (2014). S-(−)-10,11-Dihydroxyfarnesoic Acid Methyl Ester Inhibits Melanin Synthesis in Murine Melanocyte Cells. International Journal of Molecular Sciences, 15(7), 12750-12763. https://doi.org/10.3390/ijms150712750