
Cell Death Pathways and Phthalocyanine as an Efficient Agent for Photodynamic Cancer Therapy



Abstract
:1. Introduction
1.1. Apoptosis

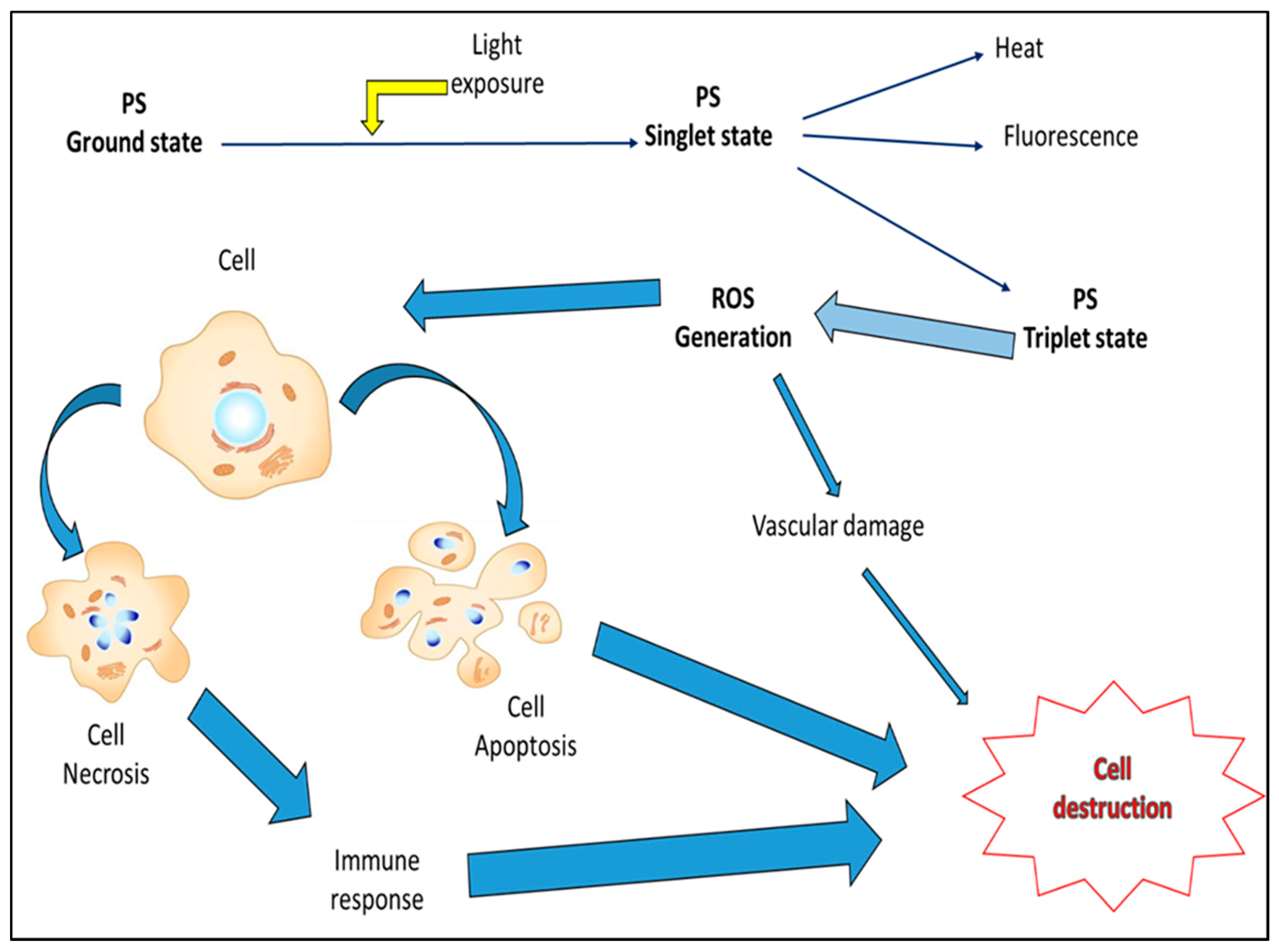{kind=link}
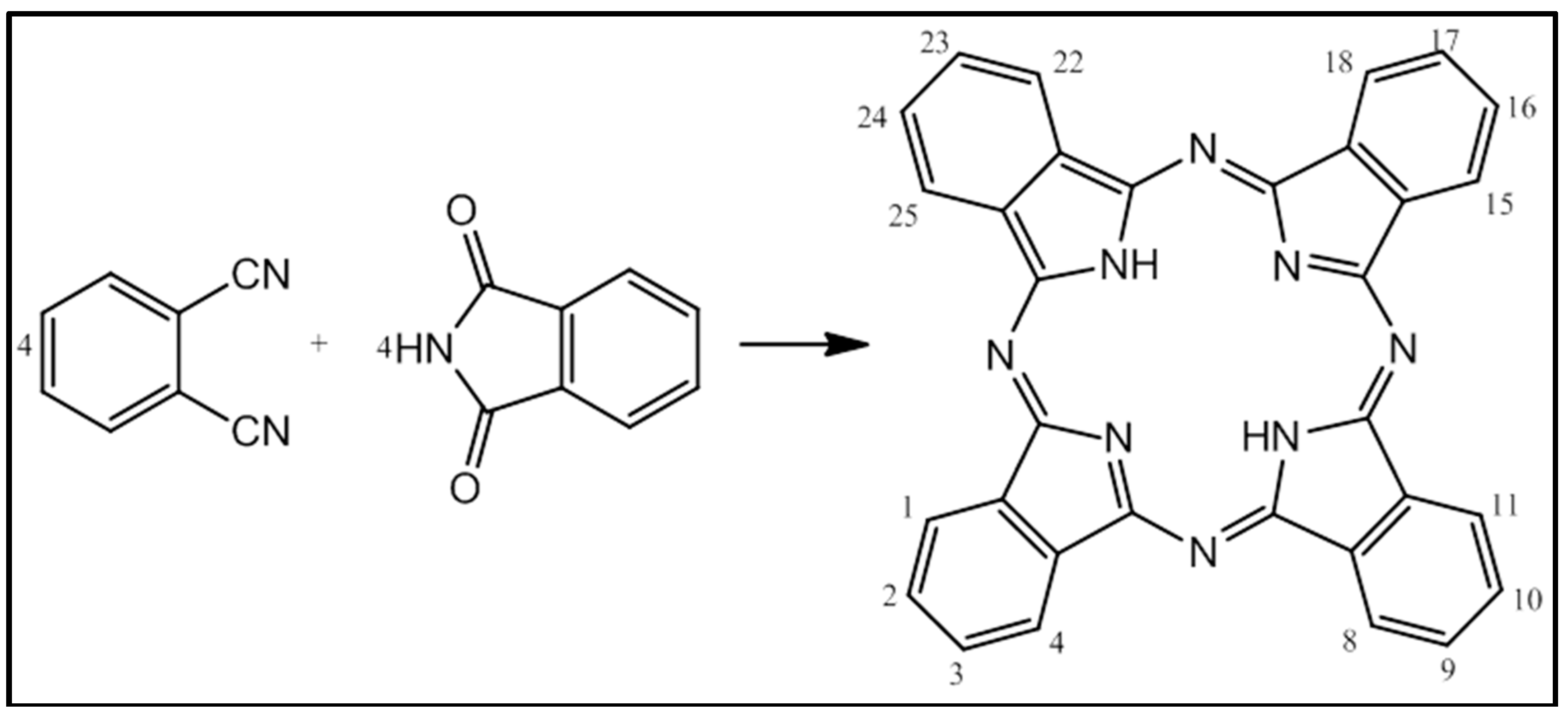{kind=link}
| Distinctive Features | Cell Death Pathways | ||
|---|---|---|---|
| Apoptosis | Autophagy | Necrosis | |
| Morphologies | Shrinkage; blebbing; chromatin condensation; DNA degradation; nuclear fragmentation, apoptotic bodies | Decreased cell size; double membrane vesicles; organelle degradation | Cell swelling; loss of membrane integrity; organelle swelling; NO DNA laddering |
| Regulators | Death receptors; Bcl-2 family; Beclin 1; caspases; IAPs; adaptor proteins; kinases; phosphatases; calcium ions, calpains; BCNI1 | mTOR; PI3 kinase; ATG family; UPR stress sensors; Beclin 1; kinase (JNK); Bcl-2 family; IP3 receptor | Calcium ions; ion channels; metabolic failure; PARB, calcium-regulated proteins; RIP kinase; death receptors; ceramides |
| Stimuli | ROS; DNA damage; death receptors ligands; developmental programs; organelle stress; anti-cancer drugs; ER calcium release | Nutrient starvation; protein aggregation; ER stress; calcium overload; developmental programs; hypoxia; ischemia; damaged organelles; proteasome impairment | bacterial toxins; metabolic poisons; ischemia; stroke; calcium overload |
| Response | Programmed, physiological | Survival, accidental, physiological | Accidental, pathological |
1.2. Autophagy
1.3. Necrosis
1.4. Reactive Oxygen Species and Photodynamic Cancer Therapy


2. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Engelberg-Kulka, H.; Amitai, S.; Kolodkin-Gal, I.; Hazan, R. Bacterial programmed cell death and multicellular behavior in bacteria. PLoS Genet. 2006, 2, e135. [Google Scholar] [CrossRef] [PubMed]
- Schultz, D.R.; Harrington, W.J., Jr. Apoptosis: Programmed cell death at a molecular level. Semin. Arthritis Rheum. 2003, 32, 345–369. [Google Scholar] [CrossRef] [PubMed]
- Yuan, J.; Kroemer, G. Alternative cell death mechanisms in development and beyond. Genes Dev. 2010, 24, 2592–2602. [Google Scholar] [CrossRef] [PubMed]
- Melino, G. The Sirens’ song. Nature 2001, 412, 23–31. [Google Scholar] [CrossRef] [PubMed]
- Kroemer, G.; Galluzzi, L.; Vandenabeele, P.; Abrams, J.; Alnemri, E.S.; Baehrecke, E.H.; Blagosklonny, M.V.; El-Deiry, W.S.; Golstein, P.; Green, D.R.; et al. Classification of cell death. Cell Death Differ. 2009, 16, 3–11. [Google Scholar] [CrossRef] [PubMed]
- Erental, A.; Sharon, I.; Engelberg-Kulka, H. Two programmed cell death systems in Escherichia coli: An apoptotic-like death is inhibited by the mazEF-mediated death pathway. PLoS Biol. 2012, 10, e1001281. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.S.; Cho, C.H.; Park, E.K.; Jung, M.H.; Yoon, K.S.; Park, H.K. AFM-Detected apoptotic changes in morphology and biophysical property caused by paclitaxel in ishikawa and heLa cells. PLoS ONE 2012, 7, e30066. [Google Scholar] [CrossRef] [PubMed]
- Kang, R.; Zeh, H.J.; Lotze, M.T.; Tang, D. The Beclin 1 network regulates autophagy and apoptosis. Cell Death Differ. 2011, 18, 571–580. [Google Scholar] [CrossRef] [PubMed]
- Lam, M; Lee, Y.J.; Deng, M.; Hsia, A.H.; Morrissey, K.A.; Yan, C.; Azzizudin, K.; Oleinick, N.L.; McCormick, T.S.; Cooper, K.D.; et al. Photodynamic therapy with the silicon phthalocyanine Pc 4 induces apoptosis inMycosis fungoides and sezary syndrome. Adv. Hematol. 2010, 2010. [Google Scholar] [CrossRef]
- Mfouo-Tynga, I.; Houreld, N.N.; Abrahamse, H. Induced cell death pathway post photodynamic therapy using a metallophthalocyanine photosensitizer in breast cancer cells. Photomed. Laser Surg. 2014, 32, 205–211. [Google Scholar] [CrossRef] [PubMed]
- Mroz, P.; Yaroslavsky, A.; Kharkwal, G.B.; Hamblin, M.R. Cell death pathways in photodynamic therapy of cancer. Cancers 2011, 3, 2516–2539. [Google Scholar] [CrossRef] [PubMed]
- Giorgi, C.; Bonora, M.; Sorrentino, G.; Missiroli, S.; Poletti, F.; Suski, J.M.; Ramirez, F.G.; Rizzuto, R.; di Virgilio, F.; Zito, E.; et al. p53 at the endoplasmic reticulum regulates apoptosis in a Ca2+-dependent manner. Proc. Natl Acad. Sci. USA 2015, 112, 1779–1784. [Google Scholar] [CrossRef] [PubMed]
- Giorgi, C.; Bonora, M.; Missiroli, S.; Poletti, F.; Ramirez, F.G.; Morciano, G.; Morganti, C.; Pandolfi, P.P.; Mammano, F.; Pinton, P. Intravital imaging reveals p53-dependent cancer cell death induced by phototherapy via calcium signaling. Oncotarget 2015, 6, 1435–1445. [Google Scholar] [PubMed]
- Tsujimoto, Y.; Shimizu, S. Another way to die: Autophagic programmed cell death. Cell Death Differ. 2005, 12, 1528–1534. [Google Scholar] [CrossRef] [PubMed]
- Jain, K.; Paranandi, K.S.; Sridharan, S.; Basu, A. Autophagy in breast cancer and its implications for therapy. Am. J. Cancer Res. 2013, 3, 251–265. [Google Scholar] [PubMed]
- Michaud, M.; Martins, I.; Sukkurwala, A.Q.; Adjemian, S.; Ma, Y.; Pellegatti, P.; Shen, S.; Kepp, O.; Scoazec, M.; Mignot, G.; et al. Autophagy-dependent anticancer immune responses induced by chemotherapeutic agents in mice. Science 2011, 334, 1573–1577. [Google Scholar] [CrossRef] [PubMed]
- Di, X.; Zhang, G.; Zhang, Y.; Takeda, K.; Rivera Rosado, L.A.; Zhang, B. Accumulation of autophagosomes in breast cancer cells induces TRAIL resistance through downregulation of surface expression of death receptors 4 and 5. Oncotarget. 2013, 4, 1349–1364. [Google Scholar] [PubMed]
- Xie, Z.; Klionsky, D.J. Autophagosome formation: Core machinery and adaptation. Nat. Cell Biol. 2007, 9, 1102–1109. [Google Scholar] [CrossRef] [PubMed]
- Nikoletopoulou, V.; Markaki, M.; Palikaras, K.; Tavernarakis, N. Crosstalk between apoptosis, necrosis and autophagy. Biochem. Biophys. Acta 2013, 1833, 3448–3459. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, L.; Shi, Z.; Zhao, S.; Wang, F.T.; Zhou, T.T.; Liu, B.; Bao, J.K. Programmed cell death pathways in cancer: A review of apoptosis, autophagy and programmed necrosis. Cell Prolif. 2012, 45, 487–498. [Google Scholar] [CrossRef] [PubMed]
- Lamkanfi, M.; Festjens, N.; Declercq, W.; Vanden Berghe, T.; Vandenabeele, P. Caspases in cell survival, proliferation and differentiation. Cell Death Differ. 2007, 14, 44–55. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.; Wan, F.; Dutta, S.; Welsh, S.; Liu, Z.; Freundt, E.; Baehrecke, E.H.; Lenardo, M. Autophagic programmed cell death by selective catalase degradation. Proc. Natl. Acad. Sci. USA 2006, 103, 4952–4957. [Google Scholar] [CrossRef] [PubMed]
- Mizushima, N.; Klionsky, D.J. Protein turnover via autophagy: Implications for metabolism. Annu. Rev. Nutr. 2007, 27, 19–40. [Google Scholar] [CrossRef] [PubMed]
- Separovic, D.; Joseph, N.; Breen, P.; Bielawski, J.; Pierce, J.S.; van Buren, E.; Bhatti, G.; Saad, Z.H.; Bai, A.; Bielawska, A. Combining anticancer agents photodynamic therapy and LCL85 leads to distinct changes in the sphingolipid profile, autophagy, caspase-3 activation in the absence of cell death, and long-term sensitization. Biochem. Biophys. Res. Commun. 2011, 409, 372–377. [Google Scholar] [CrossRef] [PubMed]
- Patel, A.S.; Lin, L.; Geyer, A.; Haspel, J.A.; An, C.H.; J. Cao, J.; Rosas, I.O.; Morse, D. Autophagy in Idiopathic Pulmonary Fibrosis. PLoS ONE 2012, 7, e41394. [Google Scholar] [CrossRef]
- Peracchio, C.; Alabiso, O.; Valente, G.; Isidoro, C. Involvement of autophagy in ovarian cancer: A working hypothesis. J. Ovarian Res. 2012, 5, 22–32. [Google Scholar] [CrossRef] [PubMed]
- Xue, L.Y.; Chiu, S.M.; Oleinick, N.L. Atg7 deficiency increases resistance of MCF-7 human breast cancer cells to photodynamic therapy. Autophagy 2010, 6, 248–255. [Google Scholar] [CrossRef] [PubMed]
- Garg, A.D.; Maes, H.; Romanoa, E.; Agostinis, P. Autophagy, a major adaptation pathway shaping cancer cell death and anticancer immunity responses following photodynamic therapy. Photochem. Photobiol. Sci. 2015. [Google Scholar] [CrossRef]
- Lemasters, J.J. Dying a thousand deaths: Redundant pathways from different organelles to apoptosis and necrosis. Gastroentrology 2005, 129, 351–360. [Google Scholar] [CrossRef]
- Cho, Y.; McQuade, T.; Zhang, H.; Zhang, J.; Chan, F.K.M. RIP1-Dependent and independent effects of necrostatin-1 in necrosis and T cell activation. PLoS ONE 2011, 6, e23209. [Google Scholar] [CrossRef] [PubMed]
- Christofferson, D.E.; Li, Y.; Hitomi, J.; Zhou, W.; Upperman, C.; Zhu, H.; Gerber, S.A.; Gygi, S.; Yuan, J. A novel role for RIP1 kinase in mediating TNFα production. Cell Death Dis. 2012, 3. doi:1038/cdd15.2012.64. [Google Scholar]
- Longo, J.P.F.; Lozzi, S.P.; Simioni, A.R.; Morais, P.C.; Tedesco, A.C.; Azevedo, R.B. Photodynamic therapy with aluminum-chloro-phthalocyanine induces necrosis and vascular damage in mice tongue tumors. J. Photochem. Photobiol. B 2009, 94, 143–146. [Google Scholar] [CrossRef]
- Longo, J.P.F.; de Melo, L.N.D.; Mijan, M.C.; Valois, C.R.A.; Joanitti, G.A.; Simioni, A.R.; Tedesco, A.C.; de Azevedo, R.B. Photodynamic therapy mediated by liposomal chloroaluminum-phthalocyanine induces necrosis in oral cancer cells. J. Biomater. Tissue Eng. 2013, 3, 148–156. [Google Scholar] [CrossRef]
- Portilho, F.A.; Cavalcanti, C.E.O.; Miranda-Vilela, A.L.; Estevanato, L.L.C.; Longo, J.P.F.; Santos, M.F.M.A.; Bocca, A.L.; Martins, O.P.; Simioni, A.R.; Morais, P.C.; et al. Antitumor activity of photodynamic therapy performed with nanospheres containing zinc-phthalocyanine. J. Nanobiotechnol. 2013, 11, 41–65. [Google Scholar] [CrossRef] [Green Version]
- Wood, S.R.; Holroyd, J.A.; Brown, S.B. The subcellular localization of Zn (II) phthalocyanines and their redistribution on exposure to light. Photochem. Photobiol. 1997, 65, 397–402. [Google Scholar] [CrossRef] [PubMed]
- Rada, B.; Leto, T.L. Oxidative innate immune defenses by Nox/Duox family NADPH oxidases. Contrib. Microbiol. 2008, 15, 164–187. [Google Scholar] [PubMed]
- Arwert, E.N.; Hoste, E.; Watt, F.W. Epithelial stem cells, wound healing and cancer. Nat. Rev. Cancer 2012, 12, 170–180. [Google Scholar] [CrossRef] [PubMed]
- Jung, S.N.; Yang, W.K.; Kim, J.; Kim, H.S.; Kim, E.J.; Yun, H.; Park, H.; Kim, S.S.; Choe, W.; Kang, I.; et al. Reactive oxygen species stabilize hypoxia-inducible factor-1 alpha protein and stimulate transcriptional activity via AMP-activated protein kinase in DU145 human prostate cancer cells. Carcinogenesis 2008, 29, 713–721. [Google Scholar] [CrossRef] [PubMed]
- Serra, A.; Pineiro, M.; Pereira, N.; Gonsalves, A.R.; Laranjo, M.; Abrantes, M. A look at clinical applications and developments of photodynamic therapy. Oncol. Rev. 2008, 2, 235–249. [Google Scholar] [CrossRef]
- Brown, S.B.; Brown, E.A.; Walker, I. The present and future role of photodynamic therapy in cancer treatment. Lancet Oncol. 2004, 5, 497–508. [Google Scholar] [CrossRef] [PubMed]
- Kudinova, N.V.; Berezov, T.T. Photodynamic therapy: Search for ideal photosensitizer. (in Russian). Biomed. Khim. 2009, 55, 558–569. [Google Scholar] [PubMed]
- Dougherty, T.J.; Gomer, C.J.; Henderson, B.W.; Jori, G.; Kessel, D.; Korbelik, M.; Moan, J.; Peng, Q. Photodynamic therapy. J. Natl. Cancer Inst. 1998, 90, 889–905. [Google Scholar] [CrossRef] [PubMed]
- Manoto, S.L.; Abrahamse, H. Effect of a newly synthesized Zn sulfophthalocyanine derivative on cell morphology, viability, proliferation, and cytotoxicity in a human lung cancer cell line (A549). Lasers Med. Sci. 2011, 26, 523–530. [Google Scholar] [CrossRef] [PubMed]
- Mroz, P.; Szekalska, A.; Wu, M.X.; Hamblin, M.R. Photodynamic therapy of tumors can lead to development of system antigen specific immune response. PLoS ONE 2010, 5, e15194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agostinis, P.; Berg, K.; Cengel, K.A.; Foster, T.H.; Girotti, A.W.; Gollnick, S.O.; Hahn, S.M.; Hamblin, M.R.; Juzeniene, A.; Kessel, D.; et al. Photodynamic therapy of cancer: An update CA: A cancer. J. Clin. 2011, 61, 250–281. [Google Scholar] [CrossRef]
- Ahn, J.C.; Biswas, R.; Moon, J.H.; Chung, P.S. Cellular uptake of 9-hydroxypheophorbide-α and its photoactivation to induce ER stress-related apoptosis in human cervical cancer cells. Lasers Med. Sci. 2014, 29, 289–299. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, C.S.; Turchiello, R.; Kowaltowski, A.J.; Indig, G.L.; Baptista, M.S. Major determinants of photoinduced cell death: Subcellular localization versus photosensitization efficiency. Free Radic. Biol. Med. 2011, 51, 824–833. [Google Scholar] [CrossRef] [PubMed]
- Firdous, S.; Nawaz, M.; Ikram, M.; Ahmed, M. In vitro study of cell death with 5-aminolevulinic acid based photodynamic therapy to improve the efficiency of cancer treatment. Laser Phys. 2012, 22, 626–633. [Google Scholar] [CrossRef]
- Brackett, C.; Gollnick, S. Photodynamic therapy enhancement of anti-tumour immunity. Photochem. Photobiol. 2011, 83, 1063–1068. [Google Scholar]
- Pereira, P.M.R.; Silva, S.; Cavaleiro, J.A.S.; Ribeiro, C.A.F.; Tome, J.P.C.; Fernandes, R. Galactodendritic phthalocyanine targets carbohydrate-binding proteins enhancing photodynamic therapy. PLoS ONE 2014, 9, e95529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dent, C.E.; Linstead, P.R.; Lowe, A.R. Phthalocyanines. Part VI. The structure of the phthalocyanines. J. Chem. Soc. 1934, 1033–1039. [Google Scholar] [CrossRef]
- Guillaud, G.; Simon, J.; Germain, J.P. Metallophthalocyanines: Gas sensors, resistors and field effect transistors. Coord. Chem. Rev. 1998, 178–180, 1433–1484. [Google Scholar] [CrossRef]
- Allen, C.M.; Sharman, W.M.; van Lier, J.E. Current status of phthalocyanines in the photodynamic therapy of cancer. J. Porphyr. Phthalocyanines 2001, 5, 161–169. [Google Scholar] [CrossRef]
- Coupienne, I.; Fettweis, G.; Piette, J. RIP3 expression induces a death profile change in U2OS osteosarcoma cells after 5-ALA-PDT. Lasers Surg. Med. 2011, 43, 557–564. [Google Scholar] [PubMed]
- Coupienne, I.; Fettweis, G.; Rubio, N.; Agostinis, P.; Piette, J. 5-ALA-PDT induces RIP3-dependent necrosis in glioblastoma. Photochem. Photobiol. Sci. 2011, 10, 1868–1878. [Google Scholar] [CrossRef] [PubMed]
- Scheer, H.; Inhoffen, H.H. In The Porphyrins; Dolphin, D., Ed.; Academic Press: New York, NY, USA, 1978; Volume 2, pp. 45–62. [Google Scholar]
- Sternberg, E.D.; Dolphin, D. Porphyrin-based photosensitizers for use in photodynamic therapy. Tetrahedron 1988, 54, 4151–4202. [Google Scholar] [CrossRef]
- Barrett, P.A.; Dent, C.E.; Linstead, R.P. Phthalocyanines. Part VII. Phthalocyanine as a co-ordinating group. A general investigation of the metallic derivatives. J. Chem. Soc. 1934, 179, 1718–1736. [Google Scholar]
- Cid, J.-J.; Yum, J.-H.; Jang, S.-R.; Nazeeruddin, M.K.; Martinez-Ferrero, E.; Palomares, E.; Ko, J.; Graetzel, M.; Torres, T. Molecular cosensitization for efficient panchromatic dye-sensitized solar cells. Angew. Chem. Int. Ed. 2007, 46, 8358–8362. [Google Scholar] [CrossRef]
- Kadish, K.M.; Smith, K.M.; Guilard, R. The Porphyrin Handbook; Academic Press: San Diego, CA, USA, 2003. [Google Scholar]
- Pinzon, J.R.; Plonska-Brzezinska, M.E.; Cadona, C.M.; Athans, A.J.; Gayathri, S.S.; Guldi, D.M.; Herranz, M.A.; Martin, N.; Torres, T.; Echegoyen, L. Sc3N@C80-ferrocene electrondonor/acceptor conjugates as promising materials for photovoltalic applications. Angew. Chem. Int. Ed. 2008, 47, 4173–4176. [Google Scholar] [CrossRef]
- Campidelli, S.; Ballesteros, B.; Filoramo, A.; Diaz, D.; de la Torre, G.; Torres, T.; Rahman, G.M.A.; Aminur, E.C.; Kiessling, D.; Werner, F.; et al. Facile decoration of functionalized single-wall carbon nanotubes with phthalocyanines via “Click Chemistry”. J. Am. Chem. Soc. 2008, 130, 11503–11509. [Google Scholar] [CrossRef] [PubMed]
- Whitacre, C.M.; Feyes, D.K.; Satoh, T.; Grossmann, J.; Mulvihill, J.W.; Mukhtar, H.; Oleinick, N.L. Photodynamic therapy with the phthalocyanine photosensitizer Pc 4 of SW480 human colon cancer xenografts in athymic mice. Clin. Cancer Res. 2000, 6, 2021–2027. [Google Scholar] [PubMed]
- Velloso, N.V.; Muehlmann, L.A.; Longo, J.P.F.; Silva, J.R.D.; Zancanela, D.C.; Tedesco, A.C.; Azevedo, R.B.D. Aluminum-phthalocyanine chloride-based photodynamic therapy inhibits PI3K/Akt/Mtor pathway in oral squamous cell carcinoma cells in vitro. Chemo 2012, 1, 107–111. [Google Scholar]
- Staicu, A.; Pascu, A.; Nuta, A.; Sorescu, A.; Raditoiu, V.; Pascu, M.L. Studies about phthalocyanine photosensitizers to be used in photodynamic therapy. Romanian Rep. Phys. 2013, 65, 1032–1051. [Google Scholar]
- Manoto, S.L.; Sekhejane, P.R.; Houreld, N.N.; Abrahamse, H. Localization and phototoxic effect of zinc sulfophthalocyanine photosensitizer in human colon (DLD-1) and lung (A549) carcinoma cells (in vitro). Photodiagn. Photodyn. Ther. 2012, 9, 52–59. [Google Scholar] [CrossRef]
- Mfouo-Tynga, I.; Houreld, N.N.; Abrahamse, H. The primary subcellular localization of Zinc phthalocyanine and its cellular impact on viability, proliferation and structure of breast cancer cells (MCF-7). J. Photochem. Photobiol. B 2012, 120, 171–176. [Google Scholar] [CrossRef] [PubMed]
- Manoto, S.L.; Houreld, N.N.; Abrahamse, H. Phototoxic effect of photodynamic therapy on lung cancer cells grown as a monolayer and three dimensional multicellular spheroids. Lasers Surg. Med. 2013, 45, 186–194. [Google Scholar] [CrossRef] [PubMed]
- Abrahamse, H.; Kresfelder, T.; Horne, T.; Cronje, M.; Nyokong, T. Apoptotic inducing ability of a novel photosensitizing agent, Ge sulfophthalocyanine, on oesophageal and breast cancer cell lines. In Optical Methods for Tumor Treatment and Detection: Mechanisms and Techniques in Photodynamic Therapy XV. In Proceedings of the SPIE, San Jose, CA, USA, 21 January 2006; Kessel, D., Ed.; SPIE: Bellingham, WA, USA, 2006; p. 613904. [Google Scholar] [CrossRef]
- Seotsanyana-Mokhosi, I.; Kresfelder, T.; Abrahamse, H.; Nyokong, T. The effect of Ge, Si and Sn phthalocyanine photosensitizers on cell proliferation and viability of human oesophageal carcinoma cells. J. Photochem. Photobiol. B 2006, 83, 55–62. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mfouo-Tynga, I.; Abrahamse, H. Cell Death Pathways and Phthalocyanine as an Efficient Agent for Photodynamic Cancer Therapy. Int. J. Mol. Sci. 2015, 16, 10228-10241. https://doi.org/10.3390/ijms160510228
Mfouo-Tynga I, Abrahamse H. Cell Death Pathways and Phthalocyanine as an Efficient Agent for Photodynamic Cancer Therapy. International Journal of Molecular Sciences. 2015; 16(5):10228-10241. https://doi.org/10.3390/ijms160510228
Chicago/Turabian StyleMfouo-Tynga, Ivan, and Heidi Abrahamse. 2015. "Cell Death Pathways and Phthalocyanine as an Efficient Agent for Photodynamic Cancer Therapy" International Journal of Molecular Sciences 16, no. 5: 10228-10241. https://doi.org/10.3390/ijms160510228
APA StyleMfouo-Tynga, I., & Abrahamse, H. (2015). Cell Death Pathways and Phthalocyanine as an Efficient Agent for Photodynamic Cancer Therapy. International Journal of Molecular Sciences, 16(5), 10228-10241. https://doi.org/10.3390/ijms160510228