Localization of MLH3 at the Centrosomes

{kind=link}
{kind=link}
Abstract
:1. Introduction
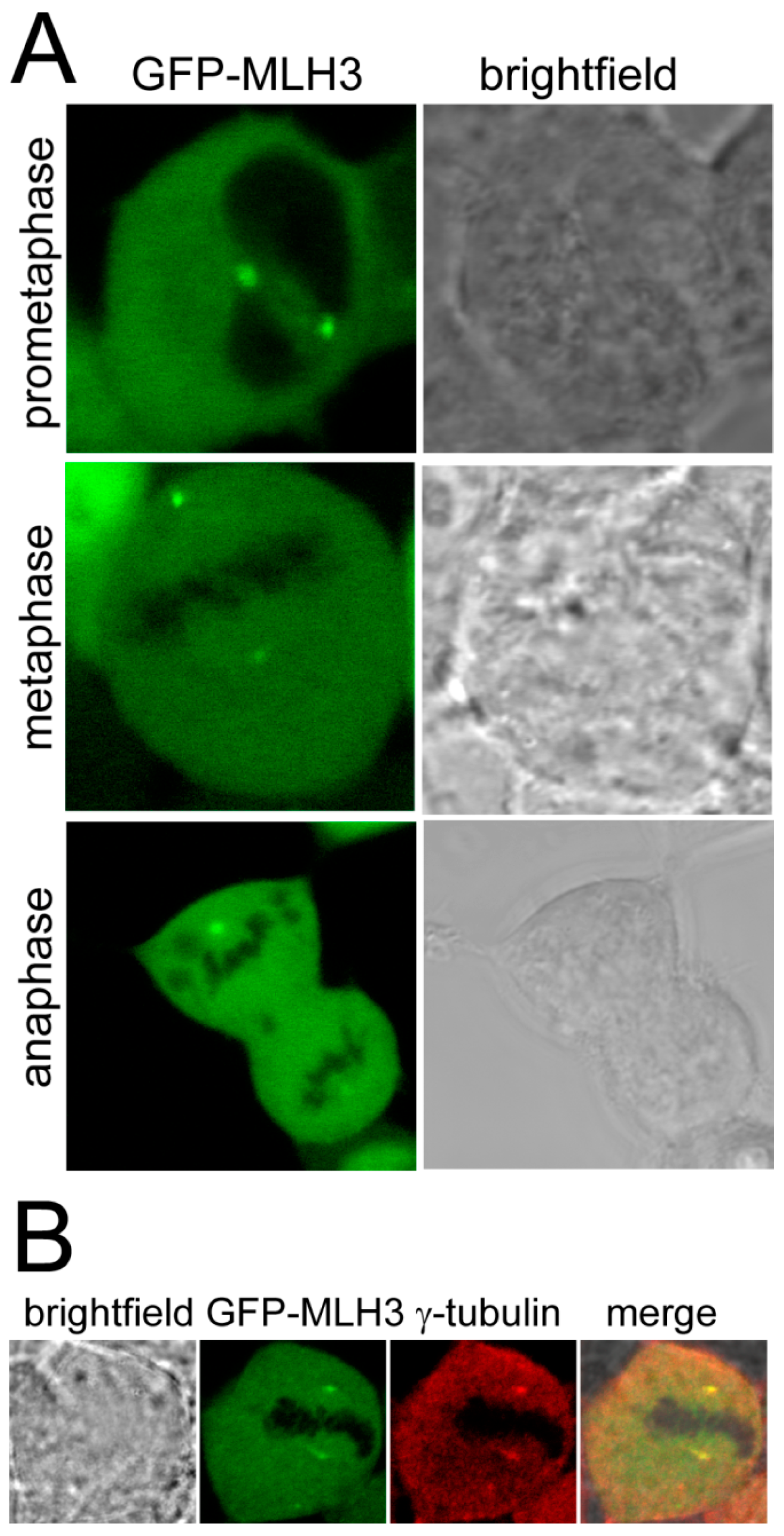
2. Results and Discussion


3. Experimental Section
3.1. Cell Lines
3.2. Immunofluorescence Staining
3.3. Microscopy
3.4. Fluorescence Recovery after Photobleaching (FRAP)
4. Conclusions
Author Contributions
Conflicts of Interest
References
- Cohen, P.E.; Pollack, S.E.; Pollard, J.W. Genetic analysis of chromosome pairing, recombination, and cell cycle control during first meiotic prophase in mammals. Endocr. Rev. 2006, 27, 398–426. [Google Scholar] [CrossRef]
- Kolas, N.K.; Svetlanov, A.; Lenzi, M.L.; Macaluso, F.P.; Lipkin, S.M.; Liskay, R.M.; Greally, J.; Edelmann, W.; Cohen, P.E. Localization of MMR proteins on meiotic chromosomes in mice indicates distinct functions during prophase I. J. Cell. Biol. 2005, 171, 447–458. [Google Scholar] [CrossRef]
- Lipkin, S.M.; Moens, P.B.; Wang, V.; Lenzi, M.; Shanmugarajah, D.; Gilgeous, A.; Thomas, J.; Cheng, J.; Touchman, J.W.; Green, E.D.; et al. Meiotic arrest and aneuploidy in MLH3-deficient mice. Nat. Genet. 2002, 31, 385–390. [Google Scholar]
- Santucci-Darmanin, S.; Neyton, S.; Lespinasse, F.; Saunieres, A.; Gaudray, P.; Paquis-Flucklinger, V. The DNA mismatch-repair MLH3 protein interacts with MSH4 in meiotic cells, supporting a role for this MutL homolog in mammalian meiotic recombination. Hum. Mol. Genet. 2002, 11, 1697–1706. [Google Scholar] [CrossRef]
- Charbonneau, N.; Amunugama, R.; Schmutte, C.; Yoder, K.; Fishel, R. Evidence that hMLH3 functions primarily in meiosis and in hMSH2–hMSH3 mismatch repair. Cancer Biol. Ther. 2009, 8, 1411–1420. [Google Scholar] [CrossRef]
- Lipkin, S.M.; Wang, V.; Jacoby, R.; Banerjee-Basu, S.; Baxevanis, A.D.; Lynch, H.T.; Elliott, R.M.; Collins, F.S. MLH3: A DNA mismatch repair gene associated with mammalian microsatellite instability. Nat. Genet. 2000, 24, 27–35. [Google Scholar] [CrossRef]
- Roesner, L.M.; Mielke, C.; Fahnrich, S.; Merkhoffer, Y.; Dittmar, K.E.; Drexler, H.G.; Dirks, W.G. Stable expression of MutLγ in human cells reveals no specific response to mismatched DNA, but distinct recruitment to damage sites. J. Cell. Biochem. 2013, 114, 2405–2414. [Google Scholar] [CrossRef]
- Mortusewicz, O.; Leonhardt, H. XRCC1 and PCNA are loading platforms with distinct kinetic properties and different capacities to respond to multiple DNA lesions. BMC Mol. Biol. 2007, 8, 81. [Google Scholar] [CrossRef]
- Mortusewicz, O.; Rothbauer, U.; Cardoso, M.C.; Leonhardt, H. Differential recruitment of DNA Ligase I and III to DNA repair sites. Nucleic Acids Res. 2006, 34, 3523–3532. [Google Scholar] [CrossRef]
- Nigg, E.A. Centrosome aberrations: Cause or consequence of cancer progression? Nat. Rev. Cancer 2002, 2, 815–825. [Google Scholar] [CrossRef]
- Kwon, M.; Godinho, S.A.; Chandhok, N.S.; Ganem, N.J.; Azioune, A.; Thery, M.; Pellman, D. Mechanisms to suppress multipolar divisions in cancer cells with extra centrosomes. Genes Dev. 2008, 22, 2189–2203. [Google Scholar] [CrossRef]
- Shimada, M.; Komatsu, K. Emerging connection between centrosome and DNA repair machinery. J. Radiat. Res. 2009, 50, 295–301. [Google Scholar] [CrossRef]
- Campbell, M.R.; Wang, Y.; Andrew, S.E.; Liu, Y. MSH2 deficiency leads to chromosomal abnormalities, centrosome amplification, and telomere capping defect. Oncogene 2006, 25, 2531–2536. [Google Scholar] [CrossRef]
- Narine, K.A.; Keuling, A.M.; Gombos, R.; Tron, V.A.; Andrew, S.E.; Young, L.C. Defining the DNA mismatch repair-dependent apoptotic pathway in primary cells: Evidence for p53-independence and involvement of centrosomal caspase 2. DNA Repair 2010, 9, 161–168. [Google Scholar] [CrossRef]
- Geissler, S.; Siegers, K.; Schiebel, E. A novel protein complex promoting formation of functional α- and γ-tubulin. EMBO J. 1998, 17, 952–966. [Google Scholar] [CrossRef]
- Her, C.; Wu, X.; Griswold, M.D.; Zhou, F. Human MutS homologue MSH4 physically interacts with von Hippel–Lindau tumor suppressor-binding protein 1. Cancer Res. 2003, 63, 865–872. [Google Scholar]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Roesner, L.M.; Mielke, C.; Faehnrich, S.; Merkhoffer, Y.; Dittmar, K.E.J.; Drexler, H.G.; Dirks, W.G. Localization of MLH3 at the Centrosomes. Int. J. Mol. Sci. 2014, 15, 13932-13937. https://doi.org/10.3390/ijms150813932
Roesner LM, Mielke C, Faehnrich S, Merkhoffer Y, Dittmar KEJ, Drexler HG, Dirks WG. Localization of MLH3 at the Centrosomes. International Journal of Molecular Sciences. 2014; 15(8):13932-13937. https://doi.org/10.3390/ijms150813932
Chicago/Turabian StyleRoesner, Lennart M., Christian Mielke, Silke Faehnrich, Yvonne Merkhoffer, Kurt E. J. Dittmar, Hans G. Drexler, and Wilhelm G. Dirks. 2014. "Localization of MLH3 at the Centrosomes" International Journal of Molecular Sciences 15, no. 8: 13932-13937. https://doi.org/10.3390/ijms150813932
APA StyleRoesner, L. M., Mielke, C., Faehnrich, S., Merkhoffer, Y., Dittmar, K. E. J., Drexler, H. G., & Dirks, W. G. (2014). Localization of MLH3 at the Centrosomes. International Journal of Molecular Sciences, 15(8), 13932-13937. https://doi.org/10.3390/ijms150813932