
Increased ARPP-19 Expression Is Associated with Hepatocellular Carcinoma

Abstract
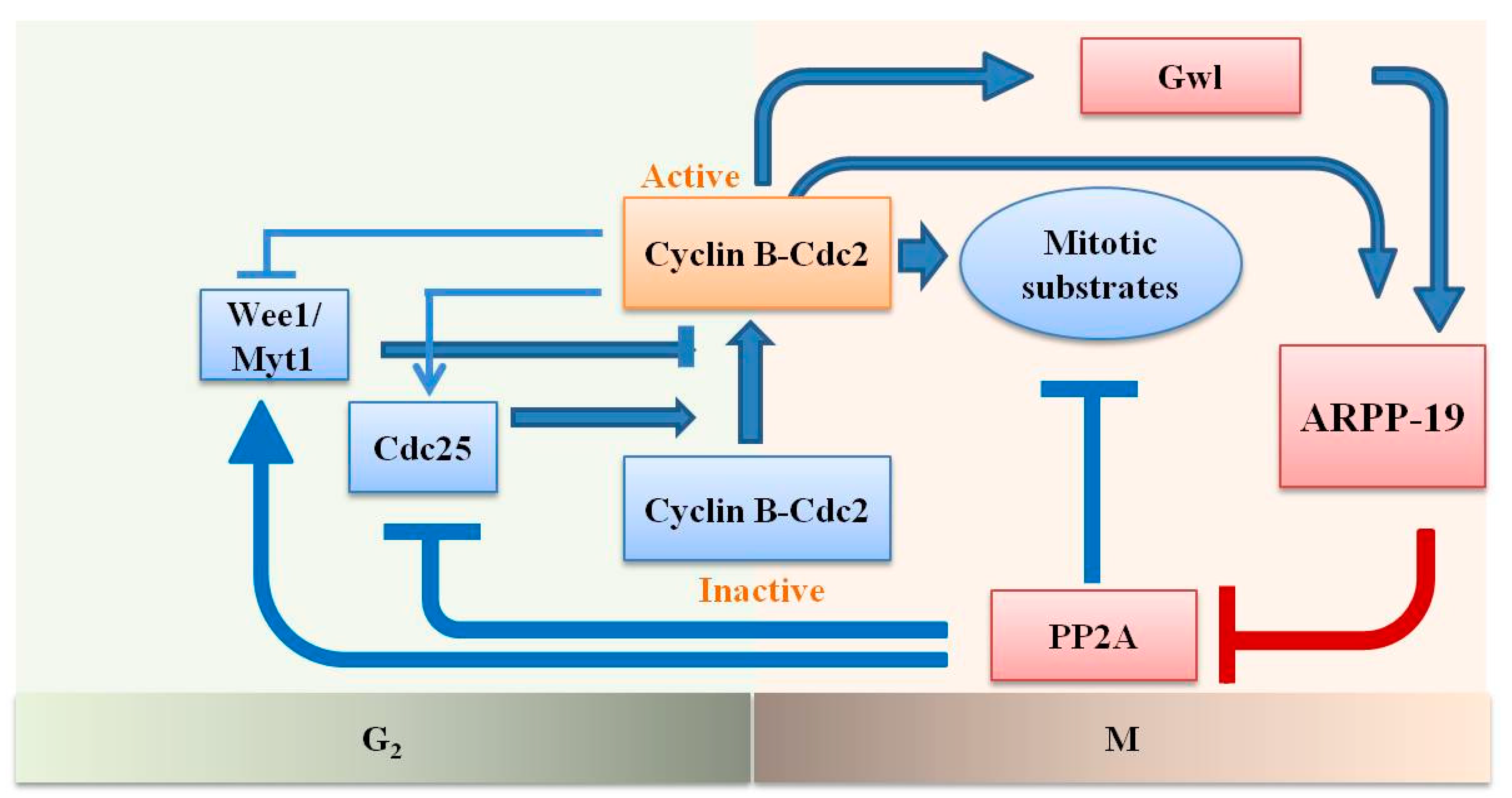
:1. Introduction

2. Results
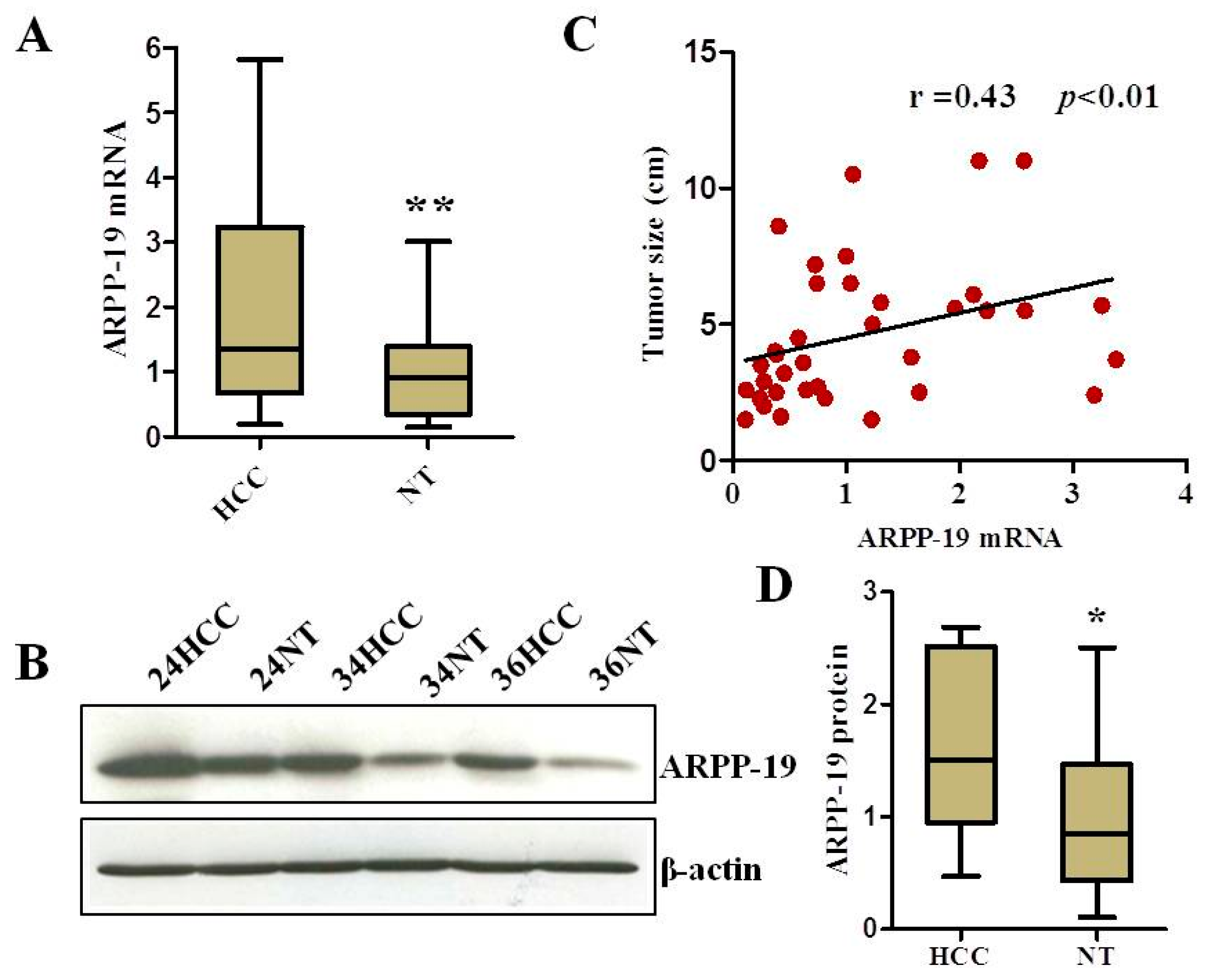
2.1. ARPP-19 Expression in HCC and Corresponding Non-Tumorous Liver Tissues


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Category | Number | ARPP-19 Median (Range) | p-Value |
|---|---|---|---|---|
| Age (years) | ≤50 | 16 | 0.77 (0.12, 2.2) | 0.23 |
| >50 | 30 | 0.91 (0.11, 3.4) | ||
| Gender | Male | 31 | 1.0 (0.24, 3.4) | 0.08 |
| Female | 5 | 0.28 (0.11, 2.1) | ||
| Etiology | HBV | 30 | 0.74 (0.11, 3.3) | 0.17 |
| HCV | 2 | 0.99 (0.74, 1.2) | ||
| Unknown | 4 | 2.1 (0.28, 3.4) | ||
| Histologic grade | I–II | 17 | 0.58 (0.11, 2.0) | 0.06 |
| III–IV | 19 | 1.0 (0.12, 3.4) | ||
| Tumor size | ≤5 cm | 22 | 0.52 (0.11, 3.4) | 0.009 |
| >5 cm | 14 | 1.6 (0.41, 3.3) | ||
| Liver Cirrhosis | Absent | 5 | 0.74 (0.25, 3.3) | 0.91 |
| Present | 31 | 0.82 (0.11, 3.4) | ||
| Serum AFP | ≤20 ng/mL | 16 | 0.7 (0.11, 3.3) | 0.18 |
| >20 ng/mL | 20 | 1.0 (0.12, 3.4) |
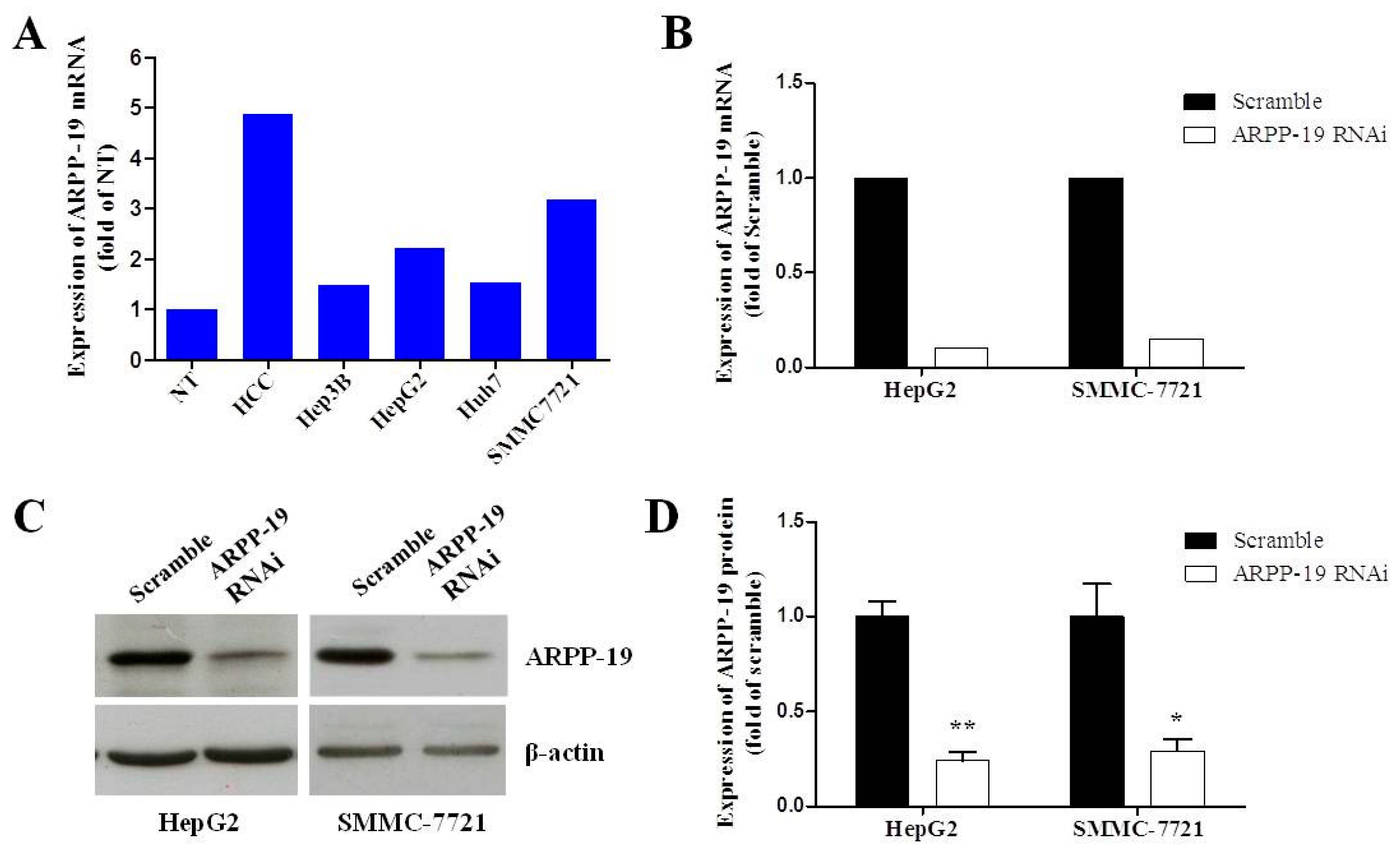
2.2. Silencing ARPP-19 Expression in Hepatocarcinoma Cells

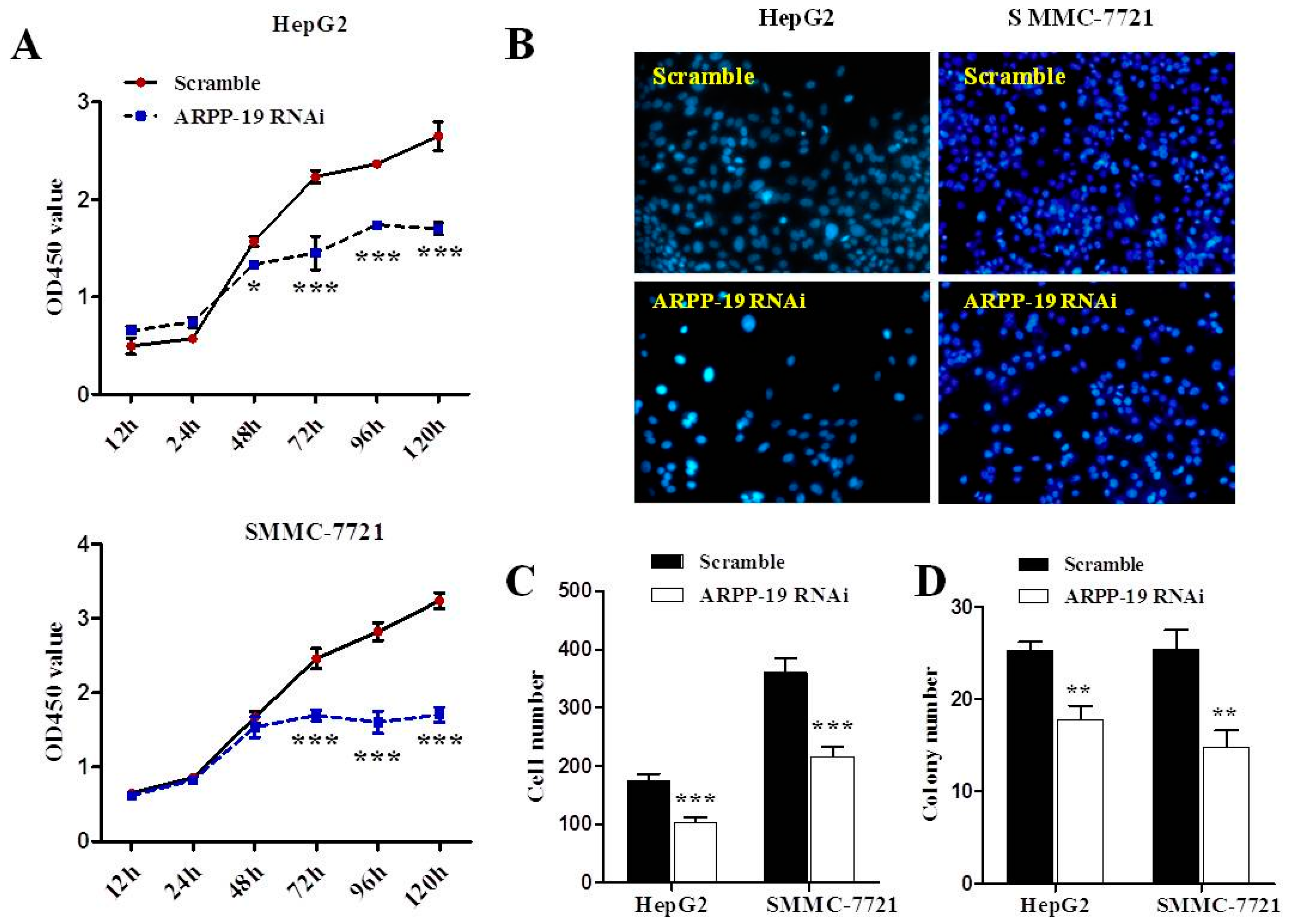
2.3. Down-Expression of ARPP-19 Inhibits Cell Proliferation

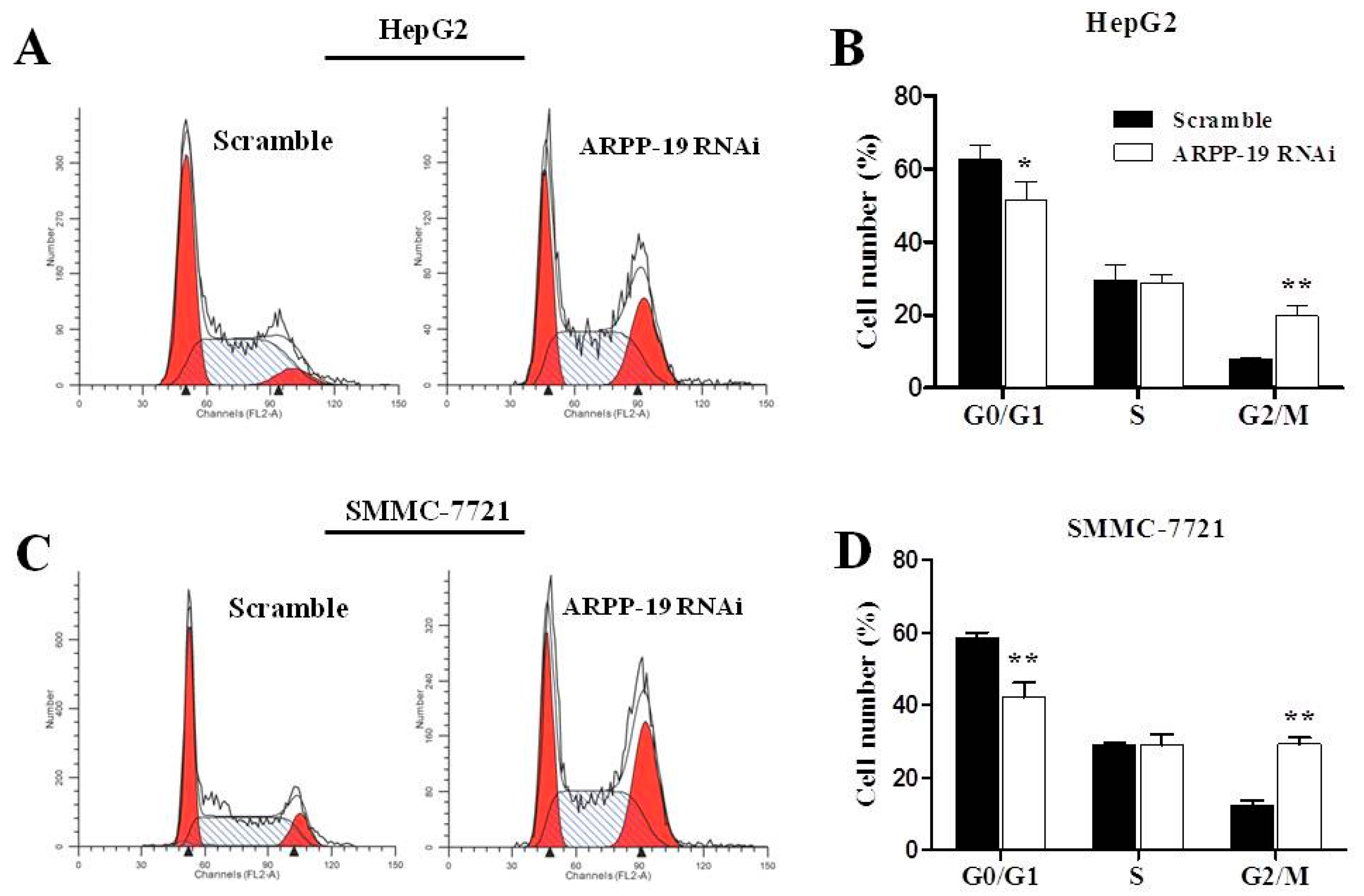
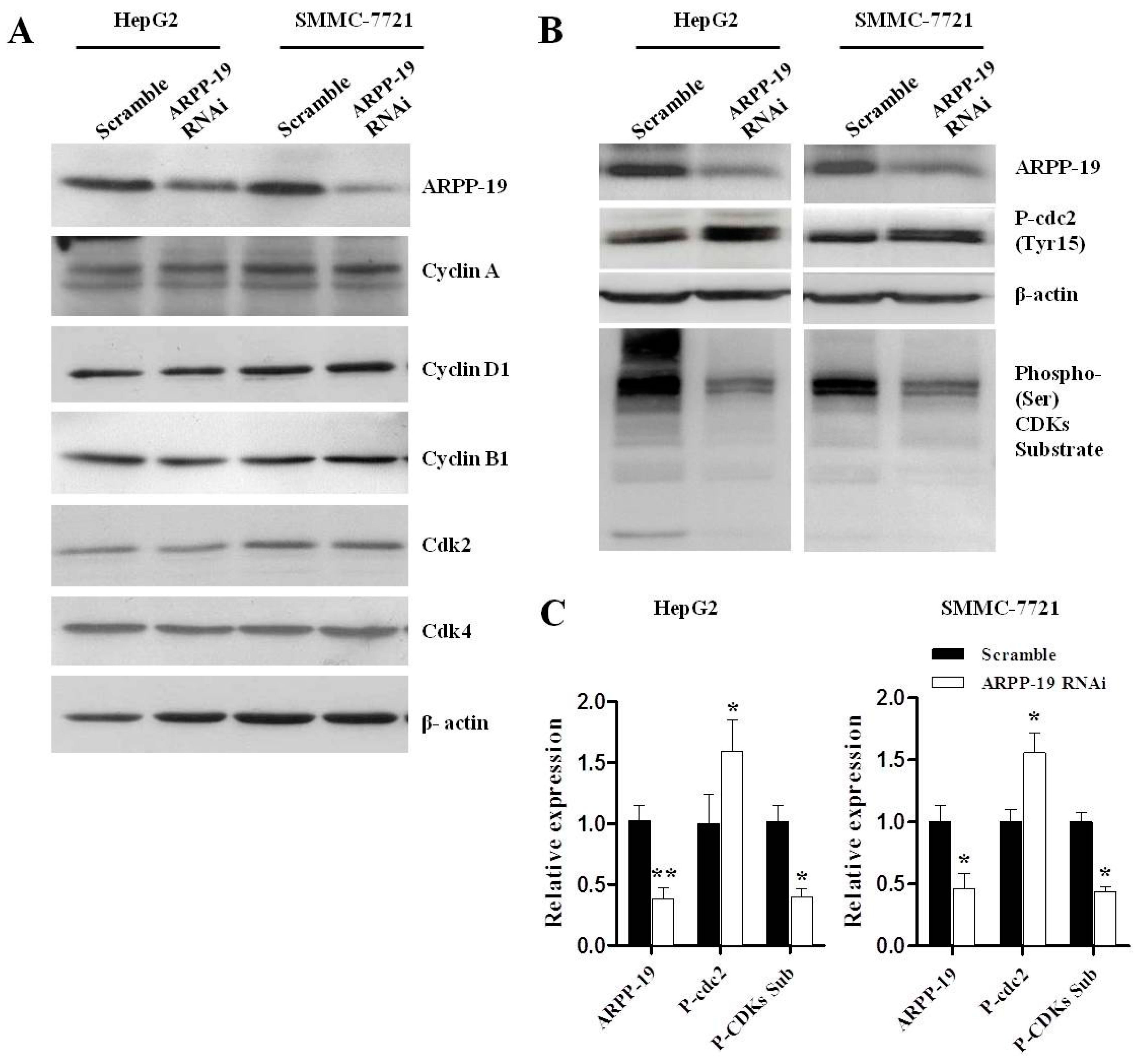
2.4. Down-Expression of ARPP-19 Affects Cell Cycle at G2/M Phase


3. Discussion
4. Materials and Methods
4.1. Tissue Specimen and Cell Lines
4.2. RNA Extraction and Quantitative Reverse Transcription-Polymerase Chain Reaction (qRT-PCR)
4.3. Western Blot Analysis
4.4. Knockdown of ARPP-19 in Hepatocarcinoma Cells
4.5. Cell Counting
4.6. Cell Counting Kit-8 Assay for Cell Proliferation
4.7. Clone Formation Assay
4.8. Cell Cycle Distribution Analysis
4.9. Cell Synchronization
4.10. Statistical Analyses
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Forner, A.; Llovet, J.M.; Bruix, J. Hepatocellular carcinoma. Lancet 2012, 379, 1245–1255. [Google Scholar]
- Shiraha, H.; Yamamoto, K.; Namba, M. Human hepatocyte carcinogenesis (review). Int. J. Oncol. 2013, 42, 1133–1138. [Google Scholar]
- Zhai, X.F.; Chen, Z.; Li, B.; Shen, F.; Fan, J.; Zhou, W.P.; Yang, Y.K.; Xu, J.; Qin, X.; Li, L.Q.; et al. Traditional herbal medicine in preventing recurrence after resection of small hepatocellular carcinoma: A multicenter randomized controlled trial. J. Integr. Med. 2013, 11, 90–100. [Google Scholar] [CrossRef] [PubMed]
- Villanueva, A.; Hernandez-Gea, V.; Llovet, J.M. Medical therapies for hepatocellular carcinoma: A critical view of the evidence. Nat. Rev. Gastroenterol. Hepatol. 2013, 10, 34–42. [Google Scholar]
- Marrero, J.A. Multidisciplinary management of hepatocellular carcinoma: Where are we today? Semin. Liver Dis. 2013, 33, S3–S10. [Google Scholar] [CrossRef] [PubMed]
- Horiuchi, A.; Williams, K.R.; Kurihara, T.; Nairn, A.C.; Greengard, P. Purification and cDNA cloning of ARPP-16, a cAMP-regulated phosphoprotein enriched in basal ganglia, and of a related phosphoprotein, ARPP-19. J. Biol. Chem. 1990, 265, 9476–9484. [Google Scholar]
- Dulubova, I.; Horiuchi, A.; Snyder, G.L.; Girault, J.A.; Czernik, A.J.; Shao, L.; Ramabhadran, R.; Greengard, P.; Nairn, A.C. ARPP-16/ARPP-19: A highly conserved family of cAMP-regulated phosphoproteins. J. Neurochem. 2001, 77, 229–238. [Google Scholar] [CrossRef] [PubMed]
- Girault, J.A.; Horiuchi, A.; Gustafson, E.L.; Rosen, N.L.; Greengard, P. Differential expression of ARPP-16 and ARPP-19, two highly related cAMP-regulated phosphoproteins, one of which is specifically associated with dopamine-innervated brain regions. J. Neurosci. 1990, 10, 1124–1133. [Google Scholar] [PubMed]
- Irwin, N.; Chao, S.; Goritchenko, L.; Horiuchi, A.; Greengard, P.; Nairn, A.C.; Benowitz, L.I. Nerve growth factor controls GAP-43 mRNA stability via the phosphoprotein ARPP-19. Proc. Natl. Acad. Sci. USA 2002, 99, 12427–12431. [Google Scholar] [PubMed]
- Kim, S.H.; Nairn, A.C.; Cairns, N.; Lubec, G. Decreased levels of ARPP-19 and PKA in brains of Down syndrome and Alzheimer’s disease. J. Neural. Transm. Suppl. 2001, 61, 263–272. [Google Scholar] [PubMed]
- Gharbi-Ayachi, A.; Labbe, J.C.; Burgess, A.; Vigneron, S.; Strub, J.M.; Brioudes, E.; van-Dorsselaer, A.; Castro, A.; Lorca, T. The substrate of Greatwall kinase, Arpp19, controls mitosis by inhibiting protein phosphatase 2A. Science 2010, 330, 1673–1677. [Google Scholar] [PubMed]
- Haccard, O.; Jessus, C. Greatwall kinase, ARPP-19 and protein phosphatase 2A: Shifting the mitosis paradigm. Results Probl. Cell Differ. 2011, 53, 219–234. [Google Scholar] [PubMed]
- Lorca, T.; Castro, A. The Greatwall kinase: A new pathway in the control of the cell cycle. Oncogene 2013, 32, 537–543. [Google Scholar] [CrossRef] [PubMed]
- Dupre, A.; Buffin, E.; Roustan, C.; Nairn, A.C.; Jessus, C.; Haccard, O. The phosphorylation of ARPP19 by Greatwall renders the auto-amplification of MPF independently of PKA in Xenopus oocytes. J. Cell Sci. 2013, 126, 3916–3926. [Google Scholar] [CrossRef] [PubMed]
- Hara, M.; Abe, Y.; Tanaka, T.; Yamamoto, T.; Okumura, E.; Kishimoto, T. Greatwall kinase and cyclin B-Cdk1 are both critical constituents of M-phase-promoting factor. Nat. Commun. 2012, 3, 1059. [Google Scholar] [CrossRef] [PubMed]
- Okumura, E.; Morita, A.; Wakai, M.; Mochida, S.; Hara, M.; Kishimoto, T. Cyclin B-Cdk1 inhibits protein phosphatase PP2A-B55 via a Greatwall kinase-independent mechanism. J. Cell Biol. 2014, 204, 881–889. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, G.K.; Shah, M.A. Targeting the cell cycle: A new approach to cancer therapy. J. Clin. Oncol. 2005, 23, 9408–9421. [Google Scholar] [CrossRef] [PubMed]
- Fu, L.Y.; Jia, H.L.; Dong, Q.Z.; Wu, J.C.; Zhao, Y.; Zhou, H.J.; Ren, N.; Ye, Q.H.; Qin, L.X. Suitable reference genes for real-time PCR in human HBV-related hepatocellular carcinoma with different clinical prognoses. BMC Cancer 2009, 9, 49. [Google Scholar] [CrossRef] [PubMed]
- Waxman, S.; Wurmbach, E. De-regulation of common housekeeping genes in hepatocellular carcinoma. BMC Genomics 2007, 8, 243. [Google Scholar] [CrossRef] [PubMed]
- Arzumanyan, A.; Reis, H.M.; Feitelson, M.A. Pathogenic mechanisms in HBV- and HCV-associated hepatocellular carcinoma. Nat. Rev. Cancer 2013, 13, 123–135. [Google Scholar] [CrossRef] [PubMed]
- Hunt, T. On the regulation of protein phosphatase 2A and its role in controlling entry into and exit from mitosis. Adv. Biol. Regul. 2013, 53, 173–178. [Google Scholar] [CrossRef] [PubMed]
- Burgess, A.; Vigneron, S.; Brioudes, E.; Labbe, J.C.; Lorca, T.; Castro, A. Loss of human Greatwall results in G2 arrest and multiple mitotic defects due to deregulation of the cyclin B-Cdc2/PP2A balance. Proc. Natl. Acad. Sci. USA 2010, 107, 12564–12569. [Google Scholar] [CrossRef] [PubMed]
- Voets, E.; Wolthuis, R.M. MASTL is the human orthologue of Greatwall kinase that facilitates mitotic entry, anaphase and cytokinesis. Cell Cycle 2010, 9, 3591–3601. [Google Scholar] [CrossRef] [PubMed]
- Mochida, S.; Hunt, T. Protein phosphatases and their regulation in the control of mitosis. EMBO Rep. 2012, 13, 197–203. [Google Scholar] [CrossRef] [PubMed]
- Lorca, T.; Bernis, C.; Vigneron, S.; Burgess, A.; Brioudes, E.; Labbe, J.C.; Castro, A. Constant regulation of both the MPF amplification loop and the Greatwall-PP2A pathway is required for metaphase II arrest and correct entry into the first embryonic cell cycle. J. Cell Sci. 2010, 123, 2281–2291. [Google Scholar] [CrossRef] [PubMed]
- Eichhorn, P.J.; Creyghton, M.P.; Bernards, R. Protein phosphatase 2A regulatory subunits and cancer. Biochim. Biophys. Acta 2009, 1795, 1–15. [Google Scholar] [PubMed]
- Girault, J.A.; Shalaby, I.A.; Rosen, N.L.; Greengard, P. Regulation by cAMP and vasoactive intestinal peptide of phosphorylation of specific proteins in striatal cells in culture. Proc. Natl. Acad. Sci. USA 1988, 85, 7790–7794. [Google Scholar] [CrossRef] [PubMed]
- Veljkovic, M.; Dopsaj, V.; Dopsaj, M.; Branch, D.R.; Veljkovic, N.; Sakarellos-Daitsiotis, M.M.; Veljkovic, V.; Glisic, S.; Colombatti, A. Physical activity and natural anti-VIP antibodies: Potential role in breast and prostate cancer therapy. PLoS One 2011, 6, e28304. [Google Scholar] [CrossRef] [PubMed]
- Sachlos, E.; Risueno, R.M.; Laronde, S.; Shapovalova, Z.; Lee, J.H.; Russell, J.; Malig, M.; McNicol, J.D.; Fiebig-Comyn, A.; Graham, M.; et al. Identification of drugs including a dopamine receptor antagonist that selectively target cancer stem cells. Cell 2012, 149, 1284–1297. [Google Scholar] [CrossRef] [PubMed]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Song, H.; Pan, J.; Liu, Y.; Wen, H.; Wang, L.; Cui, J.; Liu, Y.; Hu, B.; Yao, Z.; Ji, G. Increased ARPP-19 Expression Is Associated with Hepatocellular Carcinoma. Int. J. Mol. Sci. 2015, 16, 178-192. https://doi.org/10.3390/ijms16010178
Song H, Pan J, Liu Y, Wen H, Wang L, Cui J, Liu Y, Hu B, Yao Z, Ji G. Increased ARPP-19 Expression Is Associated with Hepatocellular Carcinoma. International Journal of Molecular Sciences. 2015; 16(1):178-192. https://doi.org/10.3390/ijms16010178
Chicago/Turabian StyleSong, Haiyan, Jielu Pan, Yang Liu, Hongzhu Wen, Lei Wang, Jiefeng Cui, Yinkun Liu, Bing Hu, Zemin Yao, and Guang Ji. 2015. "Increased ARPP-19 Expression Is Associated with Hepatocellular Carcinoma" International Journal of Molecular Sciences 16, no. 1: 178-192. https://doi.org/10.3390/ijms16010178
APA StyleSong, H., Pan, J., Liu, Y., Wen, H., Wang, L., Cui, J., Liu, Y., Hu, B., Yao, Z., & Ji, G. (2015). Increased ARPP-19 Expression Is Associated with Hepatocellular Carcinoma. International Journal of Molecular Sciences, 16(1), 178-192. https://doi.org/10.3390/ijms16010178