
Characterization of a Type 1 Metallothionein Gene from the Stresses-Tolerant Plant Ziziphus jujuba

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
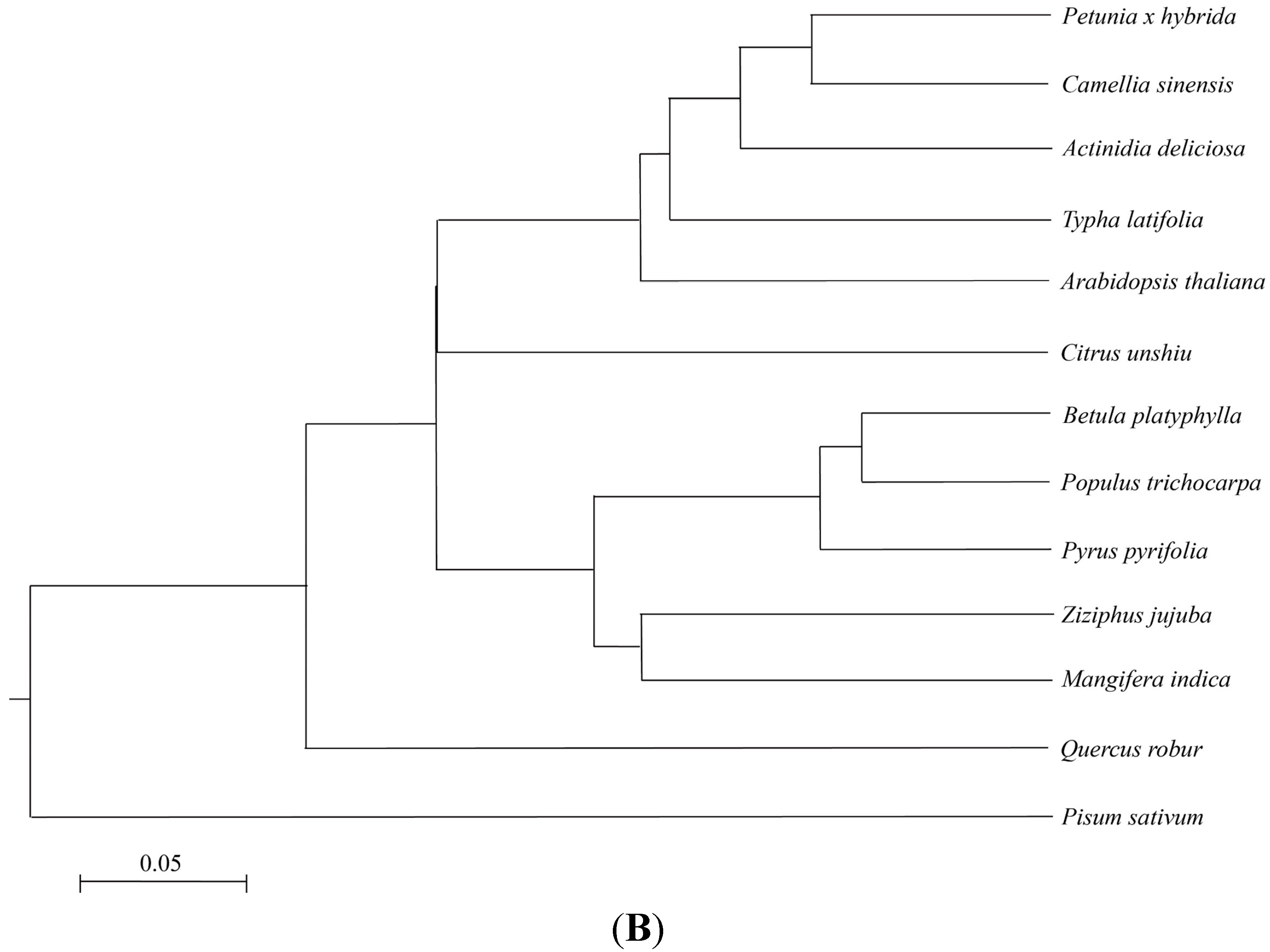
2.1. ZjMT Encodes a Protein with a Metallothionein (MT) Domain


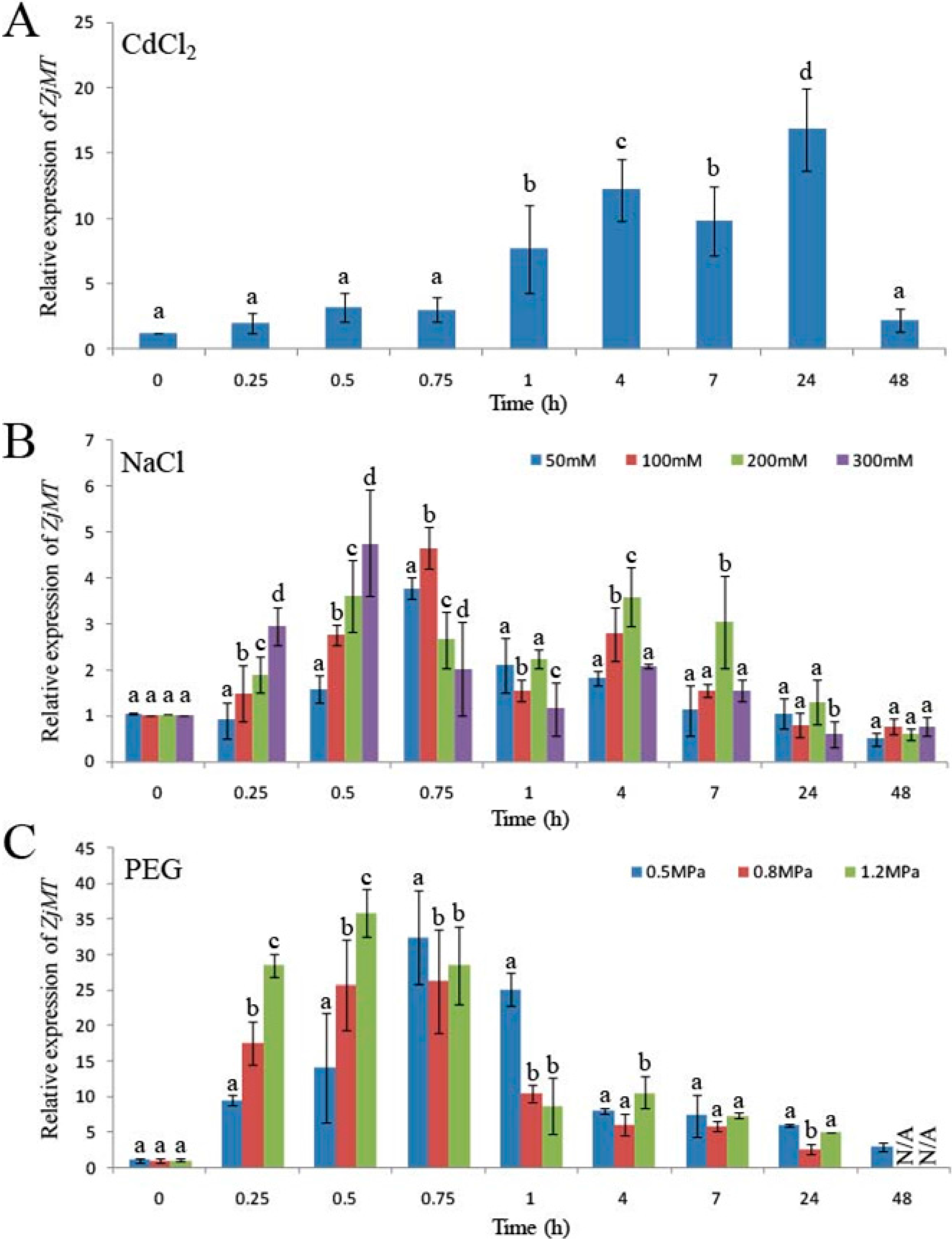
2.2. ZjMT Is a Potential Stress-Related Gene

2.3. Subcelluar Localization of ZjMT

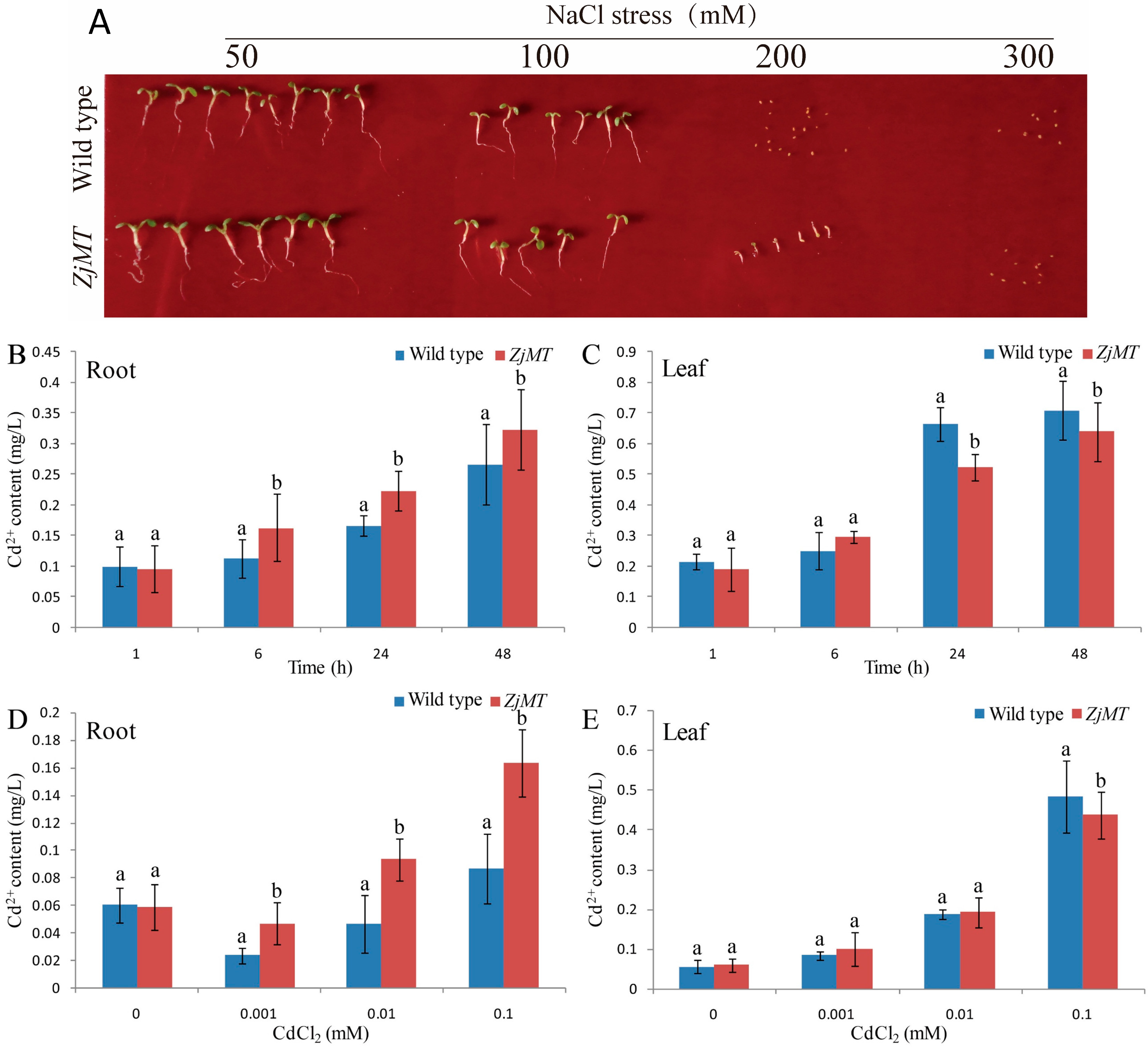
2.4. Constitutive Expression of ZjMT in Arabidopsis Enhances Their High Salinity Salt Tolerance

2.5. Cd2+ Accumulation and Distribution in Transgenic Plants
3. Discussion
4. Experimental Section
4.1. Stress Treatments and Real-Time Polymerase Chain Reaction (PCR) Analyses
4.2. Sequence Analysis of ZjMT
4.3. Subcellular Localization
4.4. Generation of Transgenic Plants
4.5. Stress Treatments
4.6. Measurement of Cd2+ Content
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Hall, J. Cellular mechanisms for heavy metal detoxification and tolerance. J. Exp. Bot. 2002, 53, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Sugiyama, M. Role of cellular antioxidants in metal-induced damage. Cell Biol. Toxicol. 1994, 10, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Benavides, M.P.; Gallego, S.M.; Tomaro, M.L. Cadmium toxicity in plants. Braz. J. Plant Physiol. 2005, 17, 21–34. [Google Scholar] [CrossRef]
- Trvedi, S.; Erdei, L. Effects of cadmium and lead on the accumulation of Ca2+ and K+ and on the influx and translocation of K+ in wheat of low and high K+ status. Physiol. Plant. 1992, 84, 94–100. [Google Scholar] [CrossRef]
- Hernandez, L.; Garate, A.; Carpena-Ruiz, R. Effects of cadmium on the uptake, distribution and assimilation of nitrate in Pisum sativum. Plant Soil 1997, 189, 97–106. [Google Scholar] [CrossRef]
- Krupa, Z.; Öquist, G.; Huner, N. The effects of cadmium on photosynthesis of Phaseolus vulgaris—A fluorescence analysis. Physiol. Plant. 1993, 88, 626–630. [Google Scholar] [CrossRef]
- Fodor, E.; Szabó-Nagy, A.; Erdei, L. The effects of cadmium on the fluidity and H+-ATPase activity of plasma membrane from sunflower and wheat roots. J. Plant Physiol. 1995, 147, 87–92. [Google Scholar] [CrossRef]
- Kägi, J.H.; Vallee, B.L. Metallothionein: A cadmium-and zinc-containing protein from equine renal cortex. J. Biol. Chem. 1960, 235, 3460–3465. [Google Scholar] [PubMed]
- Kojima, Y.; Berger, C.; Vallee, B.L.; Kägi, J. Amino-acid sequence of equine renal metallothionein-1B. Proc. Natl. Acad. Sci. USA 1976, 73, 3413–3417. [Google Scholar] [CrossRef] [PubMed]
- Margoshes, M.; Vallee, B.L. A cadmium protein from equine kidney cortex. J. Am. Chem. Soc. 1957, 79, 4813–4814. [Google Scholar] [CrossRef]
- Loebus, J.; Leitenmaier, B.; Meissner, D.; Braha, B.; Krauss, G.J.; Dobritzsch, D.; Freisinger, E. The major function of a metallothionein from the aquatic fungus Heliscus lugdunensis is cadmium detoxification. J. Inorg. Biochem. 2013, 127, 253–260. [Google Scholar] [CrossRef] [PubMed]
- Dąbrowska, G. Plant metallothioneins: Putative functions identified by promoter analysis in silico. Acta Biol. Cracov. Bot. 2012, 54, 109–120. [Google Scholar] [CrossRef]
- Zhou, B.; Yao, W.; Wang, S.; Wang, X.; Jiang, T. The metallothionein gene, TaMT3, from Tamarix androssowii confers Cd2+ tolerance in tobacco. Int. J. Mol. Sci. 2014, 15, 10398–10409. [Google Scholar] [CrossRef] [PubMed]
- Lv, Y.; Deng, X.; Quan, L.; Xia, Y.; Shen, Z. Metallothioneins BcMT1 and BcMT2 from Brassica campestris enhance tolerance to cadmium and copper and decrease production of reactive oxygen species in Arabidopsis thaliana. Plant Soil 2013, 367, 507–519. [Google Scholar] [CrossRef]
- Hassinen, V.; Tervahauta, A.; Schat, H.; Kärenlampi, S. Plant metallothioneins-metal chelators with ROS scavenging activity? Plant Biol. 2011, 13, 225–232. [Google Scholar] [CrossRef] [PubMed]
- Higashimoto, M.; Isoyama, N.; Ishibashi, S.; Inoue, M.; Takiguchi, M.; Suzuki, S.; Ohnishi, Y.; Sato, M. Tissue-dependent preventive effect of metallothionein against DNA damage in dyslipidemic mice under repeated stresses of fasting or restraint. Life Sci. 2009, 84, 569–575. [Google Scholar] [CrossRef] [PubMed]
- Cobbett, C.; Goldsbrough, P. Phytochelatins and metallothioneins: Roles in heavy metal detoxification and homeostasis. Annu. Rev. Plant Biol. 2002, 53, 159–182. [Google Scholar] [CrossRef] [PubMed]
- Robinson, N.J.; Tommey, A.M.; Kuske, C.; Jackson, P.J. Plant metallothioneins. Biochem. J. 1993, 295, 1. [Google Scholar] [PubMed]
- Capdevila, M.; Atrian, S. Metallothionein protein evolution: A miniassay. J. Biol. Inorg. Chem. 2011, 16, 977–989. [Google Scholar] [CrossRef] [PubMed]
- Guo, W.J.; Bundithya, W.; Goldsbrough, P.B. Characterization of the Arabidopsis metallothionein gene family: Tissue-specific expression and induction during senescence and in response to copper. New Phytol. 2003, 159, 369–381. [Google Scholar] [CrossRef]
- Murphy, A.; Zhou, J.; Goldsbrough, P.B.; Taiz, L. Purification and immunological identification of metallothioneins 1 and 2 from Arabidopsis thaliana. Plant Physiol. 1997, 113, 1293–1301. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Goldsbrough, P.B. Structure, organization and expression of the metallothionein gene family in Arabidopsis. Mol. Gen. Genet. 1995, 248, 318–328. [Google Scholar] [PubMed]
- Waters, D.L.; Holton, T.A.; Ablett, E.M.; Lee, L.S.; Henry, R.J. cDNA microarray analysis of developing grape (Vitis vinifera cv. Shiraz) berry skin. Funct. Integr. Genomic 2005, 5, 40–58. [Google Scholar] [CrossRef] [PubMed]
- Ledger, S.E.; Gardner, R.C. Cloning and characterization of five cDNAs for genes differentially expressed during fruit development of kiwifruit (Actinidia deliciosa var. deliciosa). Plant Mol. Biol. 1994, 25, 877–886. [Google Scholar] [CrossRef] [PubMed]
- Chyan, C.L.; Lee, T.T.; Liu, C.P.; Yang, Y.C.; Tzen, J.T.; Chou, W.M. Cloning and expression of a seed-specific metallothionein-like protein from sesame. Biosci. Biotechnol. Biochem. 2005, 69, 2319–2325. [Google Scholar] [CrossRef] [PubMed]
- Leszczyszyn, O.I.; Imam, H.T.; Blindauer, C.A. Diversity and distribution of plant metallothioneins: A review of structure, properties and functions. Metallomics 2013, 5, 1146–1169. [Google Scholar] [CrossRef] [PubMed]
- Kawashima, I.; Kennedy, T.D.; Chino, M.; Lane, B.G. Wheat Ec metallothionein genes: Like mammalian Zn2+ metallothionein genes are conspicuously expressed during embryogenesis. Eur. J. Biochem. 1992, 209, 971–976. [Google Scholar] [CrossRef] [PubMed]
- Kawashima, I.; Inokuchi, Y.; Chino, M.; Kimura, M.; Shimizu, N. Isolation of a gene for a metallothionein-like protein from soybean. Plant Cell Physiol. 1991, 32, 913–916. [Google Scholar]
- Hsieh, H.-M.; Liu, W.-K.; Huang, P. A novel stress-inducible metallothionein-like gene from rice. Plant Mol. Biol. 1995, 28, 381–389. [Google Scholar] [CrossRef] [PubMed]
- Giritch, A.; Ganal, M.; Stephan, U.W.; Bäumlein, H. Structure, expression and chromosomal localisation of the metallothionein-like gene family of tomato. Plant Mol. Biol. 1998, 37, 701–714. [Google Scholar] [CrossRef] [PubMed]
- Moyle, R.; Fairbairn, D.J.; Ripi, J.; Crowe, M.; Botella, J.R. Developing pineapple fruit has a small transcriptome dominated by metallothionein. J. Exp. Bot. 2005, 56, 101–112. [Google Scholar] [CrossRef] [PubMed]
- Yuan, J.; Chen, D.; Ren, Y.; Zhang, X.; Zhao, J. Characteristic and expression analysis of a metallothionein gene, OsMT2b, down-regulated by cytokinin suggests functions in root development and seed embryo germination of rice. Plant Physiol. 2008, 146, 1637–1650. [Google Scholar] [CrossRef] [PubMed]
- Mir, G.; Domènech, J.; Huguet, G.; Guo, W.J.; Goldsbrough, P.; Atrian, S.; Molinas, M. A plant type 2 metallothionein (MT) from cork tissue responds to oxidative stress. J. Exp. Bot. 2004, 55, 2483–2493. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Chen, Y.Y.; Yang, S.G.; Tian, W.M. Cloning and characterization of HbMT2a, a metallothionein gene from Hevea brasiliensis Muell. Arg differently responds to abiotic stress and heavy metals. Biochem. Biophys. Res. Commun. 2015, 461, 95–101. [Google Scholar] [CrossRef] [PubMed]
- Zimeri, A.M.; Dhankher, O.P.; McCaig, B.; Meagher, R.B. The plant MT1 metallothioneins are stabilized by binding cadmiums and are required for cadmium tolerance and accumulation. Plant Mol. Biol. 2005, 58, 839–855. [Google Scholar] [CrossRef] [PubMed]
- Xia, Y.; Lv, Y.; Yuan, Y.; Wang, G.; Chen, Y.; Zhang, H.; Shen, Z. Cloning and characterization of a type 1 metallothionein gene from the copper-tolerant plant Elsholtzia haichowensis. Acta Physiol. Plant 2012, 34, 1819–1826. [Google Scholar] [CrossRef]
- Yang, Z.; Wu, Y.; Li, Y.; Ling, H.Q.; Chu, C. OsMT1a, a type 1 metallothionein, plays the pivotal role in zinc homeostasis and drought tolerance in rice. Plant Mol. Biol. 2009, 70, 219–229. [Google Scholar] [CrossRef] [PubMed]
- Grennan, A.K. Metallothioneins, a diverse protein family. Plant Physiol. 2011, 155, 1750–1751. [Google Scholar] [CrossRef] [PubMed]
- An, Z.; Cuijie, L.; Yuangang, Z.; Yejie, D.; Wachter, A.; Gromes, R.; Rausch, T. Expression of BjMT2, a metallothionein 2 from Brassica juncea, increases copper and cadmium tolerance in Escherichia coli and Arabidopsis thaliana, but inhibits root elongation in Arabidopsis thaliana seedlings. J. Exp. Bot. 2006, 57, 3575–3582. [Google Scholar]
- Ren, Y.; Liu, Y.; Chen, H.; Li, G.; Zhang, X.; Zhao, J. Type 4 metallothionein genes are involved in regulating Zn ion accumulation in late embryo and in controlling early seedling growth in Arabidopsis. Plant Cell Environ. 2012, 35, 770–789. [Google Scholar] [CrossRef] [PubMed]
- Kawasaki, S.; Borchert, C.; Deyholos, M.; Wang, H.; Brazille, S.; Kawai, K.; Galbraith, D.; Bohnert, H.J. Gene expression profiles during the initial phase of salt stress in rice. Plant Cell 2001, 13, 889–905. [Google Scholar] [CrossRef] [PubMed]
- Ozturk, Z.N.; Talamé, V.; Deyholos, M.; Michalowski, C.B.; Galbraith, D.W.; Gozukirmizi, N.; Tuberosa, R.; Bohnert, H.J. Monitoring large-scale changes in transcript abundance in drought- and salt-stressed barley. Plant Mol. Biol. 2002, 48, 551–573. [Google Scholar] [CrossRef]
- Sekhar, K.; Priyanka, B.; Reddy, V.; Rao, K. Metallothionein 1 (CcMT1) of pigeonpea (Cajanus cajan, L.) confers enhanced tolerance to copper and cadmium in Escherichia coli and Arabidopsis thaliana. Environ. Exp. Bot. 2011, 72, 131–139. [Google Scholar] [CrossRef]
- Zhao, J.; Sun, Z.; Zheng, J.; Guo, X.; Dong, Z.; Huai, J.; Gou, M.; He, J.; Jin, Y.; Wang, J. Cloning and characterization of a novel CBL-interacting protein kinase from maize. Plant Mol. Biol. 2009, 69, 661–674. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Dudley, J.; Nei, M.; Kumar, S. MEGA4: Molecular evolutionary genetics analysis (MEGA) software version 4.0. Mol. Biol. Evol. 2007, 24, 1596–1599. [Google Scholar] [CrossRef] [PubMed]
- Earley, K.W.; Haag, J.R.; Pontes, O.; Opper, K.; Juehne, T.; Song, K.; Pikaard, C.S. Gateway-compatible vectors for plant functional genomics and proteomics. Plant J. 2006, 45, 616–629. [Google Scholar] [CrossRef] [PubMed]
- Clough, S.J.; Bent, A.F. Floral dip: A simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J. 1998, 16, 735–743. [Google Scholar] [CrossRef] [PubMed]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, M.; Zhang, F.; Wang, F.; Dong, Z.; Cao, Q.; Chen, M. Characterization of a Type 1 Metallothionein Gene from the Stresses-Tolerant Plant Ziziphus jujuba. Int. J. Mol. Sci. 2015, 16, 16750-16762. https://doi.org/10.3390/ijms160816750
Yang M, Zhang F, Wang F, Dong Z, Cao Q, Chen M. Characterization of a Type 1 Metallothionein Gene from the Stresses-Tolerant Plant Ziziphus jujuba. International Journal of Molecular Sciences. 2015; 16(8):16750-16762. https://doi.org/10.3390/ijms160816750
Chicago/Turabian StyleYang, Mingxia, Fan Zhang, Fan Wang, Zhigang Dong, Qiufen Cao, and Mingchang Chen. 2015. "Characterization of a Type 1 Metallothionein Gene from the Stresses-Tolerant Plant Ziziphus jujuba" International Journal of Molecular Sciences 16, no. 8: 16750-16762. https://doi.org/10.3390/ijms160816750
APA StyleYang, M., Zhang, F., Wang, F., Dong, Z., Cao, Q., & Chen, M. (2015). Characterization of a Type 1 Metallothionein Gene from the Stresses-Tolerant Plant Ziziphus jujuba. International Journal of Molecular Sciences, 16(8), 16750-16762. https://doi.org/10.3390/ijms160816750