
Homogeneous Nature of Malaysian Marine Fish Epinephelus fuscoguttatus (Perciformes; Serranidae): Evidence Based on Molecular Markers, Morphology and Fourier Transform Infrared Analysis

 ,
,
Abstract
:1. Introduction
2. Results
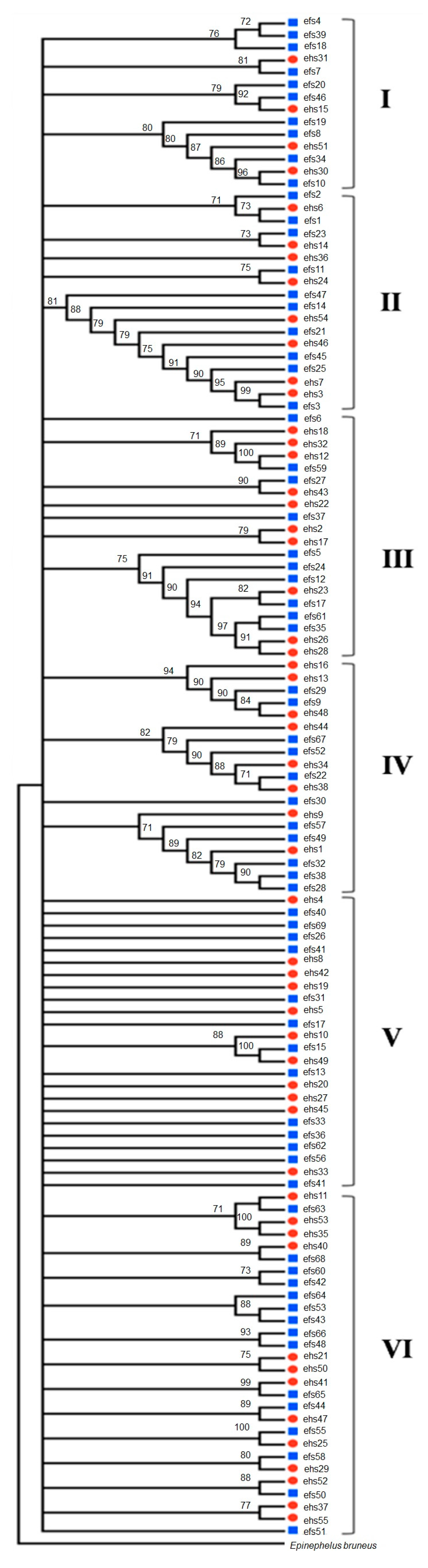
2.1. Phylogenetic Trees

2.2. Morphological Analysis


{kind=link}
{kind=link}
{kind=link}
| Characters | Description |
|---|---|
| 11 Morphometrics | |
| Total length (TL) | Tip of the lower jaw to the end of caudal fin |
| Fork length (FL) | Tip of the upper jaw to the tail base |
| Standard length (SL) | Tip of the upper jaw to the end of caudal fin |
| Head length (HL) | From the front of the upper lip to the posterior end of the opercular |
| Caudal fin length (CFL) | From tail base to tip of the caudal fin |
| Dorsal fin length (DFL) | Front of the upper lip to the origin of the dorsal fin |
| Pectoral fin length (PFL) | From base to tip of the pectoral fin |
| Anal fin length (AFL) | Front of the upper lip to the origin of the anal fin |
| Mouth length (ML) | Straight line measurement between the snout tip and posterior |
| Snout length (SnL) | The front of the upper lip to the flesh anterior edge of the orbit |
| Body width length (BwL) | The greatest width just posterior to the gill opening |
| 6 Meristic | |
| Pelvic fin ray | Number of soft fin rays in the pelvic fin |
| 1st dorsal fin ray | Number of soft fin rays in 1st dorsal fin |
| 2nd dorsal fin ray | Number of soft fin rays in 2nd dorsal fin |
| Anal fin ray | Number of soft fin rays in anal fin Caudal fin ray |
| Anal fin ray | Number of soft fin rays in anal fin Caudal fin ray |
| Pectoral fin ray | Number of soft fin rays in pectoral fin |
| Morphometrics | Meristics | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Measurements (cm) | Min | Max/Mean | Mean ± SD | TL (%) | Measurement | Range | Mean ± SD | ||||||
| EF | EH | EF | EH | EF | EH | EF | EH | EF | EH | EF | EH | ||
| Total length (TL) | 58 | 58 | 61 | 59 | 59.5 ± 1.13 | 58.5 ± 1.21 | |||||||
| Fork length (FL) | 50.5 | 49.4 | 53.7 | 53.3 | 52.3 ± 1.31 | 51.4 ± 0.89 | 87.8 | 87.8 | Pelvic fin ray | 6 to 7 | 6 to 7 | 6.5 ± 0.53 | 6.5 ± 0.5 |
| Standard length (SL) | 48.3 | 45.2 | 51.7 | 52 | 50.1 ± 1.18 | 48.6 ± 1.03 | 84 | 83.1 | 1st dorsal fin ray | 11 to 12 | 11 to 12 | 11.6 ± 0.52 | 11.5 ± 0.5 |
| Head length (HL) | 14 | 14 | 15.5 | 14.8 | 14.8 ± 0.47 | 14.4 ± 0.58 | 24.9 | 24.6 | 2nd dorsal fin ray | 9 to 13 | 9 to 14 | 11.1 ± 1.73 | 11.5 ± 1.78 |
| Caudal fin length (CFL) | 5.7 | 5.5 | 6.4 | 7.1 | 6.07 ± 0.25 | 6.3 ± 0.31 | 10.2 | 10.8 | Anal fin ray | 7 to 9 | 7 to 8 | 8 ± 0.82 | 7.5 ± 0.73 |
| Dorsal fin length (DFL) | 13.2 | 14.2 | 16.4 | 15.8 | 15.0 ± 1.13 | 15 ± 1.03 | 25.3 | 25.6 | Caudal fin ray | 14 to 15 | 14 to 15 | 14.5 ± 0.53 | 14.5 ± 0.42 |
| Pectoral fin length (PFL) | 8.2 | 8.2 | 9.5 | 9.5 | 8.85 ± 0.43 | 8.85 ± 0.4 | 14.9 | 15.1 | Pectoral fin ray | 18 to 20 | 17 to 20 | 18.8 ± 0.92 | 18.5 ± 1.13 |
| Anal fin length (AFL) | 7.9 | 7.5 | 11.3 | 10.8 | 9.62 ± 1.21 | 9.15 ± 1.33 | 16.1 | 15.6 | |||||
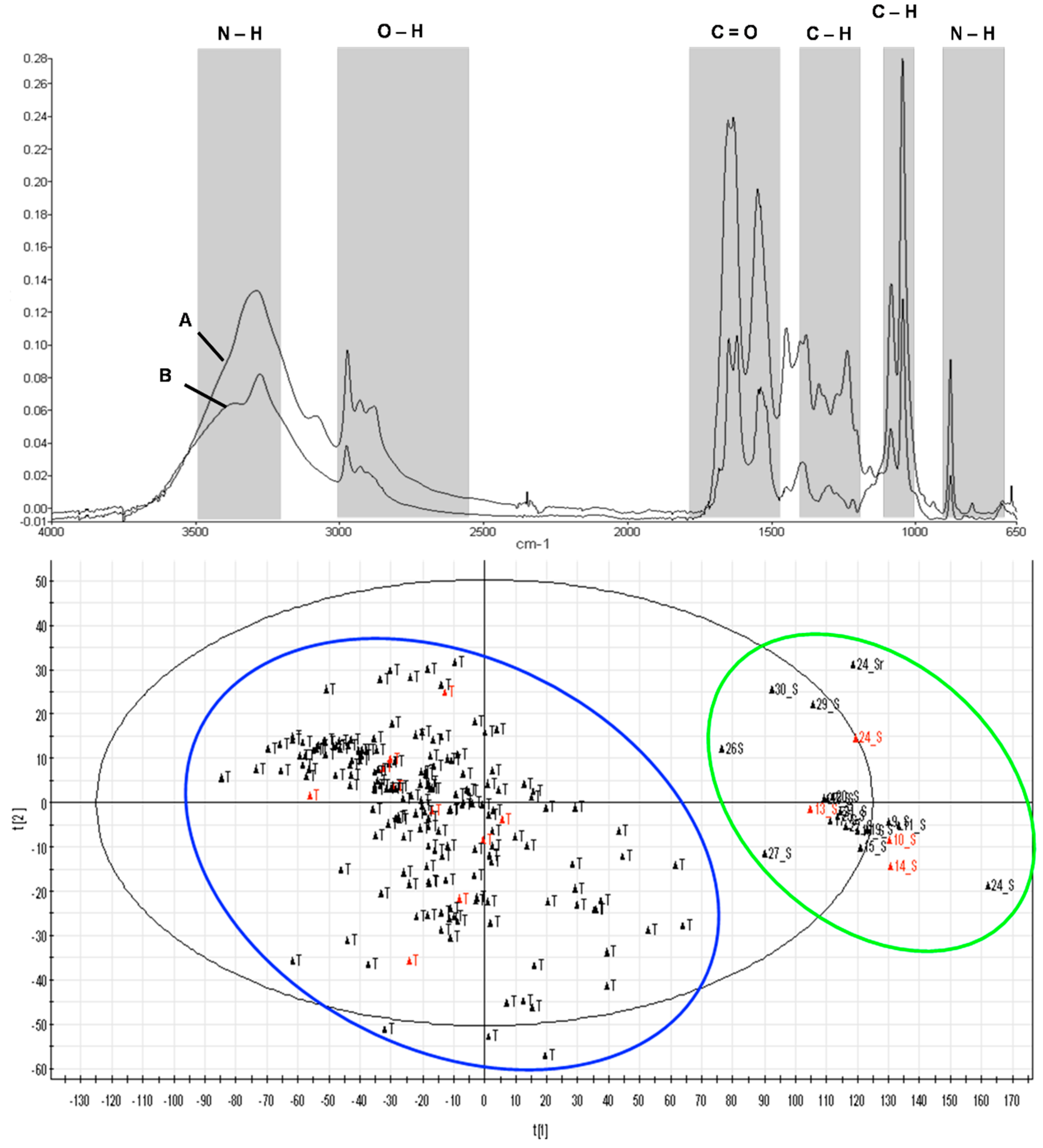
2.3. Attenuated Total Reflectance-Fourier Transform Infrared Analysis (ATR-FTIR)

| Functional Group | Spectrum Region (cm−1) | |
|---|---|---|
| Amines | N–H Stretch | 3500–3300 |
| Carboxylic acids | O–H Stretch | 3000–2500 |
| Carboxylic acids | C=O Stretch | 1700–1500 |
| Alkenes | C–H Bend | 1430–1290 |
| Alcohols | C–O | 1260–1000 |
| Amines | N–H Bend | ~800 |
3. Discussion
3.1. Molecular Analysis
3.2. Morphological Analysis
3.3. Attenuated Total Reflectance-Fourier Transform Infrared Analysis (ATR-FTIR)
4. Experimental Section
4.1. Collection and Preservation
4.2. DNA Extraction
4.3. Amplifying and Sequencing
4.4. Data Analysis
4.5. Morphological Analysis
4.6. Attenuated Total Reflectance-Fourier Transform Infrared Analysis (ATR-FTIR)
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- James, C.M.; Al-Thobaiti, S.A.; Rasem, B.M.; Carlos, M.H. Potential of Grouper Hybrid Epinephelus fuscoguttatus Х E. polyphekadion for Aquaculture Naga. ICLARM Q. 1999, 22, 19–23. [Google Scholar]
- Hseu, J.R.; Huang, W.I.; Yeong, T.C. What causes cannibalization-associated suffocation in cultured brown-marbled grouper, Epinephelus fuscoguttatus (Forsskal, 1775)? Aquac. Res. 2007, 38, 1056–1060. [Google Scholar] [CrossRef]
- Yeh, S.L.; Kuo, C.M.; Ting, Y.Y.; Chang, C.F. Androgens stimulate sex change in protogynous grouper, Epinephelus coioides: spawning performance in sex-changed males. Comp. Biochem. Physiol. C 2003, 135, 375–382. [Google Scholar] [CrossRef]
- Kuo, H.-C.; Hsu, H.-H.; Chua, C.S.; Wang, T.-Y.; Chen, Y.-M.; Chen, T.-Y. Development of pedigree classification using microsatellite and mitochondrial markers for giant grouper broodstock (Epinephelus lanceolatus) management in Taiwan. Mar. Drugs 2014, 12, 2397–2407. [Google Scholar] [CrossRef] [PubMed]
- Nedwman, S.J.; Cappo, D.; Williams, M.B. Age, growth, mortality rates and corresponding yield estimates using otoliths of the tropical red snappers, Lutjanus erythropterus, L. malabaricus and L. sebae, from the central Great Barrier Reef. Fish. Res. 2000, 48, 1–14. [Google Scholar] [CrossRef]
- Craig, M.T.; Pondella, D.J.; Franck, J.P.C.; Hafner, J.C. On the status of the serranid fish genus Epinephelus: Evidence for paraphyly based upon 16S rDNA sequence. Mol. Phylogenet. Evol. 2001, 19, 121–130. [Google Scholar] [CrossRef] [PubMed]
- Maggio, T.; Andaloro, F.; Hemida, F.; Arculeo, M. A molecular analysis of some Eastern Atlantic grouper from the Epinephelus and Mycteroperca genus. J. Exp. Mar. Biol. Ecol. 2005, 321, 83–92. [Google Scholar] [CrossRef]
- Zhu, Z.Y.; Yue, G.H. The complete mitochondrial genome of red grouper Plectropomus leopardus and its applications in identification of grouper species. Aquaculture 2008, 276, 44–49. [Google Scholar] [CrossRef]
- Craig, M.T.; Hastings, P.A. A molecular phylogeny of the groupers of the subfamily Epinephelinae (Serranidae) with a revised classification of the Epinephelini. Ichthyol. Res. 2007, 54, 1–17. [Google Scholar] [CrossRef]
- Baharum, S.N.; Nurdalila, A.A. Phylogenetic Relationships of Epinephelus fuscoguttatus and Epinephelus hexagonatus Inferred from Mitochondrial Cytochrome b Gene Sequences using Bioinformatic Tools. Int. J. Biosci. Biochem. Bioinform. 2011, 1, 47–52. [Google Scholar] [CrossRef]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef]
- Chisholm, L.A.; Morgan, J.A.T.; Adlard, R.D.; Whittington, I.D. Phylogenetic analysis of the Monocotylidae (Monogenea) inferred from 28S rDNA sequences. Int. J. Parasitol. 2001, 31, 1537–1547. [Google Scholar] [CrossRef]
- Meyer, A. Phylogenetic relationship and evolutionary processes in east African cichlid fishes. Trends Ecol. Evol. 1993, 8, 279–284. [Google Scholar] [CrossRef]
- De-La, H.R.; Ruiz, R.C.; Ruiz, R.M.; Garrido-Ramos, M.A. The molecular phylogeny of the Sparidae species (Pieces, Perciformes) based on two satellite DNA families. Heredity 2001, 87, 691–697. [Google Scholar]
- Birstein, V.J.; Desalle, R. Molecular phylogeny of Acipenserinae. Mol. Phylogenet. Evol. 1998, 9, 141–155. [Google Scholar] [CrossRef] [PubMed]
- Turan, C. Stock identification of Mediterranean horse mackerel (Trachurus mediterraneus) using morphometric and meristic characters. ICES J. Mar. Sci. 2004, 61, 774–781. [Google Scholar] [CrossRef]
- Meyer, X. Morphometrics and allometry in the trophically polymorphic cichlid fish, Cichlasoma citrinellum: Alternative adaptations and ontogenic changes in shape. J. Zool. 1990, 221, 237–260. [Google Scholar] [CrossRef]
- Kuhm, A.E.; Suter, D.; Felleisen, R.; Rau, J. Application of Fourier transform infrared spectroscopy (FT-IR) for the identification of Yersinia enterocolitica on species and subspecies level. Appl. Environ. Microbiol. 2009, 75, 5809–5813. [Google Scholar] [CrossRef] [PubMed]
- Khairudin, K.; Sukiran, N.A.; Goh, H.-H.; Baharum, S.N.; Noor, M.N. Direct discrimination of different plant populations and study on temperature effects by Fourier transform infrared spectroscopy. Metabolomics 2014, 10, 203–211. [Google Scholar] [CrossRef]
- Jagadeesan, G.; Kavitha, A.V.; Subasini, J. FTIR study of the influence of Tribulus terrestris on mercury intoxicated mice, Mus musculus, liver. Trop. Biomed. 2005, 22, 15–22. [Google Scholar] [PubMed]
- Chen, L.; Carpita, N.C.; Reiter, W.D.; Wilson, R.H.; Jeffries, C.; McCann, M.C. A rapid method to screen for cell-wall mutants using discriminant analysis of Fourier transformation infrared spectra. Plant J. 1998, 16, 385–392. [Google Scholar] [CrossRef] [PubMed]
- Wilson, R.H.; Slack, P.T.; Appleton, G.P.; Sun, L.; Belton, P.S. Determination of the fruit content of jam using Fourier transform infrared spectroscopy. Food Chem. 1993, 47, 303–308. [Google Scholar] [CrossRef]
- Rodriguez-Saona, L.E.; Khambaty, F.; Fry, F.; Dubois, J.; Calvey, E.M. Detection and identification of bacteria in a juice matrix with Fourier transform-near infrared spectroscopy and multivariate analysis. J. Food Prot. 2004, 67, 2555–2559. [Google Scholar] [PubMed]
- Evans, P.A. Differentiating “hard” from “soft” woods using Fourier transform infrared and Fourier transform spectroscopy. Spectrochim. Acta 1991, 47, 1441–1447. [Google Scholar] [CrossRef]
- Müller, G.; Schöpper, C.; Vos, H.; Kharazipour, A.; Polle, A. FTIR-ATR spectroscopic analysis of chages in wood properties during particle- and fibreboard production of hard- and softwood trees. Bioresources 2009, 4, 49–71. [Google Scholar]
- Smith, A.R.; Johnson, H.E.; Hall, M. Metabolic Fingerprinting of Salt-Stressed Tomatoes. Bulg. J. Plant Physiol. 2003, 153–163. Available online: http://www.bio21.bas.bg/ippg/bg/wp-content/uploads/2011/06/03_essa_153-163.pdf (accessed on 25 June 2015). [Google Scholar]
- Espinoza, E.O.; Baker, B.W.; Moores, T.D.; Voin, D. Forensic identification of elephant and giraffe hair artifacts using HATR FTIR spectroscopy and discriminant analysis. Endang. Spec. Res. 2008, 9, 239–246. [Google Scholar] [CrossRef]
- Boskey, A.; Camacho, N.P. FT-IR imaging of native and tissue-engineered bone and cartilage. Biomaterials 2007, 28, 2465–2478. [Google Scholar] [CrossRef] [PubMed]
- Heemstra, P.C.; Randall, J.E. Grouper of the world. FAO Fish. Synop. 1993, 125, 104–276. [Google Scholar]
- Young, N.D. A cautiously optimistic vision for marker-assisted breeding. Mol. Breed. 1999, 5, 505–510. [Google Scholar] [CrossRef]
- Rozen, S.; Skaletsky, H.J. Primer3 on the WWW for General Users and for Biologist Programmers. In Bioinformatics Methods and Protocols: Methods in Molecular Biology; Misener, S., Krawetz, S.A., Eds.; Humana Press Inc.: Totowa, NJ, USA, 2000; pp. 365–386. [Google Scholar]
- Subramaniam, S. The Biology Workbench-a seamless database and analysis environment for the biologist. Proteins 1998, 32, 1–2. [Google Scholar] [CrossRef]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [PubMed]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Dudley, J.; Nei, M.; Kumar, S. MEGA4: Molecular Evolutionary Genetics Analysis (MEGA) software version 4.0. Mol. Biol. Evol. 2007, 24, 1596–1599. [Google Scholar] [CrossRef] [PubMed]
- Nei, M.; Kumar, S. Molecular Evolution and Phylogenetics; Oxford University Press: New York, NY, USA, 2000. [Google Scholar]
- Librado, P.; Rozas, J. DnaSP v5: A software for comprehensive analysis of DNA polymorphism data. Bioinformatics 2009, 25, 1451–1452. [Google Scholar] [CrossRef] [PubMed]
- Excoffier, L. Computational and Molecular Population Genetics Lab CMPG. 2010. Available online: http://cmpg.unibe.ch/software/arlequin3 (accessed on 1 July 2012).
- Clement, M.; Posada, D.; Crandall, K.A. TCS: A computer program to estimate gene genealogies. Mol. Ecol. 2000, 9, 1657–1660. [Google Scholar] [CrossRef] [PubMed]
- Heemstra, P.C.; Randall, J.E. Groupers of the world (Family Serranidae, Subfamily Epinephelinae). In An Annotated and Illustrated Catalogue of the Grouper, Rockcod, Hind, Coral Grouper and Lyretail Species Known to Date; Food and Agriculture Organization of the United Nations: Rome, Italy, 1993; p. 382. [Google Scholar]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nurdalila, A.A.; Bunawan, H.; Kumar, S.V.; Rodrigues, K.F.; Baharum, S.N. Homogeneous Nature of Malaysian Marine Fish Epinephelus fuscoguttatus (Perciformes; Serranidae): Evidence Based on Molecular Markers, Morphology and Fourier Transform Infrared Analysis. Int. J. Mol. Sci. 2015, 16, 14884-14900. https://doi.org/10.3390/ijms160714884
Nurdalila AA, Bunawan H, Kumar SV, Rodrigues KF, Baharum SN. Homogeneous Nature of Malaysian Marine Fish Epinephelus fuscoguttatus (Perciformes; Serranidae): Evidence Based on Molecular Markers, Morphology and Fourier Transform Infrared Analysis. International Journal of Molecular Sciences. 2015; 16(7):14884-14900. https://doi.org/10.3390/ijms160714884
Chicago/Turabian StyleNurdalila, A'wani Aziz, Hamidun Bunawan, Subbiah Vijay Kumar, Kenneth Francis Rodrigues, and Syarul Nataqain Baharum. 2015. "Homogeneous Nature of Malaysian Marine Fish Epinephelus fuscoguttatus (Perciformes; Serranidae): Evidence Based on Molecular Markers, Morphology and Fourier Transform Infrared Analysis" International Journal of Molecular Sciences 16, no. 7: 14884-14900. https://doi.org/10.3390/ijms160714884
APA StyleNurdalila, A. A., Bunawan, H., Kumar, S. V., Rodrigues, K. F., & Baharum, S. N. (2015). Homogeneous Nature of Malaysian Marine Fish Epinephelus fuscoguttatus (Perciformes; Serranidae): Evidence Based on Molecular Markers, Morphology and Fourier Transform Infrared Analysis. International Journal of Molecular Sciences, 16(7), 14884-14900. https://doi.org/10.3390/ijms160714884