Biological Functions of Thyroid Hormone in Placenta


Abstract
:1. Introduction
2. The Relationship between Inflammatory Processes and Placental Disorders
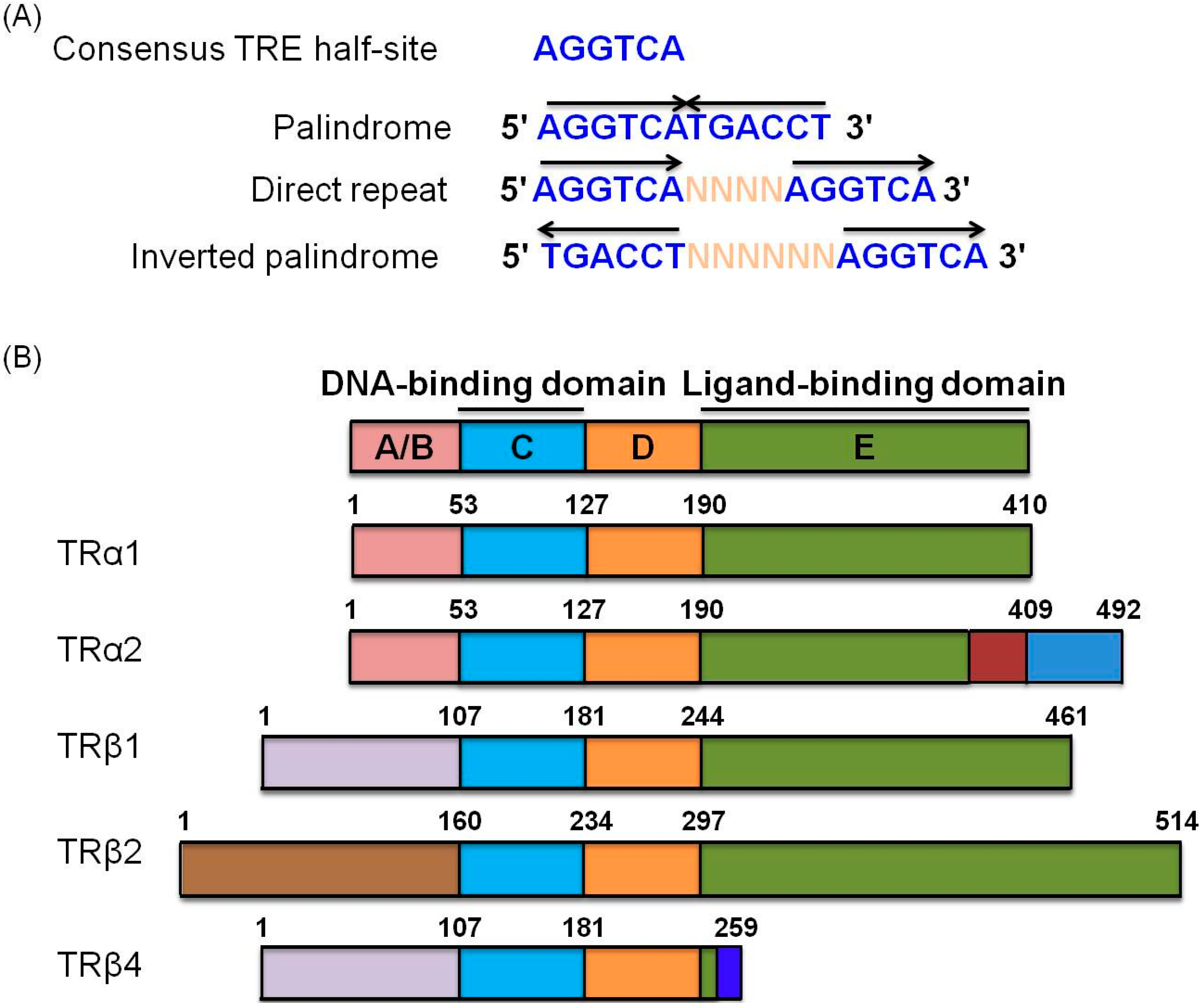
3. Nuclear Receptor Family
4. Thyroid Hormone Involvement in Inflammation Processes

5. Thyroid Hormone Is Associated with Placental Development and Disease
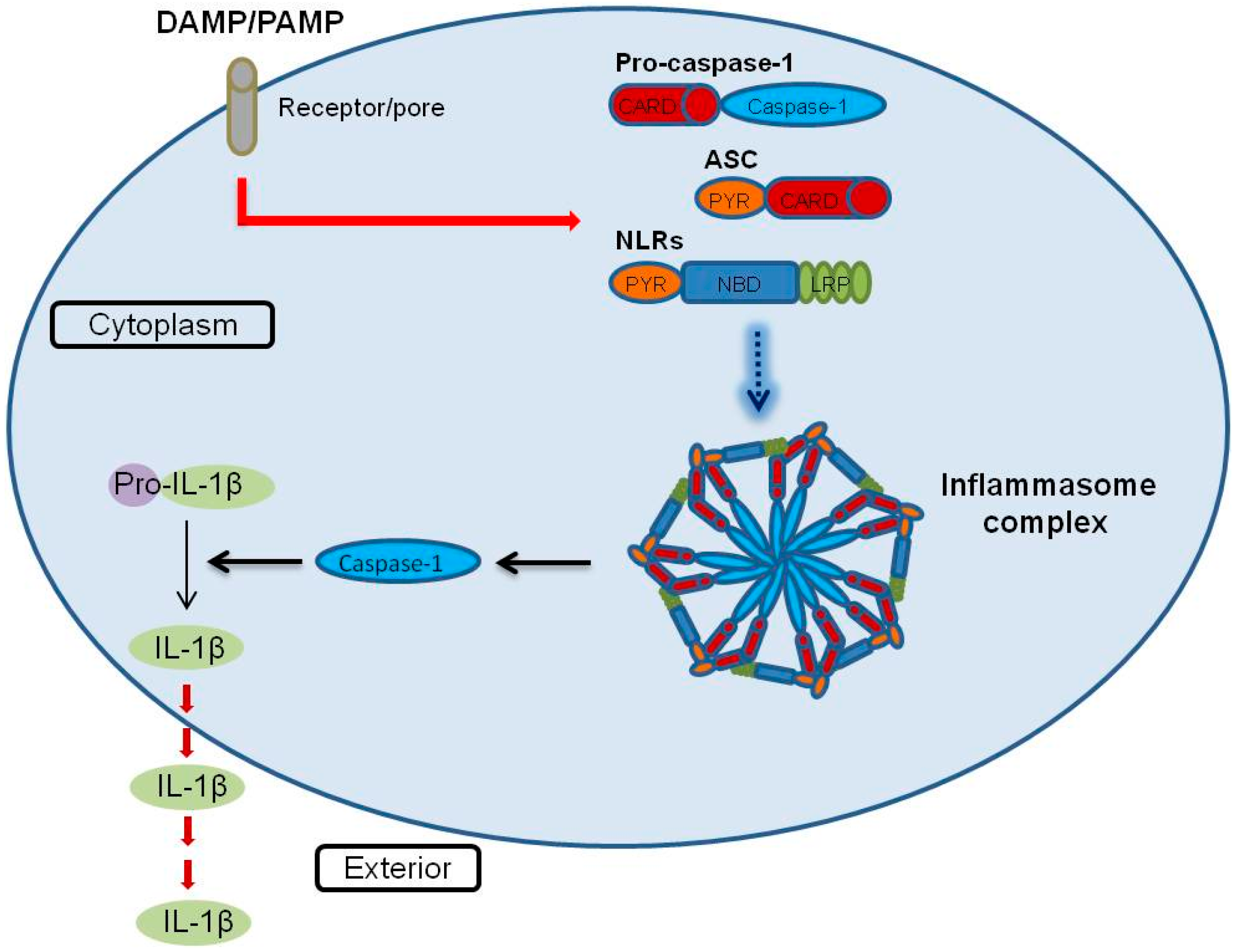
6. Inflammasome Functions in Placental Disease


{kind=link}
{kind=link}
| NLRs | DAMP | PAMP |
|---|---|---|
| NOD1 | Bacterial peptidoglycan, γ-d-glutamyl-meso-diaminopimelic acid | |
| NOD2 | Bacterial peptidoglycan, Muramyl dipeptide | |
| IPAF | Bacterial (Salmonella typhimurium, Pseudomonas aeruginosa, Legionellapneumophila) | |
| NALP1 | Bacterial | |
| NALP3 | Hyaluronan, uric acid crystals, ATP, silica, asbestos, alum | Bacteria (Staphylococcus aureus, Shigella flexneri), viruses (Adenovirus, Influenza virus), fungi (Saccharomyces cerevisiae), muramyl dipeptide, bacterial pore-forming toxin, bacteria RNA, malarial crystals |
7. Conclusions
Acknowledgments
Conflicts of Interest
References
- Everley, P.A.; Krijgsveld, J.; Zetter, B.R.; Gygi, S.P. Quantitative cancer proteomics: Stable isotope labeling with amino acids in cell culture (SILAC) as a tool for prostate cancer research. Mol. Cell. Proteomics 2004, 3, 729–735. [Google Scholar] [CrossRef] [PubMed]
- Uitto, P.M.; Lance, B.K.; Wood, G.R.; Sherman, J.; Baker, M.S.; Molloy, M.P. Comparing SILAC and two-dimensional gel electrophoresis image analysis for profiling urokinase plasminogen activator signaling in ovarian cancer cells. J. Proteome Res. 2007, 6, 2105–2112. [Google Scholar] [CrossRef] [PubMed]
- Planque, C.; Kulasingam, V.; Smith, C.R.; Reckamp, K.; Goodglick, L.; Diamandis, E.P. Identification of five candidate lung cancer biomarkers by proteomics analysis of conditioned media of four lung cancer cell lines. Mol. Cell. Proteomics 2009, 8, 2746–2758. [Google Scholar] [CrossRef] [PubMed]
- Gronborg, M.; Kristiansen, T.Z.; Iwahori, A.; Chang, R.; Reddy, R.; Sato, N.; Molina, H.; Jensen, O.N.; Hruban, R.H.; Goggins, M.G.; et al. Biomarker discovery from pancreatic cancer secretome using a differential proteomic approach. Mol. Cell. Proteomics 2006, 5, 157–171. [Google Scholar] [CrossRef]
- Chen, C.Y.; Chi, L.M.; Chi, H.C.; Tsai, M.M.; Tsai, C.Y.; Tseng, Y.H.; Lin, Y.H.; Chen, W.J.; Huang, Y.H.; Lin, K.H. Stable isotope labeling with amino acids in cell culture (SILAC)-based quantitative proteomics study of a thyroid hormone-regulated secretome in human hepatoma cells. Mol. Cell. Proteomics 2012, 11. [Google Scholar] [CrossRef] [PubMed]
- Cho, C.K.; Shan, S.J.; Winsor, E.J.; Diamandis, E.P. Proteomics analysis of human amniotic fluid. Mol. Cell. Proteomics 2007, 6, 1406–1415. [Google Scholar] [CrossRef] [PubMed]
- Kingsmore, S.F.; Kennedy, N.; Halliday, H.L.; van Velkinburgh, J.C.; Zhong, S.; Gabriel, V.; Grant, J.; Beavis, W.D.; Tchernev, V.T.; Perlee, L.; et al. Identification of diagnostic biomarkers for infection in premature neonates. Mol. Cell. Proteomics 2008, 7, 1863–1875. [Google Scholar] [CrossRef]
- Wang, L.J.; Hsu, C.W.; Chen, C.C.; Liang, Y.; Chen, L.C.; Ojcius, D.M.; Tsang, N.M.; Hsueh, C.; Wu, C.C.; Chang, Y.S. Interactome-wide analysis identifies end-binding protein 1 as a crucial component for the speck-like particle formation of activated absence in melanoma 2 (AIM2) inflammasomes. Mol. Cell. Proteomics 2012, 11, 1230–1244. [Google Scholar] [CrossRef] [PubMed]
- Saraon, P.; Cretu, D.; Musrap, N.; Karagiannis, G.S.; Batruch, I.; Drabovich, A.P.; van der Kwast, T.; Mizokami, A.; Morrissey, C.; Jarvi, K.; et al. Quantitative proteomics reveals that enzymes of the ketogenic pathway are associated with prostate cancer progression. Mol. Cell. Proteomics 2013, 12, 1589–15601. [Google Scholar] [CrossRef]
- Chen, R.; Pan, S.; Brentnall, T.A.; Aebersold, R. Proteomic profiling of pancreatic cancer for biomarker discovery. Mol. Cell. Proteomics 2005, 4, 523–533. [Google Scholar] [CrossRef] [PubMed]
- Andreyev, A.Y.; Shen, Z.; Guan, Z.; Ryan, A.; Fahy, E.; Subramaniam, S.; Raetz, C.R.; Briggs, S.; Dennis, E.A. Application of proteomic marker ensembles to subcellular organelle identification. Mol. Cell. Proteomics 2010, 9, 388–402. [Google Scholar] [CrossRef] [PubMed]
- Bell, C.; English, L.; Boulais, J.; Chemali, M.; Caron-Lizotte, O.; Desjardins, M.; Thibault, P. Quantitative proteomics reveals the induction of mitophagy in tumor necrosis factor-α-activated (TNFα) macrophages. Mol. Cell. Proteomics 2013, 12, 2394–2407. [Google Scholar] [CrossRef] [PubMed]
- Ingbar, S.H. Pre-albumin: A thyroxinebinding protein of human plasma. Endocrinology 1958, 63, 256–259. [Google Scholar] [CrossRef] [PubMed]
- Richardson, S.J. Evolutionary changes to transthyretin: Evolution of transthyretin biosynthesis. FEBS J. 2009, 276, 5342–5356. [Google Scholar] [CrossRef] [PubMed]
- Dickson, P.W.; Aldred, A.R.; Marley, P.D.; Bannister, D.; Schreiber, G. Rat choroid plexus specializes in the synthesis and the secretion of transthyretin (prealbumin). Regulation of transthyretin synthesis in choroid plexus is independent from that in liver. J. Biol. Chem. 1986, 261, 3475–3478. [Google Scholar] [PubMed]
- Roef, G.L.; Rietzschel, E.R.; van Daele, C.M.; Taes, Y.E.; de Buyzere, M.L.; Gillebert, T.C.; Kaufman, J.M. Triiodothyronine and free thyroxine levels are differentially associated with metabolic profile and adiposity-related cardiovascular risk markers in euthyroid middle-aged subjects. Thyroid 2014, 24, 223–231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rinaldi, S.F.; Hutchinson, J.L.; Rossi, A.G.; Norman, J.E. Anti-inflammatory mediators as physiological and pharmacological regulators of parturition. Expert Rev. Clin. Immunol. 2011, 7, 675–696. [Google Scholar] [CrossRef] [PubMed]
- Challis, J.R.; Lockwood, C.J.; Myatt, L.; Norman, J.E.; Strauss, J.F.; Petraglia, F. Inflammation and Pregnancy. Reprod. Sci. 2009, 16, 206–215. [Google Scholar] [CrossRef] [PubMed]
- Romero, R.; Espinoza, J.; Kusanovic, J.P.; Gotsch, F.; Hassan, S.; Erez, O.; Chaiworapongsa, T.; Mazor, M. The preterm parturition syndrome. BJOG 2006, 113, 17–42. [Google Scholar] [CrossRef] [PubMed]
- Gotsch, F.; Romero, R.; Erez, O.; Vaisbuch, E.; Kusanovic, J.P.; Mazaki-Tovi, S.; Kim, S.K.; Hassan, S.; Yeo, L. The preterm parturition syndrome and its implications for understanding the biology, risk assessment, diagnosis, treatment and prevention of preterm birth. J. Matern. Fetal Neonatal Med. 2009, 22, 5–23. [Google Scholar] [CrossRef] [PubMed]
- Goldenberg, R.L.; Culhane, J.F.; Iams, J.D.; Romero, R. Epidemiology and causes of preterm birth. Lancet 2008, 371, 75–84. [Google Scholar] [CrossRef] [PubMed]
- Brosens, I.; Renaer, M. On the pathogenesis of placental infarcts in pre-eclampsia. J. Obstet. Gynaecol. Br. Commonw. 1972, 79, 794–799. [Google Scholar] [CrossRef] [PubMed]
- Pijnenborg, R.; Anthony, J.; Davey, D.A.; Rees, A.; Tiltman, A.; Vercruysse, L.; van Assche, A. Placental bed spiral arteries in the hypertensive disorders of pregnancy. Br. J. Obstet. Gynaecol. 1991, 98, 648–655. [Google Scholar] [CrossRef] [PubMed]
- Redman, C.W.; Sargent, I.L. Pre-eclampsia, the placenta and the maternal systemic inflammatory response—A review. Placenta 2003, 24, S21–S27. [Google Scholar] [CrossRef] [PubMed]
- Redman, C.W.; Sacks, G.P.; Sargent, I.L. Preeclampsia: An excessive maternal inflammatory response to pregnancy. Am. J. Obstet. Gynecol. 1999, 180, 499–506. [Google Scholar] [CrossRef] [PubMed]
- Sacks, G.P.; Studena, K.; Sargent, K.; Redman, C.W. Normal pregnancy and preeclampsia both produce inflammatory changes in peripheral blood leukocytes akin to those of sepsis. Am. J. Obstet. Gynecol. 1998, 179, 80–86. [Google Scholar] [CrossRef] [PubMed]
- Greer, I.A.; Lyall, F.; Perera, T.; Boswell, F.; Macara, L.M. Increased concentrations of cytokines interleukin-6 and interleukin-1 receptor antagonist in plasma of women with preeclampsia: A mechanism for endothelial dysfunction? Obstet. Gynecol. 1994, 84, 937–940. [Google Scholar] [PubMed]
- Vince, G.S.; Starkey, P.M.; Austgulen, R.; Kwiatkowski, D.; Redman, C.W. Interleukin-6, tumour necrosis factor and soluble tumour necrosis factor receptors in women with pre-eclampsia. Br. J. Obstet. Gynaecol. 1995, 102, 20–25. [Google Scholar] [CrossRef] [PubMed]
- Ellis, J.; Wennerholm, U.B.; Bengtsson, A.; Lilja, H.; Pettersson, A.; Sultan, B.; Wennergren, M.; Hagberg, H. Levels of dimethylarginines and cytokines in mild and severe preeclampsia. Acta Obstet. Gynecol. Scand. 2001, 80, 602–608. [Google Scholar] [CrossRef] [PubMed]
- Benyo, D.F.; Smarason, A.; Redman, C.W.; Sims, C.; Conrad, K.P. Expression of inflammatory cytokines in placentas from women with preeclampsia. J. Clin. Endocrinol. Metab. 2001, 86, 2505–2512. [Google Scholar] [PubMed]
- Mulla, M.J.; Salmon, J.E.; Chamley, L.W.; Brosens, J.J.; Boeras, C.M.; Kavathas, P.B.; Abrahams, V.M. A role for uric acid and the Nalp3 inflammasome in antiphospholipid antibody-induced IL-1β production by human first trimester trophoblast. PLoS One 2013, 8, e65237. [Google Scholar] [CrossRef] [PubMed]
- Xie, F.; Hu, Y.; Turvey, S.E.; Magee, L.A.; Brunham, R.M.; Choi, K.C.; Krajden, M.; Leung, P.C.; Money, D.M.; Patrick, D.M.; et al. Toll-like receptors 2 and 4 and the cryopyrin inflammasome in normal pregnancy and pre-eclampsia. BJOG 2010, 117, 99–108. [Google Scholar] [CrossRef]
- Margolis, R.N.; Christakos, S. The nuclear receptor superfamily of steroid hormones and vitamin D gene regulation. An update. Ann. N. Y. Acad. Sci. 2010, 1192, 208–214. [Google Scholar] [CrossRef] [PubMed]
- Mangelsdorf, D.J.; Thummel, C.; Beato, M.; Herrlich, P.; Schutz, G.; Umesono, K.; Blumberg, B.; Kastner, P.; Mark, M.; Chambon, P.; et al. The nuclear receptor superfamily: The second decade. Cell 1995, 83, 835–859. [Google Scholar] [CrossRef]
- Zhang, Z.; Burch, P.E.; Cooney, A.J.; Lanz, R.B.; Pereira, F.A.; Wu, J.; Gibbs, R.A.; Weinstock, G.; Wheeler, D.A. Genomic analysis of the nuclear receptor family: New insights into structure, regulation, and evolution from the rat genome. Genome Res. 2004, 14, 580–590. [Google Scholar] [CrossRef] [PubMed]
- Jin, L.; Li, Y. Structural and functional insights into nuclear receptor signaling. Adv. Drug Deliv. Rev. 2010, 62, 1218–1226. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.Y.; Tsai, M.M.; Chi, H.C.; Lin, K.H. Biological significance of a thyroid hormone-regulated secretome. Biochim. Biophys. Acta 2013, 1834, 2271–2184. [Google Scholar] [CrossRef] [PubMed]
- Arrese, M.; Karpen, S.J. Nuclear receptors, inflammation, and liver disease: Insights for cholestatic and fatty liver diseases. Clin. Pharmacol. Ther. 2010, 87, 473–478. [Google Scholar] [CrossRef] [PubMed]
- Trauner, M.; Halilbasic, E. Nuclear receptors as new perspective for the management of liver diseases. Gastroenterology 2011, 140, 1120–1125. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Ding, C.; Liu, W.; Song, L.; Liu, M.; Qi, L.; Fu, T.; Malovannaya, A.; Wang, Y.; Qin, J.; et al. In-depth proteomic characterization of endogenous nuclear receptors in mouse liver. Mol. Cell. Proteomics 2013, 12, 473–484. [Google Scholar] [CrossRef] [PubMed]
- Aranda, A.; Pascual, A. Nuclear hormone receptors and gene expression. Physiol. Rev. 2001, 81, 1269–1304. [Google Scholar] [PubMed]
- Walters, M.R.; Nemere, I. Receptors for steroid hormones: Membrane-associated and nuclear forms. Cell. Mol. Life Sci. 2004, 61, 2309–2321. [Google Scholar] [CrossRef] [PubMed]
- Kim, W.G.; Cheng, S.Y. Thyroid hormone receptors and cancer. Biochim. Biophys. Acta 2012, 1830, 3928–3936. [Google Scholar] [CrossRef] [PubMed]
- Cheng, S.Y. Multiple mechanisms for regulation of the transcriptional activity of thyroid hormone receptors. Rev. Endocr. Metab. Disord. 2000, 1, 9–18. [Google Scholar] [CrossRef] [PubMed]
- Izumo, S.; Mahdavi, V. Thyroid hormone receptor α isoforms generated by alternative splicing differentially activate myosin HC gene transcription. Nature 1988, 334, 539–542. [Google Scholar] [CrossRef] [PubMed]
- Mitsuhashi, T.; Tennyson, G.E.; Nikodem, V.M. Alternative splicing generates messages encoding rat C-ERBA proteins that do not bind thyroid hormone. Proc. Natl. Acad. Sci. USA 1988, 85, 5804–5808. [Google Scholar] [CrossRef] [PubMed]
- Nakai, A.; Seino, S.; Sakurai, A.; Szilak, I.; Bell, G.I.; DeGroot, L.J. Characterization of a thyroid hormone receptor expressed in human kidney and other tissues. Proc. Natl. Acad. Sci. USA 1988, 85, 2781–2785. [Google Scholar] [CrossRef] [PubMed]
- Yen, P.M.; Chin, W.W. New advances in understanding the molecular mechanisms of thyroid hormone action. Trends Endocrinol. Metab. 1994, 5, 65–72. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.Y.; Chung, I.H.; Tsai, M.M.; Tseng, Y.H.; Chi, H.C.; Tsai, C.Y.; Lin, Y.H.; Wang, Y.C.; Chen, C.P.; Wu, T.I.; et al. Thyroid hormone enhanced human hepatoma cell motility involves brain-specific serine protease 4 activation via ERK signaling. Mol. Cancer 2014, 13. [Google Scholar] [CrossRef]
- Lin, K.H.; Chen, C.Y.; Chen, S.L.; Yen, C.C.; Huang, Y.H.; Shih, C.H.; Shen, J.J.; Yang, R.C.; Wang, C.S. Regulation of fibronectin by thyroid hormone receptors. J. Mol. Endocrinol. 2004, 33, 445–458. [Google Scholar] [CrossRef] [PubMed]
- Lin, K.H.; Lee, H.Y.; Shih, C.H.; Yen, C.C.; Chen, S.L.; Yang, R.C.; Wang, C.S. Plasma protein regulation by thyroid hormone. J. Endocrinol. 2003, 179, 367–377. [Google Scholar] [CrossRef] [PubMed]
- Tai, P.J.; Huang, Y.H.; Shih, C.H.; Chen, R.N.; Chen, C.D.; Chen, W.J.; Wang, C.S.; Lin, K.H. Direct regulation of androgen receptor-associated protein 70 by thyroid hormone and its receptors. Endocrinology 2007, 148, 3485–3495. [Google Scholar] [CrossRef] [PubMed]
- Lin, K.H.; Shieh, H.Y.; Hsu, H.C. Negative regulation of the antimetastatic gene Nm23-H1 by thyroid hormone receptors. Endocrinology 2000, 141, 2540–2547. [Google Scholar] [PubMed]
- Kwakkel, J.; Surovtseva, O.V.; de Vries, E.M.; Stap, J.; Fliers, E.; Boelen, A. A novel role for the thyroid hormone-activating enzyme type 2 deiodinase in the inflammatory response of macrophages. Endocrinology 2014, 155, 2725–2734. [Google Scholar] [CrossRef] [PubMed]
- Moore, J.M.; Guy, R.K. Coregulator interactions with the thyroid hormone receptor. Mol. Cell. Proteomics 2005, 4, 475–482. [Google Scholar] [CrossRef] [PubMed]
- Flier, J.S.; Harris, M.; Hollenberg, A.N. Leptin, nutrition, and the thyroid: The why, the wherefore, and the wiring. J. Clin. Investig. 2000, 105, 859–861. [Google Scholar] [CrossRef] [PubMed]
- Konner, A.C.; Bruning, J.C. Toll-like receptors: Linking inflammation to metabolism. Trends Endocrinol. Metab. 2011, 22, 16–23. [Google Scholar] [CrossRef] [PubMed]
- Fresno, M.; Alvarez, R.; Cuesta, N. Toll-like receptors, inflammation, metabolism and obesity. Arch. Physiol. Biochem. 2011, 117, 151–164. [Google Scholar] [CrossRef] [PubMed]
- Duntas, L.H.; Biondi, B. The interconnections between obesity, thyroid function, and autoimmunity: The multifold role of leptin. Thyroid 2013, 23, 646–653. [Google Scholar] [CrossRef] [PubMed]
- Haddow, J.E.; Palomaki, G.E.; Allan, W.C.; Williams, J.R.; Knight, G.J.; Gagnon, J.; O'Heir, C.E.; Mitchell, M.L.; Hermos, R.J.; Waisbren, S.E.; et al. Maternal thyroid deficiency during pregnancy and subsequent neuropsychological development of the child. N. Engl. J. Med. 1999, 341, 549–555. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Shan, Z.; Teng, W.; Yu, X.; Fan, C.; Teng, X.; Guo, R.; Wang, H.; Li, J.; Chen, Y.; et al. Abnormalities of maternal thyroid function during pregnancy affect neuropsychological development of their children at 25–30 months. Clin. Endocrinol. 2010, 72, 825–829. [Google Scholar] [CrossRef]
- Costa, A.; Arisio, R.; Benedetto, C.; Bertino, E.; Fabris, C.; Giraudi, G.; Marozio, L.; Maula, V.; Pagliano, M.; Testori, O.; et al. Thyroid hormones in tissues from human embryos and fetuses. J. Endocrinol. Investig. 1991, 14, 559–568. [Google Scholar] [CrossRef]
- Chan, S.; Kachilele, S.; McCabe, C.J.; Tannahill, L.A.; Boelaert, K.; Gittoes, N.J.; Visser, T.J.; Franklyn, J.A.; Kilby, M.D. Early expression of thyroid hormone deiodinases and receptors in human fetal cerebral cortex. Brain Res. Dev. Brain Res. 2002, 138, 109–116. [Google Scholar] [CrossRef] [PubMed]
- Chan, S.Y.; Martin-Santos, A.; Loubiere, L.S.; Gonzalez, A.M.; Stieger, B.; Logan, A.; McCabe, C.J.; Franklyn, J.A.; Kilby, M.D. The expression of thyroid hormone transporters in the human fetal cerebral cortex during early development and in N-Tera-2 neurodifferentiation. J. Physiol. 2011, 589, 2827–2845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fleming, C.E.; Mar, F.M.; Franquinho, F.; Saraiva, M.J.; Sousa, M.M. Transthyretin internalization by sensory neurons is megalin mediated and necessary for its neuritogenic activity. J. Neurosci. 2009, 29, 3220–3322. [Google Scholar] [CrossRef] [PubMed]
- James, S.R.; Franklyn, J.A.; Kilby, M.D. Placental transport of thyroid hormone. Best Pract. Res. Clin. Endocrinol. Metab. 2007, 21, 253–264. [Google Scholar] [CrossRef] [PubMed]
- Vulsma, T.; Gons, M.H.; de Vijlder, J.J. Maternal-fetal transfer of thyroxine in congenital hypothyroidism due to a total organification defect or thyroid agenesis. N. Engl. J. Med. 1989, 321, 13–16. [Google Scholar] [CrossRef] [PubMed]
- Landers, K.A.; McKinnon, B.D.; Li, H.; Subramaniam, V.N.; Mortimer, R.H.; Richard, K. Carrier-mediated thyroid hormone transport into placenta by placental transthyretin. J. Clin. Endocrinol. Metab. 2009, 94, 2610–2616. [Google Scholar] [CrossRef] [PubMed]
- Auso, E.; Lavado-Autric, R.; Cuevas, E.; del Rey, F.E.; Morreale De Escobar, G.; Berbel, P. A moderate and transient deficiency of maternal thyroid function at the beginning of fetal neocorticogenesis alters neuronal migration. Endocrinology 2004, 145, 4037–4047. [Google Scholar] [CrossRef] [PubMed]
- Oki, N.; Matsuo, H.; Nakago, S.; Murakoshi, H.; Laoag-Fernandez, J.B.; Maruo, T. Effects of 3,5,3'-triiodothyronine on the invasive potential and the expression of integrins and matrix metalloproteinases in cultured early placental extravillous trophoblasts. J. Clin. Endocrinol. Metab. 2004, 89, 5213–5221. [Google Scholar] [CrossRef] [PubMed]
- Barber, K.J.; Franklyn, J.A.; McCabe, C.J.; Khanim, F.L.; Bulmer, J.N.; Whitley, G.S.; Kilby, M.D. The in vitro effects of triiodothyronine on epidermal growth factor-induced trophoblast function. J. Clin. Endocrinol. Metab. 2005, 90, 1655–1661. [Google Scholar] [CrossRef] [PubMed]
- Larijani, B.; Marsoosi, V.; Aghakhani, S.; Moradi, A.; Hashemipour, S. Thyroid hormone alteration in pre-eclamptic women. Gynecol. Endocrinol. 2004, 18, 97–100. [Google Scholar] [CrossRef] [PubMed]
- Belet, N.; Imdat, H.; Yanik, F.; Kucukoduk, S. Thyroid function tests in preterm infants born to preeclamptic mothers with placental insufficiency. J. Pediatr. Endocrinol. Metab. 2003, 16, 1131–1135. [Google Scholar] [PubMed]
- Kurlak, L.O.; Mistry, H.D.; Kaptein, E.; Visser, T.J.; Broughton Pipkin, F. Thyroid hormones and their placental deiodination in normal and pre-eclamptic pregnancy. Placenta 2013, 34, 395–400. [Google Scholar] [CrossRef] [PubMed]
- Iwasaki, A.; Medzhitov, R. Regulation of adaptive immunity by the innate immune system. Science 2010, 327, 291–295. [Google Scholar] [CrossRef] [PubMed]
- Valimaki, E.; Miettinen, J.J.; Lietzen, N.; Matikainen, S.; Nyman, T.A. Monosodium urate activates SRC/PYK2/PI3 kinase and cathepsin dependent unconventional protein secretion from human primary macrophages. Mol. Cell. Proteomics 2013, 12, 749–763. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Evans, J.E.; Rock, K.L. Molecular identification of a danger signal that alerts the immune system to dying cells. Nature 2003, 425, 516–521. [Google Scholar] [CrossRef] [PubMed]
- Nicholson, D.W. Caspase structure, proteolytic substrates, and function during apoptotic cell death. Cell Death Differ. 1999, 6, 1028–1042. [Google Scholar] [CrossRef] [PubMed]
- Lamkanfi, M.; Declercq, W.; Kalai, M.; Saelens, X.; Vandenabeele, P. Alice in caspase land. A phylogenetic analysis of caspases from worm to man. Cell Death Differ. 2002, 9, 358–361. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.C.; Wang, L.J.; Tsang, N.M.; Ojcius, D.M.; Chen, C.C.; Ouyang, C.N.; Hsueh, C.; Liang, Y.; Chang, K.P.; Chang, Y.S. Tumour inflammasome-derived IL-1β recruits neutrophils and improves local recurrence-free survival in EBV-induced nasopharyngeal carcinoma. EMBO Mol. Med. 2012, 4, 1276–1293. [Google Scholar] [CrossRef] [PubMed]
- Lamkanfi, M.; Kanneganti, T.D.; Van Damme, P.; Vanden Berghe, T.; Vanoverberghe, I.; Vandekerckhove, J.; Vandenabeele, P.; Gevaert, K.; Nunez, G. Targeted peptidecentric proteomics reveals caspase-7 as a substrate of the caspase-1 inflammasomes. Mol. Cell. Proteomics 2008, 7, 2350–2363. [Google Scholar] [CrossRef] [PubMed]
- Brasier, A.R. The NF-κB regulatory network. Cardiovasc. Toxicol. 2006, 6, 111–130. [Google Scholar] [CrossRef]
- Pop, C.; Salvesen, G.S. Human caspases: Activation, specificity, and regulation. J. Biol. Chem. 2009, 284, 21777–21781. [Google Scholar] [CrossRef] [PubMed]
- Fuentes-Prior, P.; Salvesen, G.S. The protein structures that shape caspase activity, specificity, activation and inhibition. Biochem. J. 2004, 384, 201–232. [Google Scholar] [CrossRef] [PubMed]
- Petrilli, V.; Dostert, C.; Muruve, D.A.; Tschopp, J. The inflammasome: A danger sensing complex triggering innate immunity. Curr. Opin. Immunol. 2007, 19, 615–622. [Google Scholar] [CrossRef] [PubMed]
- Strowig, T.; Henao-Mejia, J.; Elinav, E.; Flavell, R. Inflammasomes in health and disease. Nature 2012, 481, 278–286. [Google Scholar] [CrossRef] [PubMed]
- Kanneganti, T.D.; Ozoren, N.; Body-Malapel, M.; Amer, A.; Park, J.H.; Franchi, L.; Whitfield, J.; Barchet, W.; Colonna, M.; Vandenabeele, P.; et al. Bacterial RNA and small antiviral compounds activate caspase-1 through cryopyrin/Nalp3. Nature 2006, 440, 233–236. [Google Scholar] [CrossRef] [PubMed]
- Mariathasan, S.; Weiss, D.S.; Newton, K.; McBride, J.; O'Rourke, K.; Roose-Girma, M.; Lee, W.P.; Weinrauch, Y.; Monack, D.M.; Dixit, V.M. Cryopyrin activates the inflammasome in response to toxins and ATP. Nature 2006, 440, 228–232. [Google Scholar] [CrossRef] [PubMed]
- Martinon, F.; Agostini, L.; Meylan, E.; Tschopp, J. Identification of bacterial muramyl dipeptide as activator of the NALP3/cryopyrin inflammasome. Curr. Biol. 2004, 14, 1929–1934. [Google Scholar] [CrossRef] [PubMed]
- Zhou, R.; Yazdi, A.S.; Menu, P.; Tschopp, J. A role for mitochondria in NLRP3 inflammasome activation. Nature 2011, 469, 221–225. [Google Scholar] [CrossRef] [PubMed]
- Mariathasan, S.; Monack, D.M. Inflammasome adaptors and sensors: Intracellular regulators of infection and inflammation. Nat. Rev. Immunol. 2007, 7, 31–40. [Google Scholar] [CrossRef] [PubMed]
- Agard, N.J.; Maltby, D.; Wells, J.A. Inflammatory stimuli regulate caspase substrate profiles. Mol. Cell. Proteomics 2010, 9, 880–893. [Google Scholar] [CrossRef] [PubMed]
- Abrahams, V.M. Pattern recognition at the maternal-fetal interface. Immunol. Investig. 2008, 37, 427–447. [Google Scholar] [CrossRef]
- Costello, M.J.; Joyce, S.K.; Abrahams, V.M. NOD protein expression and function in first trimester trophoblast cells. Am. J. Reprod. Immunol. 2007, 57, 67–80. [Google Scholar] [CrossRef] [PubMed]
- Mulla, M.J.; Yu, A.G.; Cardenas, I.; Guller, S.; Panda, B.; Abrahams, V.M. Regulation of Nod1 and Nod2 in first trimester trophoblast cells. Am. J. Reprod. Immunol. 2009, 61, 294–302. [Google Scholar] [CrossRef] [PubMed]
- Abrahams, V.M. The role of the Nod-like receptor family in trophoblast innate immune responses. J. Reprod. Immunol. 2011, 88, 112–117. [Google Scholar] [CrossRef] [PubMed]
- Girardin, S.E.; Tournebize, R.; Mavris, M.; Page, A.L.; Li, X.; Stark, G.R.; Bertin, J.; DiStefano, P.S.; Yaniv, M.; Sansonetti, P.J.; et al. CARD4/NOD1 mediates NF-κB and JNK activation by invasive Shigella flexneri. EMBO Rep. 2001, 2, 736–742. [Google Scholar] [CrossRef] [PubMed]
- Ogura, Y.; Inohara, N.; Benito, A.; Chen, F.F.; Yamaoka, S.; Nunez, G. Nod2, a Nod1/Apaf-1 family member that is restricted to monocytes and activates NF-κB. J. Biol. Chem. 2001, 276, 4812–4818. [Google Scholar] [CrossRef] [PubMed]
- Pettker, C.M.; Buhimschi, I.A.; Magloire, L.K.; Sfakianaki, A.K.; Hamar, B.D.; Buhimschi, C.S. Value of placental microbial evaluation in diagnosing intra-amniotic infection. Obstet. Gynecol. 2007, 109, 739–749. [Google Scholar] [CrossRef] [PubMed]
- Jiang, K.; Akhmanova, A. Microtubule tip-interacting proteins: A view from both ends. Curr. Opin. Cell Biol. 2011, 23, 94–101. [Google Scholar] [CrossRef] [PubMed]
- Wen, Y.; Eng, C.H.; Schmoranzer, J.; Cabrera-Poch, N.; Morris, E.J.; Chen, M.; Wallar, B.J.; Alberts, A.S.; Gundersen, G.G. EB1 and APC bind to mDia to stabilize microtubules downstream of Rho and promote cell migration. Nat. Cell Biol. 2004, 6, 820–830. [Google Scholar] [CrossRef] [PubMed]
- Morrison, E.E. The APC-EB1 interaction. Adv. Exp. Med. Biol. 2009, 656, 41–50. [Google Scholar] [PubMed]
- Patel, P.C.; Fisher, K.H.; Yang, E.C.; Deane, C.M.; Harrison, R.E. Proteomic analysis of microtubule-associated proteins during macrophage activation. Mol. Cell. Proteomics 2009, 8, 2500–2514. [Google Scholar] [CrossRef] [PubMed]
- Dagenais, M.; Skeldon, A.; Saleh, M. The inflammasome: In memory of Dr. Jurg Tschopp. Cell Death Differ. 2012, 19, 5–12. [Google Scholar] [CrossRef] [PubMed]
- Roberts, J.M.; Cooper, D.W. Pathogenesis and genetics of pre-eclampsia. Lancet 2001, 357, 53–56. [Google Scholar] [CrossRef] [PubMed]
- Van’t Veer, L.J.; Dai, H.; van de Vijver, M.J.; He, Y.D.; Hart, A.A.; Mao, M.; Peterse, H.L.; van der Kooy, K.; Marton, M.J.; Witteveen, A.T.; et al. Gene expression profiling predicts clinical outcome of breast cancer. Nature 2002, 415, 530–536. [Google Scholar] [CrossRef] [PubMed]
- Sibai, B.M. Diagnosis and management of gestational hypertension and preeclampsia. Obstet. Gynecol. 2003, 102, 181–192. [Google Scholar] [CrossRef] [PubMed]
- Cox, B.; Sharma, P.; Evangelou, A.I.; Whiteley, K.; Ignatchenko, V.; Ignatchenko, A.; Baczyk, D.; Czikk, M.; Kingdom, J.; Rossant, J.; et al. Translational analysis of mouse and human placental protein and mRNA reveals distinct molecular pathologies in human preeclampsia. Mol. Cell. Proteomics 2011, 10. [Google Scholar] [CrossRef] [PubMed]
- Martin, J.A.; Osterman, M.J.; Sutton, P.D. Are preterm births on the decline in the United States? In Recent data from the National Vital Statistics System; NCHS Data Brief, 2010; pp. 1–8. [Google Scholar]
- McGuire, W.; Clerihew, L.; Fowlie, P.W. FInfection in the preterm infant. BMJ 2004, 329, 1277–1280. [Google Scholar] [CrossRef] [PubMed]
- Stoll, B.J.; Hansen, N.; Fanaroff, A.A.; Wright, L.L.; Carlo, W.A.; Ehrenkranz, R.A.; Lemons, J.A.; Donovan, E.F.; Stark, A.R.; Tyson, J.E.; et al. Late-onset sepsis in very low birth weight neonates: The experience of the nichd neonatal research network. Pediatrics 2002, 110, 285–291. [Google Scholar] [CrossRef] [PubMed]
- Lamont, R.F. The role of infection in preterm labour and birth. Hosp. Med. 2003, 64, 644–647. [Google Scholar] [CrossRef] [PubMed]
- Chan, E.C.; Fraser, S.; Yin, S.; Yeo, G.; Kwek, K.; Fairclough, R.J.; Smith, R. Human myometrial genes are differentially expressed in labor: A suppression subtractive hybridization study. J. Clin. Endocrinol. Metab. 2002, 87, 2435–2441. [Google Scholar] [CrossRef] [PubMed]
- Charpigny, G.; Leroy, M.J.; Breuiller-Fouche, M.; Tanfin, Z.; Mhaouty-Kodja, S.; Robin, P.; Leiber, D.; Cohen-Tannoudji, J.; Cabrol, D.; Barberis, C.; et al. A functional genomic study to identify differential gene expression in the preterm and term human myometrium. Biol. Reprod. 2003, 68, 2289–2296. [Google Scholar] [CrossRef] [PubMed]
- Marvin, K.W.; Keelan, J.A.; Eykholt, R.L.; Sato, T.A.; Mitchell, M.D. Use of cDNA arrays to generate differential expression profiles for inflammatory genes in human gestational membranes delivered at term and preterm. Mol. Hum. Reprod. 2002, 8, 399–408. [Google Scholar] [CrossRef] [PubMed]
- Bethin, K.E.; Nagai, Y.; Sladek, R.; Asada, M.; Sadovsky, Y.; Hudson, T.J.; Muglia, L.J. Microarray analysis of uterine gene expression in mouse and human pregnancy. Mol. Endocrinol. 2003, 17, 1454–1469. [Google Scholar] [CrossRef] [PubMed]
- Young, A.; Thomson, A.J.; Ledingham, M.; Jordan, F.; Greer, I.A.; Norman, J.E. Immunolocalization of proinflammatory cytokines in myometrium, cervix, and fetal membranes during human parturition at term. Biol. Reprod. 2002, 66, 445–449. [Google Scholar] [CrossRef] [PubMed]
- Romero, R.; Yoon, B.H.; Kenney, J.S.; Gomez, R.; Allison, A.C.; Sehgal, P.B. Amniotic fluid interleukin-6 determinations are of diagnostic and prognostic value in preterm labor. Am. J. Reprod. Immunol. 1993, 30, 167–183. [Google Scholar] [CrossRef] [PubMed]
- Maymon, E.; Ghezzi, F.; Edwin, S.S.; Mazor, M.; Yoon, B.H.; Gomez, R.; Romero, R. The tumor necrosis factor α and its soluble receptor profile in term and preterm parturition. Am. J. Obstet. Gynecol. 1999, 181, 1142–1148. [Google Scholar] [CrossRef] [PubMed]
- Osman, I.; Young, A.; Ledingham, M.A.; Thomson, A.J.; Jordan, F.; Greer, I.A.; Norman, J.E. Leukocyte density and pro-inflammatory cytokine expression in human fetal membranes, decidua, cervix and myometrium before and during labour at term. Mol. Hum. Reprod. 2003, 9, 41–45. [Google Scholar] [CrossRef] [PubMed]
- Keelan, J.A.; Marvin, K.W.; Sato, T.A.; Coleman, M.; McCowan, L.M.; Mitchell, M.D. Cytokine abundance in placental tissues: Evidence of inflammatory activation in gestational membranes with term and preterm parturition. Am. J. Obstet. Gynecol. 1999, 181, 1530–1536. [Google Scholar] [CrossRef] [PubMed]
- Hayden, M.S.; Ghosh, S. Signaling to NF-κB. Genes Dev. 2004, 18, 2195–2224. [Google Scholar] [CrossRef] [PubMed]
- Schmitz, M.L.; Mattioli, I.; Buss, H.; Kracht, M. NF-κB: A multifaceted transcription factor regulated at several levels. Chembiochem 2004, 5, 1348–1358. [Google Scholar] [CrossRef] [PubMed]
- Lindstrom, T.M.; Bennett, P.R. The role of nuclear factor kappa B in human labour. Reproduction 2005, 130, 569–581. [Google Scholar] [CrossRef] [PubMed]
- Stjernholm-Vladic, Y.; Stygar, D.; Mansson, C.; Masironi, B.; Akerberg, S.; Wang, H.; Ekman-Ordeberg, G.; Sahlin, L. Factors involved in the inflammatory events of cervical ripening in humans. Reprod. Biol. Endocrinol. 2004, 2. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.; Allport, V.; Sykes, A.; Lindstrom, T.; Slater, D.; Bennett, P. The effects of labour and of interleukin 1 β upon the expression of nuclear factor kappa B related proteins in human amnion. Mol. Hum. Reprod. 2003, 9, 213–218. [Google Scholar] [CrossRef] [PubMed]
- Allport, V.C.; Pieber, D.; Slater, D.M.; Newton, R.; White, J.O.; Bennett, P.R. Human labour is associated with nuclear factor-κB activity which mediates cyclo-oxygenase-2 expression and is involved with the “functional progesterone withdrawal”. Mol. Hum. Reprod. 2001, 7, 581–586. [Google Scholar] [CrossRef] [PubMed]
- Baltimore, R.S. Neonatal sepsis: Epidemiology and management. Paediatr. Drugs 2003, 5, 723–740. [Google Scholar] [CrossRef] [PubMed]
- Malik, A.; Hui, C.P.; Pennie, R.A.; Kirpalani, H. Beyond the complete blood cell count and C-reactive protein: A systematic review of modern diagnostic tests for neonatal sepsis. Arch. Pediatr. Adolesc. Med. 2003, 157, 511–516. [Google Scholar] [CrossRef] [PubMed]
- Underwood, M.A.; Gilbert, W.M.; Sherman, M.P. Amniotic fluid: Not just fetal urine anymore. J. Perinatol. 2005, 25, 341–348. [Google Scholar] [CrossRef] [PubMed]
- Sinha, R.A.; Singh, B.K.; Yen, P.M. Thyroid hormone regulation of hepatic lipid and carbohydrate metabolism. Trends Endocrinol. Metab. 2014, 10, 538–545. [Google Scholar] [CrossRef]
- Tseng, Y.H.; Ke, P.Y.; Liao, C.J.; Wu, S.M.; Chi, H.C.; Tsai, C.Y.; Chen, C.Y.; Lin, Y.H.; Lin, K.H. Chromosome 19 open reading frame 80 is upregulated by thyroid hormone and modulates autophagy and lipid metabolism. Autophagy 2014, 10, 20–31. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.P.; Chen, Y.Y.; Huang, J.P.; Wu, Y.H. The effect of conditioned medium derived from human placental multipotent mesenchymal stromal cells on neutrophils: Possible implications for placental infection. Mol. Hum. Reprod. 2014, 20, 1117–1125. [Google Scholar] [CrossRef] [PubMed]
- Polin, R.A. The “ins and outs” of neonatal sepsis. J. Pediatr. 2003, 143, 3–4. [Google Scholar] [CrossRef] [PubMed]
- Kavathas, P.B.; Boeras, C.M.; Mulla, M.J.; Abrahams, V.M. Nod1, but not the ASC inflammasome, contributes to induction of IL-1β secretion in human trophoblasts after sensing of Chlamydia trachomatis. Mucosal Immunol. 2013, 6, 235–243. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, C.-Y.; Chen, C.-P.; Lin, K.-H. Biological Functions of Thyroid Hormone in Placenta. Int. J. Mol. Sci. 2015, 16, 4161-4179. https://doi.org/10.3390/ijms16024161
Chen C-Y, Chen C-P, Lin K-H. Biological Functions of Thyroid Hormone in Placenta. International Journal of Molecular Sciences. 2015; 16(2):4161-4179. https://doi.org/10.3390/ijms16024161
Chicago/Turabian StyleChen, Cheng-Yi, Chie-Pein Chen, and Kwang-Huei Lin. 2015. "Biological Functions of Thyroid Hormone in Placenta" International Journal of Molecular Sciences 16, no. 2: 4161-4179. https://doi.org/10.3390/ijms16024161
APA StyleChen, C. -Y., Chen, C. -P., & Lin, K. -H. (2015). Biological Functions of Thyroid Hormone in Placenta. International Journal of Molecular Sciences, 16(2), 4161-4179. https://doi.org/10.3390/ijms16024161