
Effect of Porcine Akirin2 on Skeletal Myosin Heavy Chain Isoform Expression

Abstract
:1. Introduction
2. Results
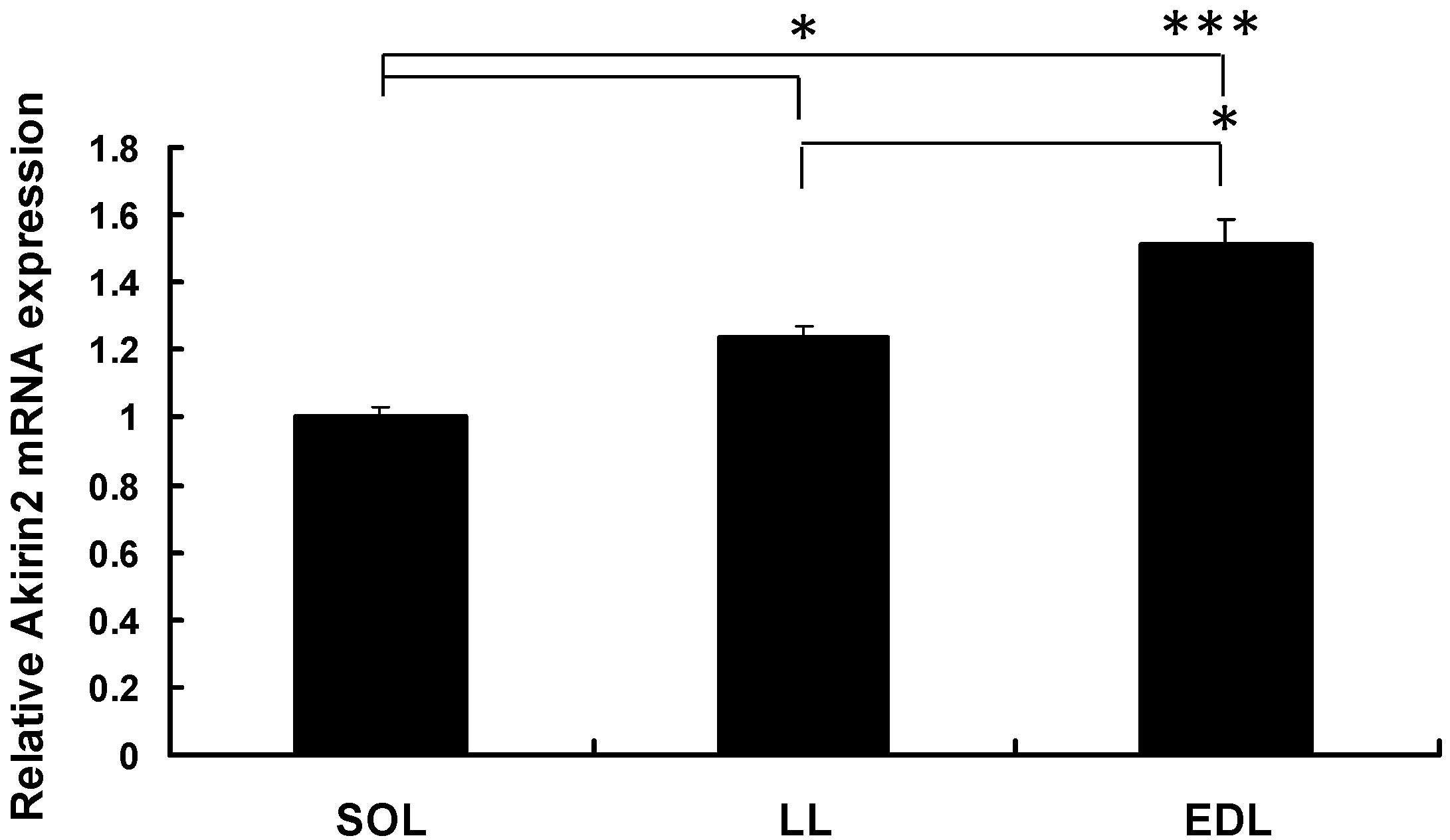
2.1. pAkirin2 mRNA Expression in Different Types of Muscle Tissues of Pigs


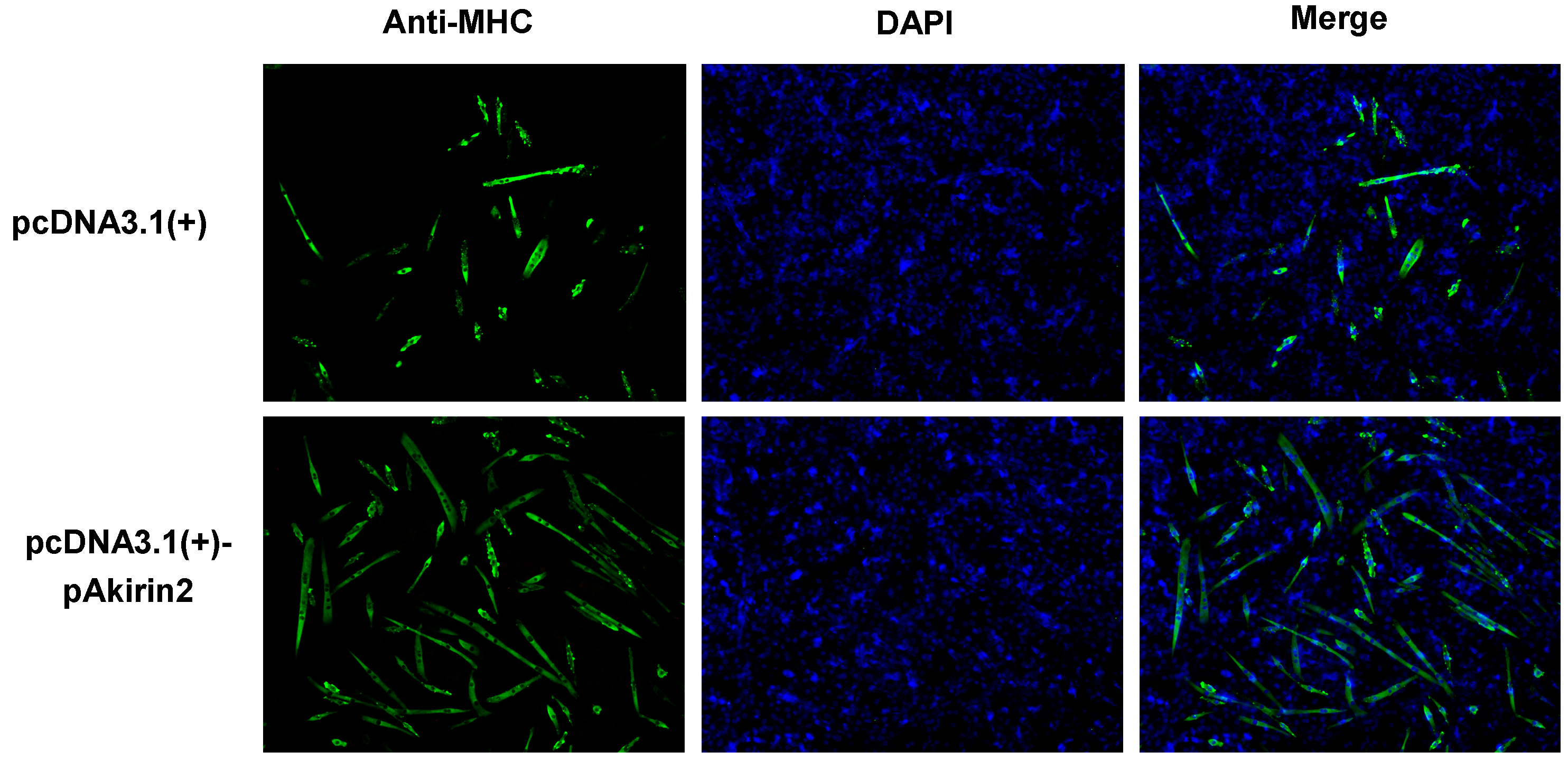
2.2. Effect of pAkirin2 on Myoblast Differentiation
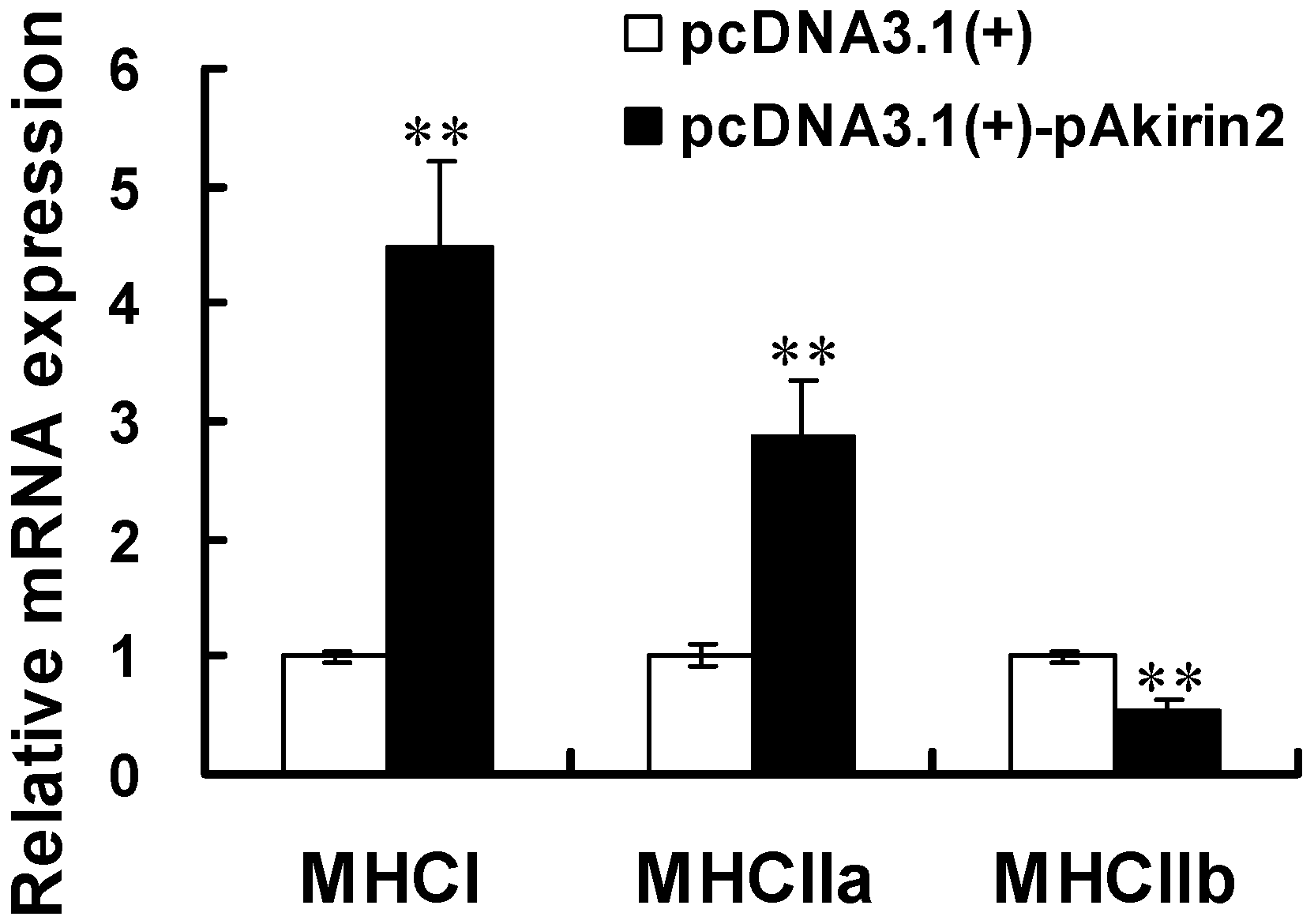
2.3. Effect of pAkirin2 on MHC Isoform Expression in C2C12 Myotubes

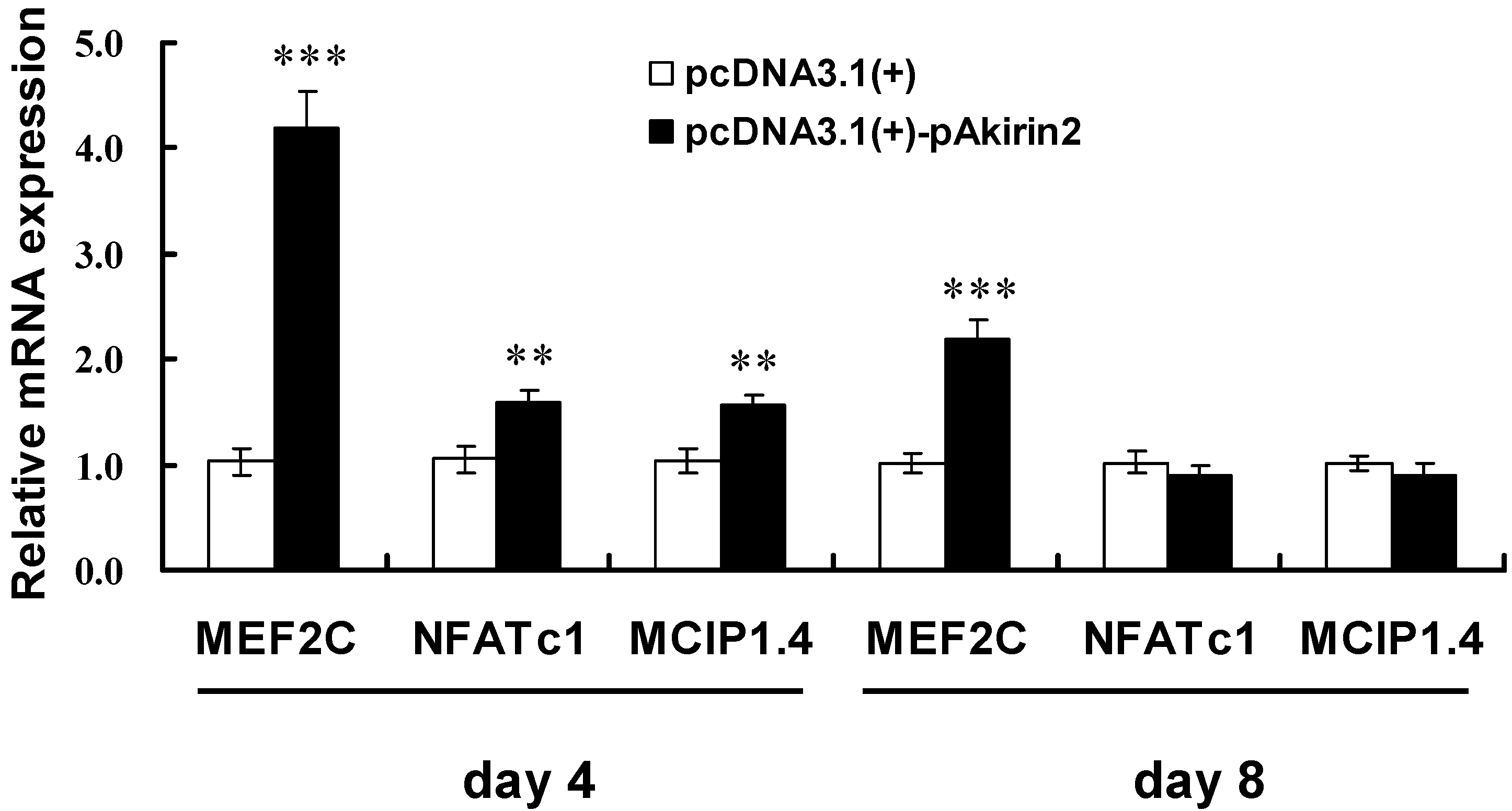
2.4. Effect of pAkirin2 on Oxidative Muscle Fiber Gene Expression in C2C12 Myotubes

3. Discussion
4. Materials and Methods
4.1. Animals and Tissue Sample Collection
4.2. RNA Isolation and Reverse Transcription
4.3. Cell Culture and Transfection
4.4. Real-Time Quantitative PCR

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Primer | Sequence | GenBank Accession No. | Product Size (bp) |
|---|---|---|---|---|
| MHCI | Forward | 5'-CTTCTACAGGCCTGGGCTTAC-3' | NM_080728 | 128 |
| Reverse | 5'-CTCCTTCTCAGACTTCCGCAG-3' | |||
| MHCIIa | Forward | 5'-TTCCAGAAGCCTAAGGTGGTC-3' | NM_001039545 | 94 |
| Reverse | 5'-GCCAGCCAGTGATGTTGTAAT-3' | |||
| MHCIIb | Forward | 5'-CTTGTCTGACTCAAGCCTGCC-3' | NM_010855 | 158 |
| Reverse | 5'-TCGCTCCTTTTCAGACTTCCG-3' | |||
| Akirin2 | Forward | 5'-GATGGGACTGGATTATCGC-3' | JN227885 | 154 |
| Reverse | 5'-GCACAAGATGAGTATGCGG-3' | |||
| MCIP1.4 | Forward | 5'-CCGTTGGCTGGAAACAAG-3' | NM_019466 | 153 |
| Reverse | 5'-GGTCACTCTCACACACGTGG-3' | |||
| NFATc1 | Forward | 5'-AATAACATGCGAGCCATCATC-3' | AF239169 | 109 |
| Reverse | 5'-TCACCCTGGTGTTCTTCCTC-3' | |||
| MEF2C | Forward | 5'-GATCTCCGCGTTCTTATCCC-3' | L13171 | 91 |
| Reverse | 5'-CCAATGACTGAGCCGACTG-3' | |||
| GAPDH | Forward | 5'-AGGGCATCTTGGGCTACAC-3' | NM_008084 | 211 |
| Reverse | 5'-TGGTCCAGGGTTTCTTACTCC-3' | |||
| mβ-actin | Forward | 5'-CCTTCCTTCTTGGGTATGGA-3' | NM_007393 | 88 |
| Reverse | 5'-GGTCTTTACGGATGTCAACG-3' | |||
| pβ-actin | Forward | 5'-CCACGAAACTACCTTCAACTCC-3' | DQ845171 | 132 |
| Reverse | 5'-GTGATCTCCTTCTGCATCCTGT-3' |
4.5. Cell Immunofluorescence Assay
4.6. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Brocks, L.; Klont, R.E.; Buist, W.; de Greef, K.; Tieman, M.; Engel, B. The effects of selection of pigs on growth rate vs. leanness on histochemical characteristics of different muscles. J. Anim. Sci. 2000, 78, 1247–1254. [Google Scholar] [PubMed]
- Ryu, Y.C.; Kim, B.C. Comparison of histochemical characteristics in various pork groups categorized by postmortem metabolic rate and pork quality. J. Anim. Sci. 2006, 84, 894–901. [Google Scholar] [PubMed]
- Brooke, M.H.; Kaiser, K.K. Muscle fiber types: How many and what kind? Arch. Neurol. 1970, 23, 369–379. [Google Scholar] [CrossRef] [PubMed]
- Levitsky, D.I. Actomyosin systems of biological motility. Biochemistry 2004, 69, 1177–1189. [Google Scholar] [PubMed]
- Lefaucheur, L.; Ecolan, P.; Plantard, L.; Gueguen, N. New insights into muscle fiber types in the pig. J. Histochem. Cytochem. 2002, 50, 719–730. [Google Scholar] [CrossRef] [PubMed]
- Kim, G.D.; Kim, B.W.; Jeong, J.Y.; Hur, S.J.; Cho, I.C.; Lim, H.T.; Joo, S.T. Relationship of carcassweight tomuscle fiber characteristics and pork quality of crossbred (Korean native black pig × Landrace) F2 pigs. Food Bioprocess Technol. 2013, 6, 522–529. [Google Scholar] [CrossRef]
- Lee, S.H.; Joo, S.T.; Ryu, Y.C. Skeletal muscle fiber type and myofibrillar proteins in relation to meat quality. Meat Sci. 2010, 86, 166–170. [Google Scholar] [CrossRef] [PubMed]
- Goto, A.; Matsushita, K.; Gesellchen, V.; Chamy, L.E.; Kuttenkeuler, D.; Takeuchi, O.; Hoffmann, J.A.; Akira, S.; Boutros, M.; Reichhart, J. Akirins are highly conserved nuclear proteins required for NF-κB-dependent gene expression in drosophila and mice. Nat. Immunol. 2008, 9, 97–104. [Google Scholar] [CrossRef] [PubMed]
- Beutler, B.; Moresco, E.M.Y. Akirins versus infection. Nat. Immunol. 2008, 9, 7–9. [Google Scholar] [CrossRef] [PubMed]
- Macqueen, D.J.; Johnston, I.A. Evolution of the multifaceted eukaryotic Akirin gene family. BMC Evol. Biol. 2009, 9, 34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Macqueen, D.J.; Bower, N.I.; Johnston, I.A. Positioning the expanded Akirin gene family of Atlantic salmon within the transcriptional networks of myogenesis. Biochem. Biophys. Res. Commun. 2010, 400, 599–605. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.L.; Huang, Z.Q.; Wang, H.; Jia, G.; Liu, G.M.; Guo, X.L.; Tang, R.Y.; Long, D.B. Role of Akirin in skeletal myogenesis. Int. J. Mol. Sci. 2013, 14, 3817–3823. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, Y.; Nagai, K.; Nagata, Y.; Doronbekov, K.; Nishimura, S.; Yoshioka, S.; Fujita, T.; Shiga, K.; Miyake, T.; Taniguchi, Y.; et al. Exploration of genes showing intramuscular fat deposition-associated changes in musculus longissimus muscle. Anim. Genet. 2006, 37, 40–46. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, S.; Yamada, T.; Sukegawa, S.; Miyake, T.; Fujita, T.; Morita, M.; Ohta, T.; Takahagi, Y.; Murakami, H.; Morimatsu, F.; et al. Association of a single nucleotide polymorphism in Akirin2 gene with marbling in Japanese Black beef cattle. BMC Res. Notes 2009, 2, 131–135. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, N.; Satoh, Y.; Fujita, T.; Ohta, T.; Kose, H.; Muramatsu, Y.; Yamamoto, T.; Yamada, T. Distribution of allele frequencies at TTN g.231054C>T, RPL27A g.3109537C>T and Akirin2 c.*188G>A between Japanese black and four other cattle breeds with differing historical selection for marbling. BMC Res. Notes 2011, 4, 10. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Lee, S.K.; Hong, M.W.; Park, S.R.; Lee, Y.S.; Kim, J.W.; Lee, H.K.; Jeong, D.K.; Song, Y.H.; Lee, S.J. Association of a single nucleotide polymorphism in the Akirin2 gene with economically important traits in Korean native cattle. Anim. Genet. 2013, 44, 750–753. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.L.; Huang, Z.Q.; Jia, G.; Wu, X.Q.; Wu, C.M. Molecular cloning, tissue distribution, and functional analysis of porcine Akirin2. Anim. Biotechnol. 2012, 23, 124–131. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Huang, Z.; Zhou, B.; Wang, H.; Jia, G.; Qiao, J. Expression and purification of porcine Akirin2 in Escherichia coli. Turk. J. Biol. 2014, 38, 339–345. [Google Scholar] [CrossRef]
- Hwang, Y.H.; Kim, G.D.; Jeong, J.Y.; Hur, S.J.; Joo, S.T. The relationship between muscle fiber characteristics and meat quality traits of highly marbled Hanwoo (Korean native cattle) steers. Meat Sci. 2010, 86, 456–461. [Google Scholar] [CrossRef] [PubMed]
- Chang, K.C.; da Costa, N.; Blackley, R.; Southwood, O.; Evans, G.; Plastow, G.; Wood, J.D.; Richardson, R.I. Relationships of myosin heavy chain fibre types to meat quality traits in traditional and modern pigs. Meat Sci. 2003, 64, 93–103. [Google Scholar] [CrossRef] [PubMed]
- Ryu, Y.C.; Choi, Y.M.; Lee, S.H.; Shin, H.G.; Choe, J.H.; Kim, J.M.; Hong, K.C.; Kim, B.C. Comparing the histochemical characteristics and meat quality traits of different pig breeds. Meat Sci. 2008, 80, 363–369. [Google Scholar] [CrossRef] [PubMed]
- Ryu, Y.C.; Kim, B.C. The relationship between muscle fiber characteristics, postmortem metabolic rate, and meat quality of pig longissimus dorsi muscle. Meat Sci. 2005, 71, 351–357. [Google Scholar] [CrossRef] [PubMed]
- Chin, E.R.; Olson, E.N.; Richardson, J.A.; Yang, Q.; Humphries, C.; Shelton, J.M.; Wu, H.; Zhu, W.; Bassel-Duby, R.; Williams, R.S. A calcineurin-dependent transcriptional pathway controls skeletal muscle fiber type. Genes Dev. 1998, 12, 2499–2509. [Google Scholar] [CrossRef] [PubMed]
- Bassel-Duby, R.; Olson, E.N. Signaling pathways in skeletal muscle remodeling. Annu. Rev. Biochem. 2009, 75, 19–37. [Google Scholar] [CrossRef]
- Olson, E.N.; Williams, R.S. Calcineurin signaling and muscle remodeling. Cell 2000, 101, 689–692. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Rothermel, B.; Kanatous, S.; Rosenberg, P.; Naya, F.J.; Shelton, J.M.; Hutcheson, K.A.; DiMaio, J.M.; Olson, E.N.; Bassel-Duby, R.; et al. Activation of MEF2 by muscle activity is mediated through a calcineurin dependent pathway. EMBO J. 2001, 20, 6414–6423. [Google Scholar] [CrossRef] [PubMed]
- Mu, X.; Brown, L.D.; Liu, Y.; Schneider, M.F. Roles of the calcineurin and CaMK signaling pathways in fast-to-slow fiber type transformation of cultured adult mouse skeletal muscle fibers. Physiol. Genomics 2007, 30, 300–312. [Google Scholar] [CrossRef] [PubMed]
- Blaeser, F.; Ho, N.; Prywes, R.; Chatila, T.A. Ca2+-dependent gene expression mediated by MEF2 transcription factors. J. Biol. Chem. 2000, 275, 197–209. [Google Scholar] [CrossRef] [PubMed]
- Abbott, K.L.; Friday, B.B.; Thaloor, D.; Murphy, T.J.; Pavlath, G.K. Activation and cellular localization of the cyclosporine A-sensitive transcription factor NF-AT in skeletal muscle cells. Mol. Biol. Cell 1998, 9, 2905–2916. [Google Scholar] [CrossRef] [PubMed]
- Im, S.H.; Rao, A. Activation and deactivation of gene expression by Ca2+/calcineurin-NFAT-mediated signaling. Mol. Cells 2004, 18, 1–9. [Google Scholar] [PubMed]
- Meissner, J.D.; Freund, R.; Krone, D.; Umeda, P.K.; Chang, K.C.; Gros, G.; Scheibe, R.J. Extracellular signal-regulated kinase 1/2-mediated phosphorylation of p300 enhances myosin heavy chain I/β gene expression via acetylation of nuclear factor of activated T cells c1. Nucleic Acids Res. 2011, 39, 5907–5925. [Google Scholar] [CrossRef] [PubMed]
- Calabria, E.; Ciciliot, S.; Moretti, I.; Garcia, M.; Picard, A.; Dyar, K.A.; Pallafacchina, G.; Tothova, J.; Schiaffino, S.; Murgia, M. FAT isoforms control activity-dependent muscle fiber type specification. Proc. Natl. Acad. Sci. USA 2009, 106, 13335–13340. [Google Scholar] [CrossRef] [PubMed]
- Meissner, J.D.; Umeda, P.K.; Chang, K.C.; Gros, G.; Scheibe, R.J. Activation of the β myosin heavy chain promoter by MEF-2D, MyoD, p300, and the calcineurin/NFATc1 pathway. J. Cell. Physiol. 2007, 211, 138–148. [Google Scholar] [CrossRef] [PubMed]
- Allen, D.L.; Leinwand, L.A. Intracellular calcium and myosin isoform transitions. Calcineurin and calcium-calmodulin kinase pathways regulate preferential activation of the IIa myosin heavy chain promoter. J. Biol. Chem. 2002, 277, 45323–45330. [Google Scholar] [CrossRef] [PubMed]
- Meissner, J.D.; Chang, K.C.; Kubis, H.P.; Nebreda, A.R.; Gros, G.; Scheibe, R.J. The p38α/β mitogen-activated protein kinases mediate recruitment of CREB-binding protein to preserve fast myosin heavy chain IId/x gene activity in myotubes. J. Biol. Chem. 2007, 282, 7265–7275. [Google Scholar] [CrossRef] [PubMed]
- Serfling, E.; Chuvpilo, S.; Liu, J.; Höfer, T.; Palmetshofer, A. NFATc1 autoregulation: A crucial step for cell-fate determination. Trends Immunol. 2006, 27, 461–469. [Google Scholar] [CrossRef] [PubMed]
- Davies, K.J.; Ermak, G.; Rothermel, B.A.; Pritchard, M.; Heitman, J.; Ahnn, J.; Henrique-Silva, F.; Crawford, D.; Canaider, S.; Strippoli, P.; et al. Renaming the DSCR1/Adapt78 gene family as RCAN: Regulators of calcineurin. FASEB J. 2007, 21, 3023–3028. [Google Scholar] [CrossRef] [PubMed]
- Fenyvesi, R.; Rácz, G.; Wuytack, F.; Zádor, E. The calcineurin activity and MCIP1.4 mRNA levels are increased by innervation in regenerating soleus muscle. Biochem. Biophys. Res. Commun. 2004, 320, 599–605. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2–ΔΔCt method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, X.; Luo, Y.; Zhou, B.; Huang, Z.; Jia, G.; Liu, G.; Zhao, H.; Yang, Z.; Zhang, R. Effect of Porcine Akirin2 on Skeletal Myosin Heavy Chain Isoform Expression. Int. J. Mol. Sci. 2015, 16, 3996-4006. https://doi.org/10.3390/ijms16023996
Chen X, Luo Y, Zhou B, Huang Z, Jia G, Liu G, Zhao H, Yang Z, Zhang R. Effect of Porcine Akirin2 on Skeletal Myosin Heavy Chain Isoform Expression. International Journal of Molecular Sciences. 2015; 16(2):3996-4006. https://doi.org/10.3390/ijms16023996
Chicago/Turabian StyleChen, Xiaoling, Yanliu Luo, Bo Zhou, Zhiqing Huang, Gang Jia, Guangmang Liu, Hua Zhao, Zhouping Yang, and Ruinan Zhang. 2015. "Effect of Porcine Akirin2 on Skeletal Myosin Heavy Chain Isoform Expression" International Journal of Molecular Sciences 16, no. 2: 3996-4006. https://doi.org/10.3390/ijms16023996
APA StyleChen, X., Luo, Y., Zhou, B., Huang, Z., Jia, G., Liu, G., Zhao, H., Yang, Z., & Zhang, R. (2015). Effect of Porcine Akirin2 on Skeletal Myosin Heavy Chain Isoform Expression. International Journal of Molecular Sciences, 16(2), 3996-4006. https://doi.org/10.3390/ijms16023996