
BubR1 Acts as a Promoter in Cellular Motility of Human Oral Squamous Cancer Cells through Regulating MMP-2 and MMP-9

 ,
, 
Abstract
:1. Introduction
2. Results and Discussion
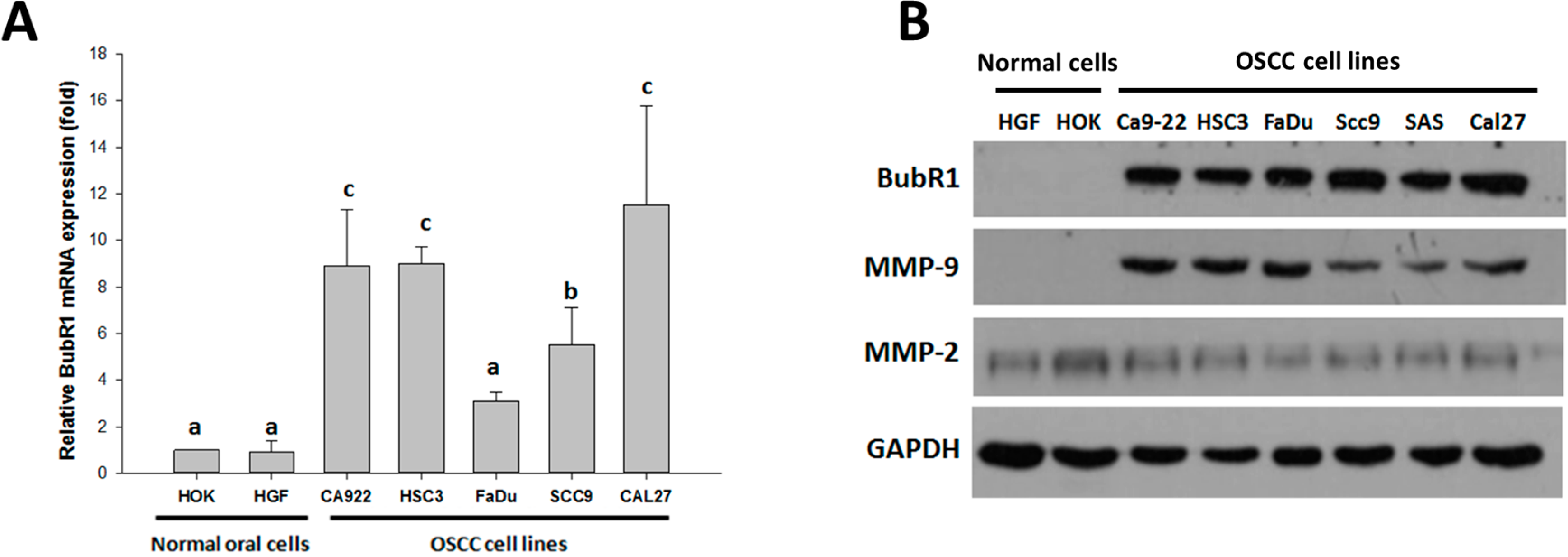
2.1. Expression of BubR1 in Oral Squamous Cell Carcinoma (OSCC) Cells

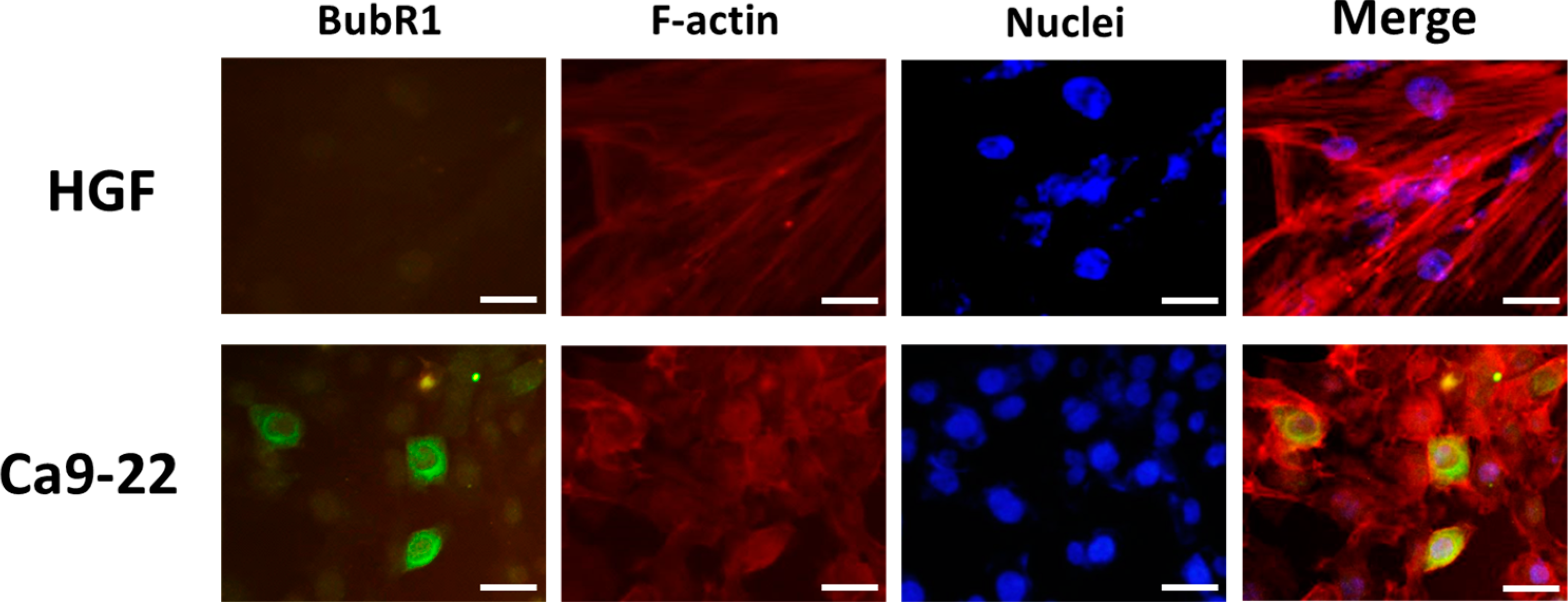
2.2. BubR1 Localizes Both in the Nuclear and Cytosol of OSCC Cells

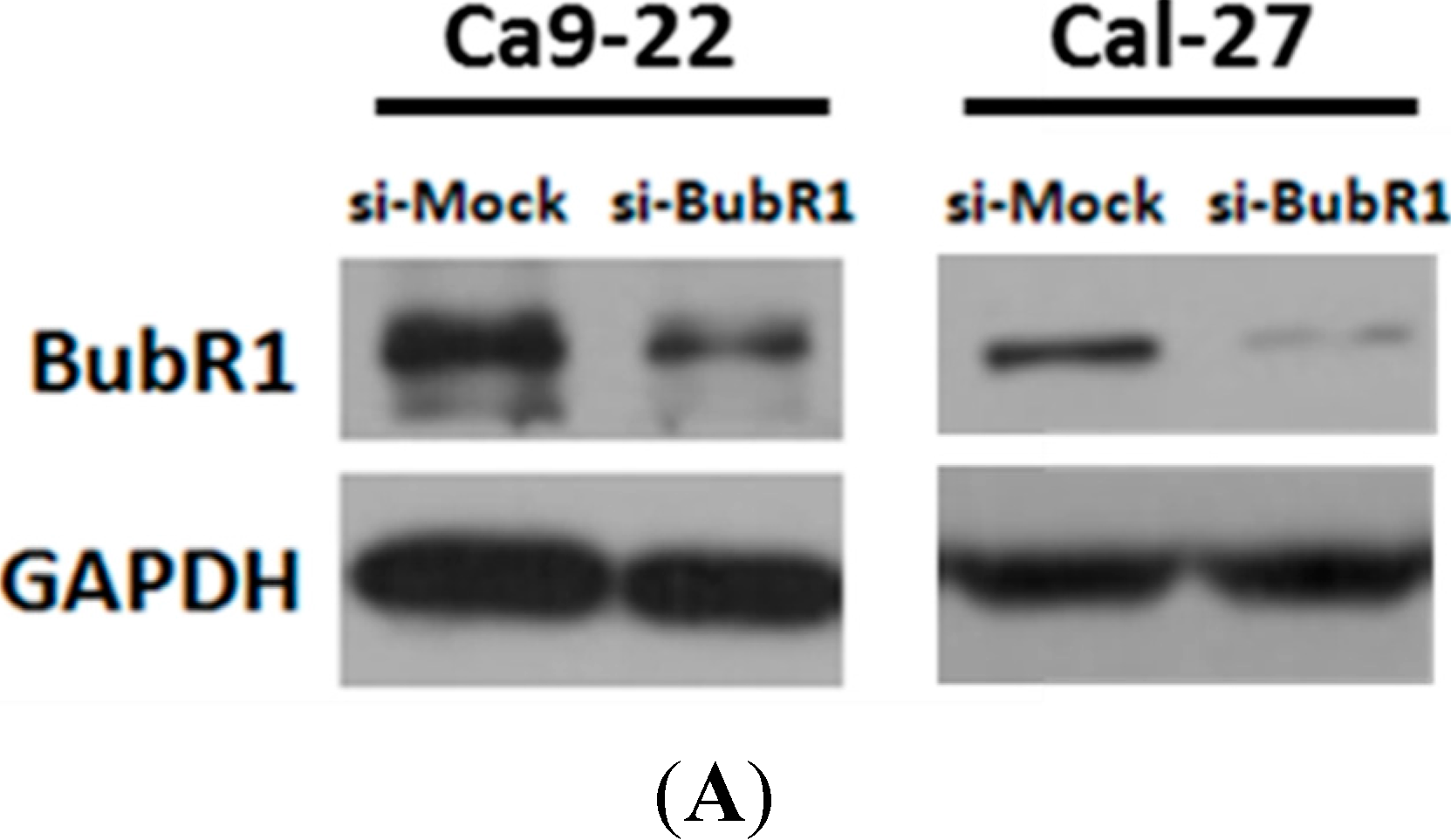
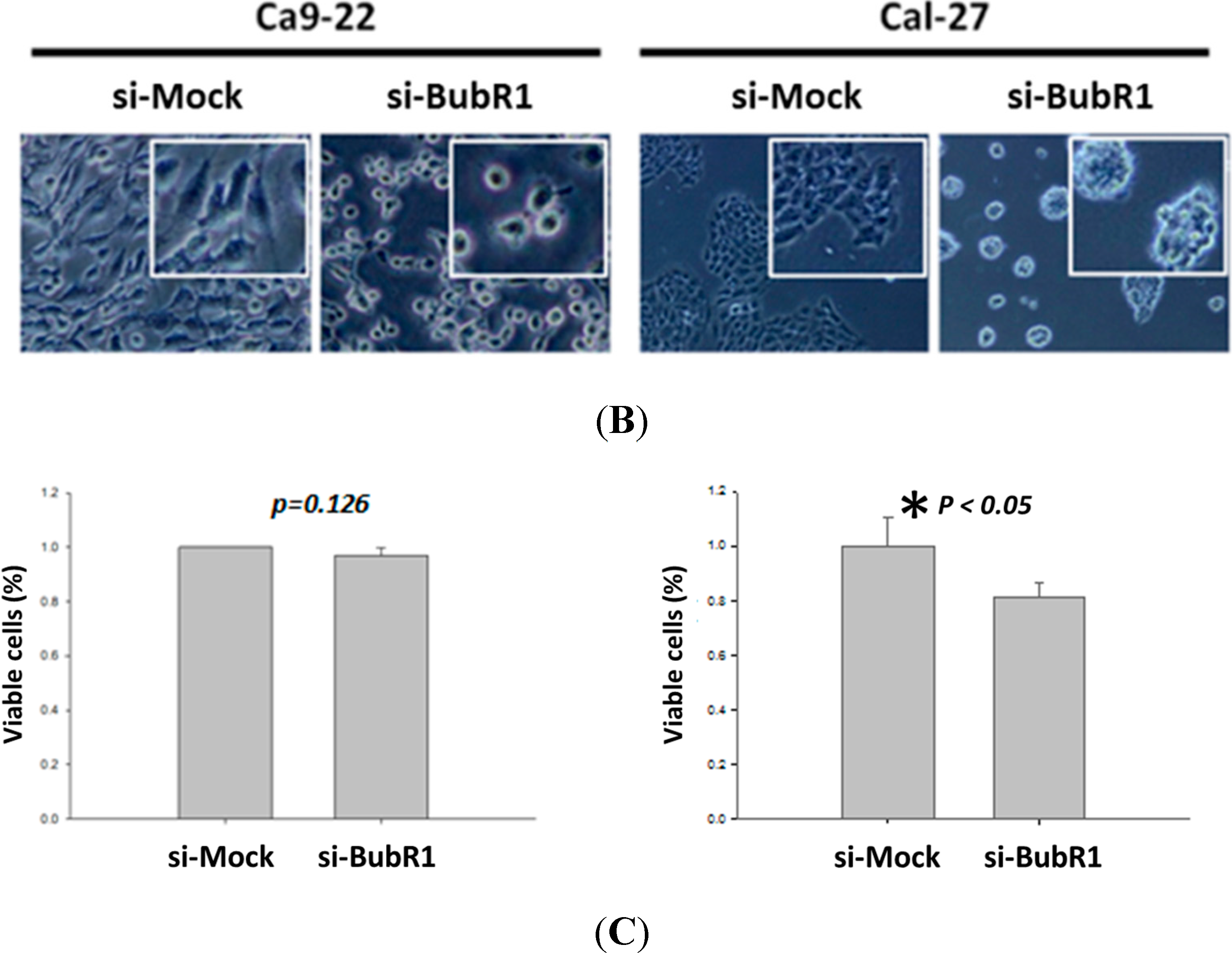
2.3. The Effect of BubR1 Knock-Down on Cell Morphology and Proliferation of OSCC Cells


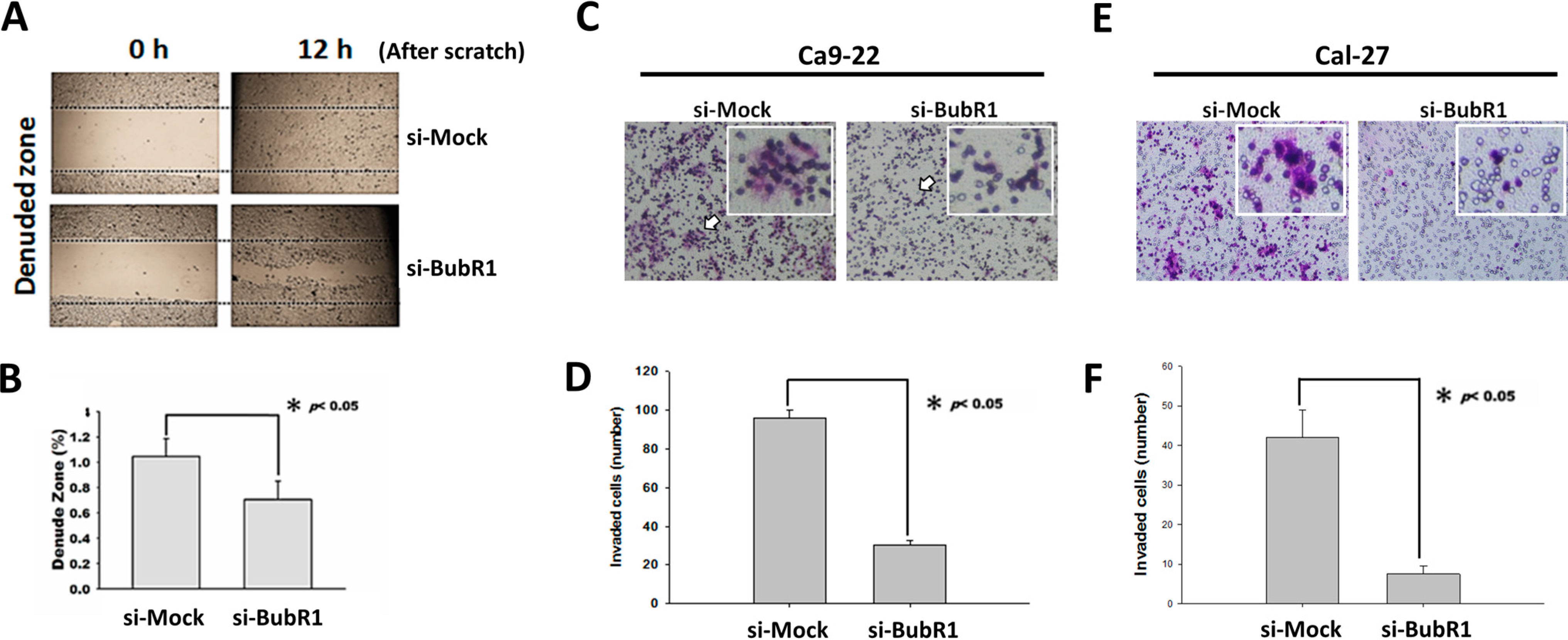
2.4. BubR1 Knockdown Reduces the Invasive Ability of OSCC Cells

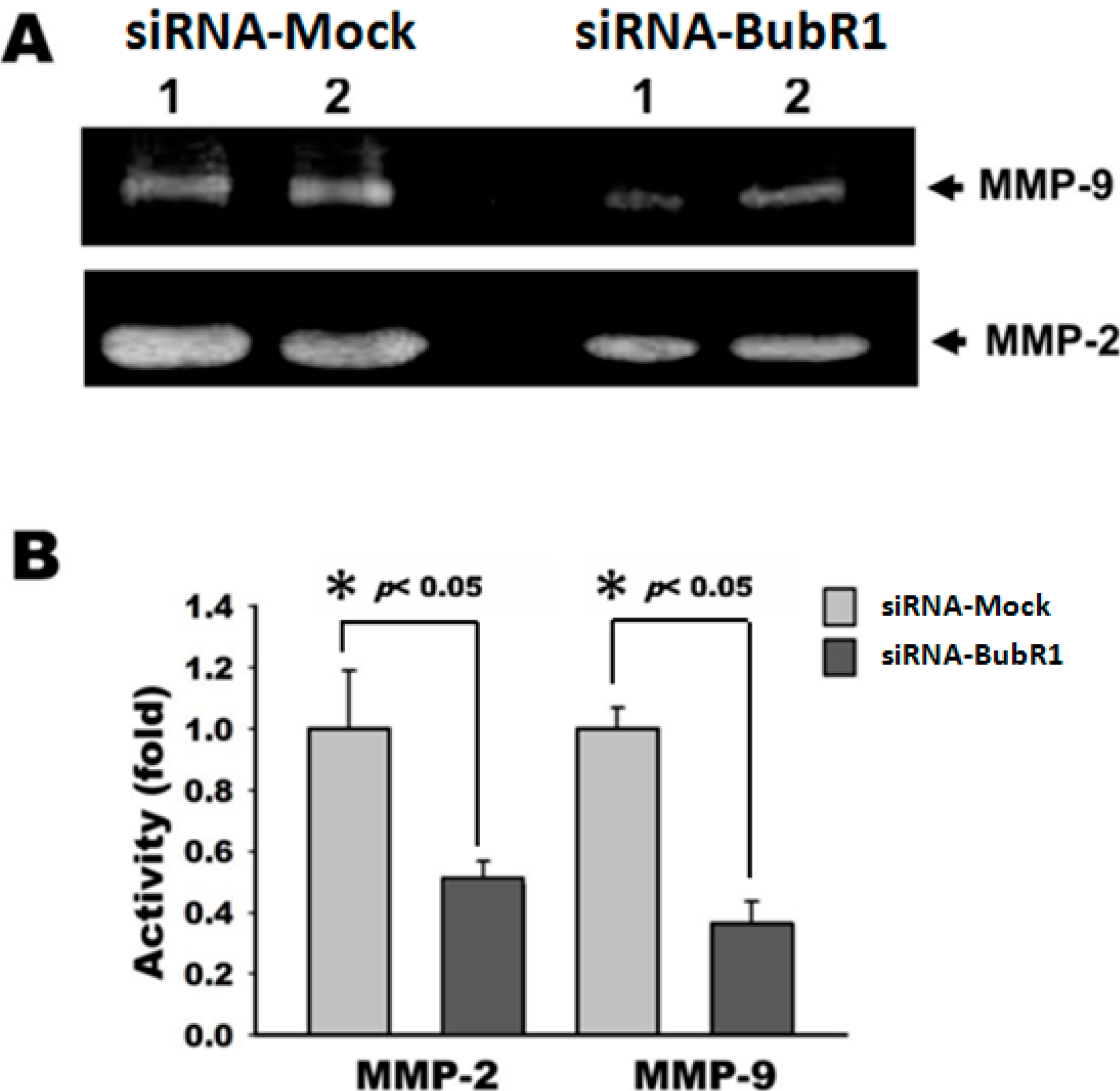
2.5. Knockdown of BubR1 Attenuates the Activity of MMP-2 and MMP-9

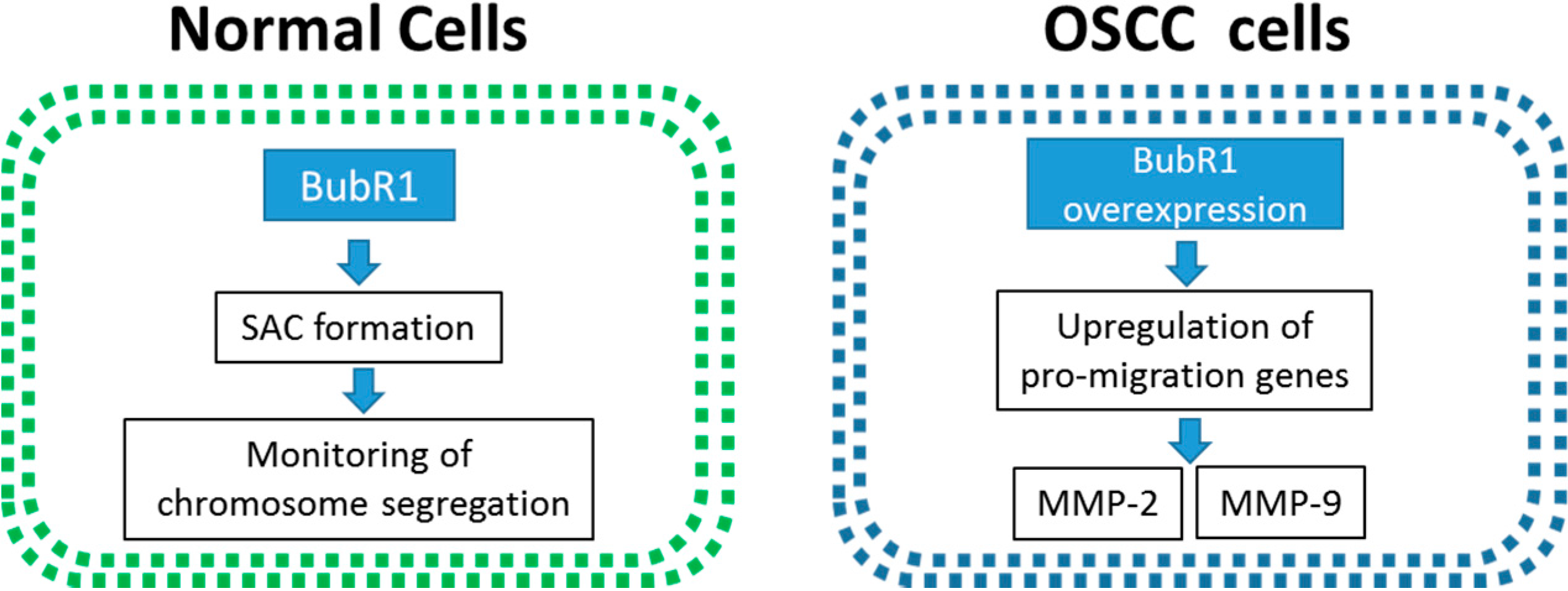
2.6. The Overexpression of BubR1 May Involve in the OSCC Tumorigenesis

3. Experimental Section
3.1. Cell Cultures
3.2. Quantitative Polymerase Chain Reaction Analysis

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Primers |
|---|---|
| BubR1 | Fʹ: 5ʹ-GCCTCGTGGCAATACAGCTT-3ʹ |
| Rʹ: 5ʹ-TGGTGTCATAACTGGCTGTTGTG-3ʹ | |
| gapdh | Fʹ: 5ʹ-GTCTTCACCACCATGGAGAA-3ʹ |
| Rʹ: 5ʹ-ATGGCATGGACTGTGGTCAT-3ʹ |
3.3. Immunofluorescence Staining
3.4. RNA Interference
3.5. Cell Proliferation Assay
3.6. Western Blotting
3.7. Wound Healing Assay
3.8. Transwell Invasion Assay
3.9. Gelatin Zymography
3.10. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Siegel, R.; Ma, J.; Zou, Z.; Jemal, A. Cancer statistics, 2014. CA Cancer J. Clin. 2014, 64, 9–29. [Google Scholar] [CrossRef] [PubMed]
- Noguti, J.; de Moura, C.F.; de Jesus, G.P.; da Silva, V.H.; Hossaka, T.A.; Oshima, C.T.; Ribeiro, D.A. Metastasis from oral cancer: An overview. Cancer Genomics Proteomics 2012, 9, 329–335. [Google Scholar] [PubMed]
- Choi, S.; Myers, J.N. Molecular pathogenesis of oral squamous cell carcinoma: Implications for therapy. J. Dent. Res. 2008, 87, 14–32. [Google Scholar] [CrossRef] [PubMed]
- Lea, J.; Bachar, G.; Sawka, A.M.; Lakra, D.C.; Gilbert, R.W.; Irish, J.C.; Brown, D.H.; Gullane, P.J.; Goldstein, D.P. Metastases to level IIb in squamous cell carcinoma of the oral cavity: A systematic review and meta-analysis. Head Neck 2010, 32, 184–190. [Google Scholar] [PubMed]
- Yadav, L.; Puri, N.; Rastogi, V.; Satpute, P.; Ahmad, R.; Kaur, G. Matrix metalloproteinases and cancer—Roles in threat and therapy. Asian Pac. J. Cancer Prev. 2014, 15, 1085–1091. [Google Scholar] [CrossRef] [PubMed]
- Mohtasham, N.; Babakoohi, S.; Shiva, A.; Shadman, A.; Kamyab-Hesari, K.; Shakeri, M.T.; Sharifi-Sistani, N. Immunohistochemical study of p53, Ki-67, MMP-2 and MMP-9 expression at invasive front of squamous cell and verrucous carcinoma in oral cavity. Pathol. Res. Pract 2013, 209, 110–114. [Google Scholar] [CrossRef] [PubMed]
- Stoeckli, S.J.; Alkureishi, L.W.; Ross, G.L. Sentinel node biopsy for early oral and oropharyngeal squamous cell carcinoma. Eur. Arch. Otorhinolaryngol. 2009, 266, 787–793. [Google Scholar] [CrossRef] [PubMed]
- Fan, S.; Tang, Q.L.; Lin, Y.J.; Chen, W.L.; Li, J.S.; Huang, Z.Q.; Yang, Z.H.; Wang, Y.Y.; Zhang, D.M.; Wang, H.J.; et al. A review of clinical and histological parameters associated with contralateral neck metastases in oral squamous cell carcinoma. Int. J. Oral Sci. 2011, 3, 180–191. [Google Scholar] [CrossRef] [PubMed]
- Bolanos-Garcia, V.M.; Blundell, T.L. BUB1 and BUBR1: Multifaceted kinases of the cell cycle. Trends Biochem. Sci. 2011, 36, 141–150. [Google Scholar] [CrossRef] [PubMed]
- Matsuura, S.; Matsumoto, Y.; Morishima, K.; Izumi, H.; Matsumoto, H.; Ito, E.; Tsutsui, K.; Kobayashi, J.; Tauchi, H.; Kajiwara, Y.; et al. Monoallelic BUB1B mutations and defective mitotic-spindle checkpoint in seven families with premature chromatid separation (PCS) syndrome. Am. J. Med. Genet. A 2006, 140, 358–367. [Google Scholar] [CrossRef] [PubMed]
- Gupta, A.; Inaba, S.; Wong, O.K.; Fang, G.; Liu, J. Breast cancer-specific gene 1 interacts with the mitotic checkpoint kinase Bubr1. Oncogene 2003, 22, 7593–7599. [Google Scholar] [CrossRef] [PubMed]
- Ando, K.; Kakeji, Y.; Kitao, H.; Iimori, M.; Zhao, Y.; Yoshida, R.; Oki, E.; Yoshinaga, K.; Matumoto, T.; Morita, M.; et al. High expression of BUBR1 is one of the factors for inducing DNA aneuploidy and progression in gastric cancer. Cancer Sci. 2010, 101, 639–645. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, P.C.; Chen, Y.K.; Tsai, K.B.; Shieh, T.Y.; Chang, Y.Y.; Chang, J.G.; Wu, H.L.; Lin, S.F. Expression of BUBR1 in human oral potentially malignant disorders and squamous cell carcinoma. Oral Surg. Oral Med. Oral Pathol. Oral Radiol. Endod. 2010, 109, 257–267. [Google Scholar] [CrossRef] [PubMed]
- Rizzardi, C.; Torelli, L.; Barresi, E.; Schneider, M.; Canzonieri, V.; Biasotto, M.; di Lenarda, R.; Melato, M. Bubr1 expression in oral squamous cell carcinoma and its relationship to tumor stage and survival. Head Neck 2011, 33, 727–733. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, Y.; Matsuyama, H.; Chochi, Y.; Okuda, M.; Kawauchi, S.; Inoue, R.; Furuya, T.; Oga, A.; Naito, K.; Sasaki, K. Overexpression of BUBR1 is associated with chromosomal instability in bladder cancer. Cancer Genet. Cytogenet. 2007, 174, 42–47. [Google Scholar] [CrossRef] [PubMed]
- Lira, R.C.; Miranda, F.A.; Guimaraes, M.C.; Simoes, R.T.; Donadi, E.A.; Soares, C.P.; Soares, E.G. BUBR1 expression in benign oral lesions and squamous cell carcinomas: Correlation with human papillomavirus. Oncol. Rep. 2010, 23, 1027–1036. [Google Scholar] [PubMed]
- Tseng, M.Y.; Liu, S.Y.; Chen, H.R.; Wu, Y.J.; Chiu, C.C.; Chan, P.T.; Chiang, W.F.; Liu, Y.C.; Lu, C.Y.; Jou, Y.S.; et al. Serine protease inhibitor (SERPIN) B1 promotes oral cancer cell motility and is over-expressed in invasive oral squamous cell carcinoma. Oral Oncol. 2009, 45, 771–776. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.K.; Choi, E.; Kim, M.A.; Park, P.G.; Park, N.H.; Lee, H. BUBR1 as a prognostic marker for recurrence-free survival rates in epithelial ovarian cancers. Br. J. Cancer 2009, 101, 504–510. [Google Scholar] [CrossRef] [PubMed]
- Cirak, Y.; Sarsik, B.; Cakar, B.; Sen, S.; Simsir, A.; Uslu, R. Predictive and prognostic values of tau and BUBR1 protein in prostate cancer and their relationship to the gleason score. Med. Oncol. 2013, 30. [Google Scholar] [CrossRef] [PubMed]
- Geback, T.; Schulz, M.M.; Koumoutsakos, P.; Detmar, M. Tscratch: A novel and simple software tool for automated analysis of monolayer wound healing assays. Biotechniques 2009, 46, 265–274. [Google Scholar] [PubMed]
- Nagase, H.; Visse, R.; Murphy, G. Structure and function of matrix metalloproteinases and timps. Cardiovasc. Res. 2006, 69, 562–573. [Google Scholar] [CrossRef] [PubMed]
- Koivunen, E.; Arap, W.; Valtanen, H.; Rainisalo, A.; Medina, O.P.; Heikkila, P.; Kantor, C.; Gahmberg, C.G.; Salo, T.; Konttinen, Y.T.; et al. Tumor targeting with a selective gelatinase inhibitor. Nat. Biotechnol. 1999, 17, 768–774. [Google Scholar] [CrossRef] [PubMed]
- Ferrand, A.; Chevrier, V.; Chauvin, J.P.; Birnbaum, D. Ajuba: A new microtubule-associated protein that interacts with BUBR1 and aurora B at kinetochores in metaphase. Biol. Cell 2009, 101, 221–235. [Google Scholar] [CrossRef] [PubMed]
- Honore, S.; Pasquier, E.; Braguer, D. Understanding microtubule dynamics for improved cancer therapy. Cell Mol. Life Sci. 2005, 62, 3039–3056. [Google Scholar] [CrossRef] [PubMed]
- Chiu, C.C.; Chen, J.Y.; Lin, K.L.; Huang, C.J.; Lee, J.C.; Chen, B.H.; Chen, W.Y.; Lo, Y.H.; Chen, Y.L.; Tseng, C.H.; et al. p38 MAPK and NF-κB pathways are involved in naphtho[1,2-b] furan-4,5-dione induced anti-proliferation and apoptosis of human hepatoma cells. Cancer Lett. 2010, 295, 92–99. [Google Scholar] [CrossRef] [PubMed]
- Chiu, C.C.; Liu, P.L.; Huang, K.J.; Wang, H.M.; Chang, K.F.; Chou, C.K.; Chang, F.R.; Chong, I.W.; Fang, K.; Chen, J.S.; et al. Goniothalamin inhibits growth of human lung cancer cells through DNA damage, apoptosis, and reduced migration ability. J. Agric. Food Chem. 2011, 59, 4288–4293. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chou, C.-K.; Wu, C.-Y.; Chen, J.Y.-F.; Ng, M.-C.; Wang, H.-M.D.; Chen, J.-H.; Yuan, S.-S.F.; Tsai, E.-M.; Chang, J.-G.; Chiu, C.-C. BubR1 Acts as a Promoter in Cellular Motility of Human Oral Squamous Cancer Cells through Regulating MMP-2 and MMP-9. Int. J. Mol. Sci. 2015, 16, 15104-15117. https://doi.org/10.3390/ijms160715104
Chou C-K, Wu C-Y, Chen JY-F, Ng M-C, Wang H-MD, Chen J-H, Yuan S-SF, Tsai E-M, Chang J-G, Chiu C-C. BubR1 Acts as a Promoter in Cellular Motility of Human Oral Squamous Cancer Cells through Regulating MMP-2 and MMP-9. International Journal of Molecular Sciences. 2015; 16(7):15104-15117. https://doi.org/10.3390/ijms160715104
Chicago/Turabian StyleChou, Chou-Kit, Chang-Yi Wu, Jeff Yi-Fu Chen, Ming-Chong Ng, Hui-Min David Wang, Jen-Hao Chen, Shyng-Shiou F. Yuan, Eing-Mei Tsai, Jan-Gowth Chang, and Chien-Chih Chiu. 2015. "BubR1 Acts as a Promoter in Cellular Motility of Human Oral Squamous Cancer Cells through Regulating MMP-2 and MMP-9" International Journal of Molecular Sciences 16, no. 7: 15104-15117. https://doi.org/10.3390/ijms160715104
APA StyleChou, C. -K., Wu, C. -Y., Chen, J. Y. -F., Ng, M. -C., Wang, H. -M. D., Chen, J. -H., Yuan, S. -S. F., Tsai, E. -M., Chang, J. -G., & Chiu, C. -C. (2015). BubR1 Acts as a Promoter in Cellular Motility of Human Oral Squamous Cancer Cells through Regulating MMP-2 and MMP-9. International Journal of Molecular Sciences, 16(7), 15104-15117. https://doi.org/10.3390/ijms160715104